
Phylogenetic Review of Acaulospora (Diversisporales, Glomeromycota) and the Homoplasic Nature of Its Ornamentations

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Acaulospora: A Review of Its Taxonomy, Morphology and Molecular Markers
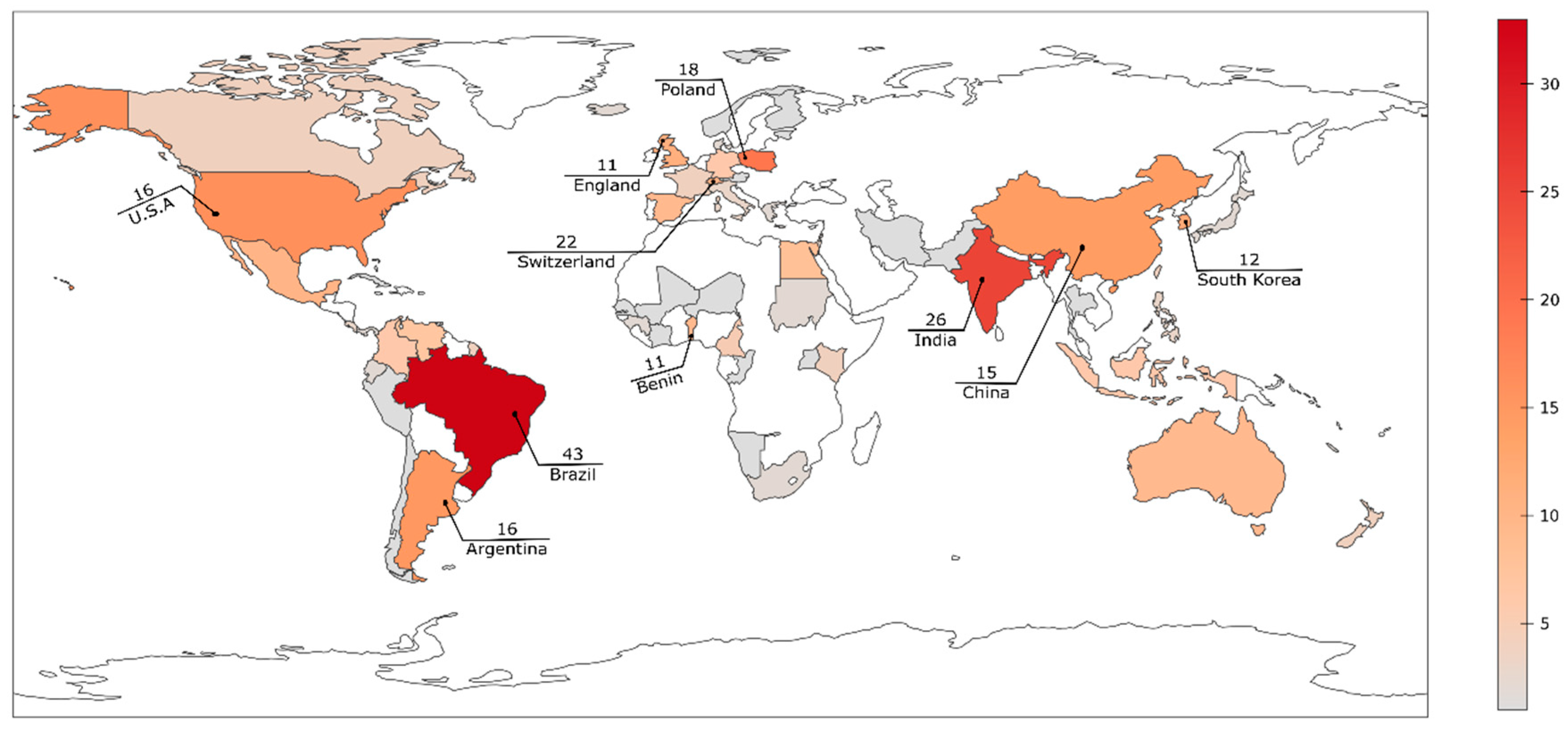
3. Diversity and Distribution of Acaulospora Species on the Globe
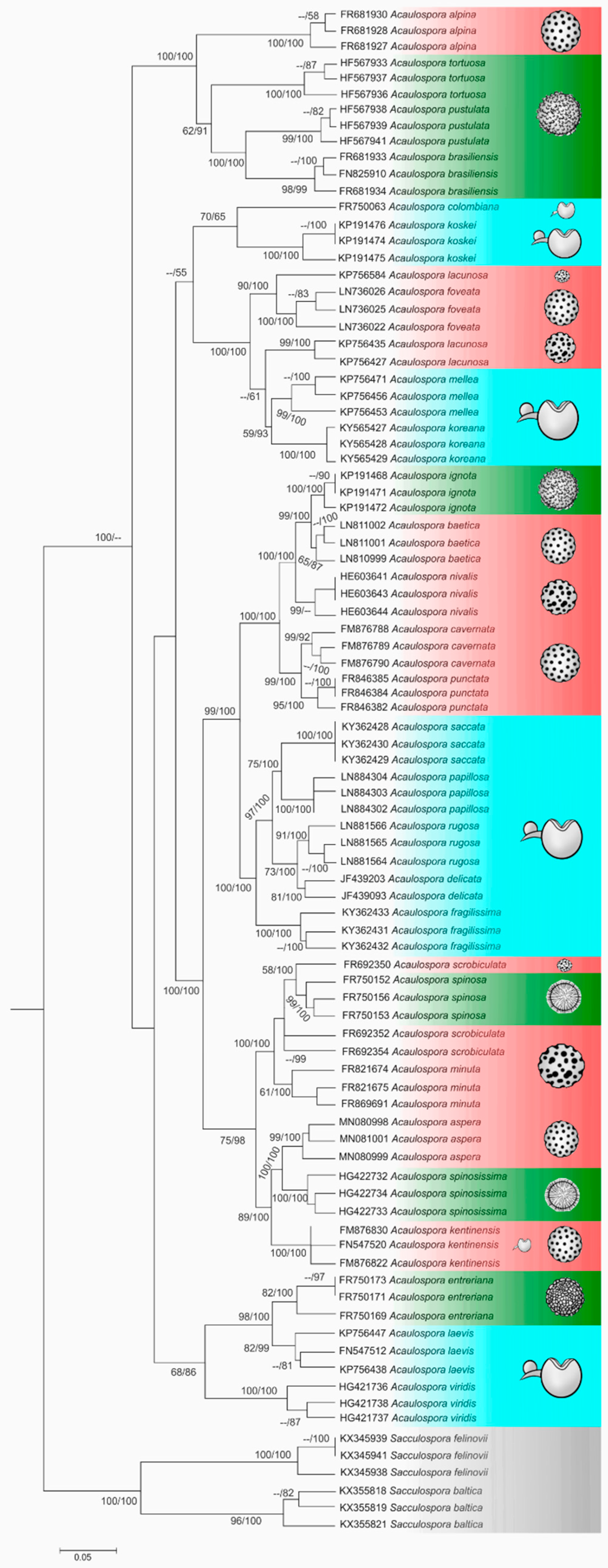
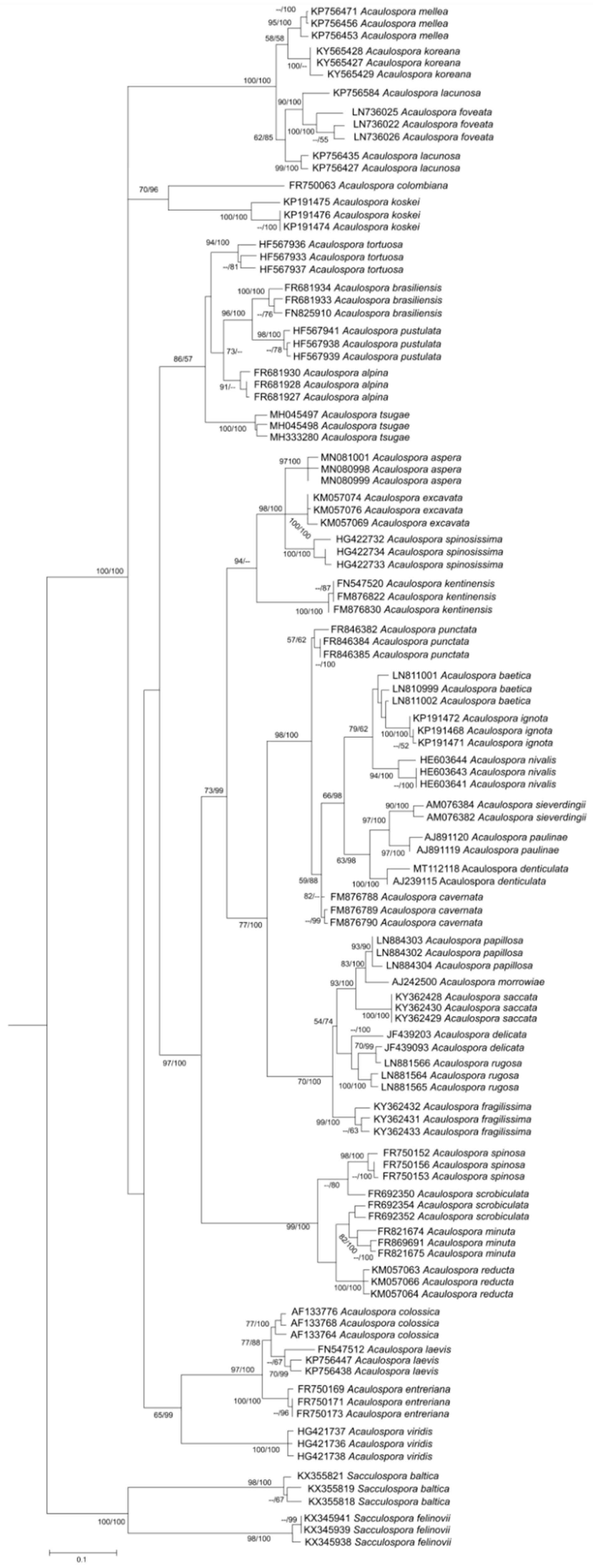
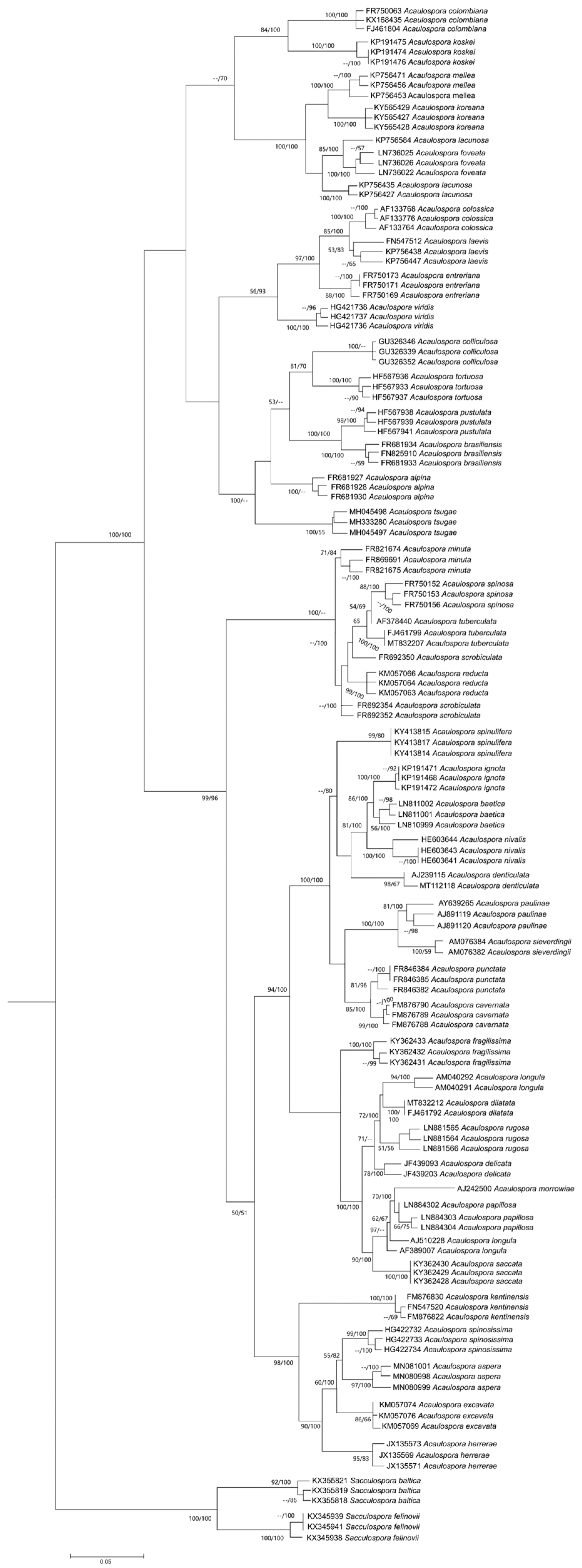
4. Phylogenetic Relationship among Acaulospora Species: Is There a Consensual Tree?
4.1. Is Kuklospora a Different Genus?
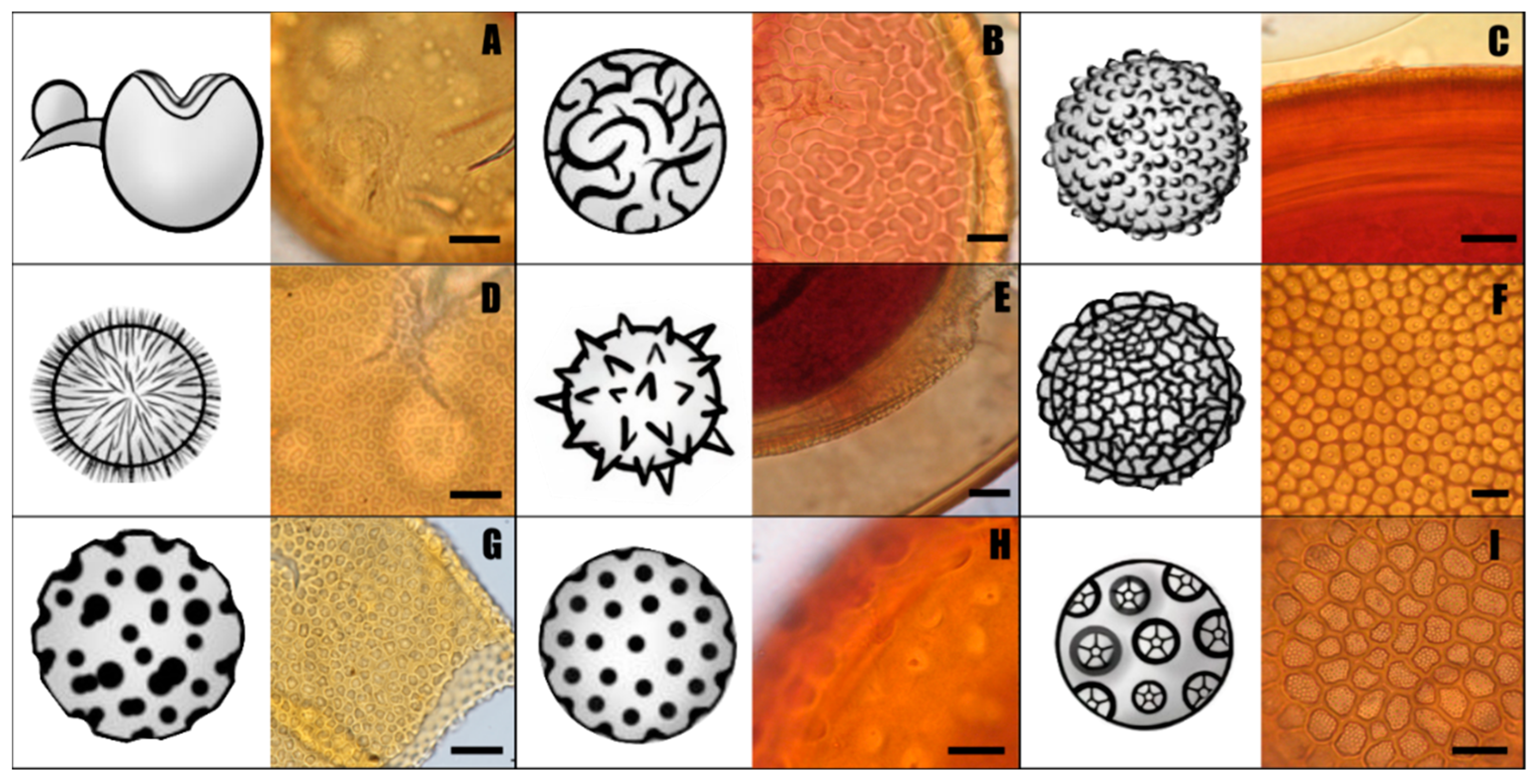
4.2. The Homoplasic Nature of Acaulospora Ornamentations
5. Morphological Characteristics of Acaulospora Species Shared with Other Genera
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schüßler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: San Diego, CA, USA, 2008; p. 787. Available online: https://www.elsevier.com/books/mycorrhizal-symbiosis/smith/978-0-12-370526-6 (accessed on 10 January 2020).
- Baar, J.; Paradi, I.; Lucassen, E.C.; Hudson-Edwards, K.A.; Redecker, D.; Roelofs, J.G.M.; Smolders, A.J.P. Molecular analysis of AMF diversity in aquatic macrophytes: A comparison of oligotrophic and utra-oligotrophic lakes. Aquat. Bot. 2011, 94, 53–61. [Google Scholar] [CrossRef]
- Queiroz, M.B.; Gomes, S.R.B.S.; Vista, X.M.; Leroy, J.A.S.; Goto, B.T. Conhecimento, Conservação e Uso de Fungos; Oliveira, L.A., Jesus, M.A., Matsuura, A.B.J., Oliveira, J.G.S., Gasparotto, L., Lima-Neto, R.G., Rocha, L.C., Eds.; Editora INPA: Manaus, Brazilm, 2019. Available online: https://www.alice.cnptia.embrapa.br/handle/doc/1118871 (accessed on 10 January 2020).
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367, 6480. [Google Scholar] [CrossRef]
- Kivlin, N.S.; Hawkes, C.V.; Treseder, K.K. Global diversity and distribution of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2011, 43, 2294–2303. [Google Scholar] [CrossRef] [Green Version]
- Trejo, D.; Barois, I.; Sangabriel-Conde, W. Disturbance and land use effect on functional diversity of the arbuscular mycorrhizal fungi. Agrofor. Syst. 2016, 90, 265–279. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Oberholzer, H.-R.; Jansa, J.; Egli, S. Diversity and biogeography of arbuscular mycorrhizal fungi in agricultural soils. Biol. Fertil. Soils 2017, 53, 777–797. [Google Scholar] [CrossRef]
- Davison, J.; Moora, M.; Jairus, T.; Vasar, M.; Öpik, M.; Zobel, M. Hierarchical assembly rules in arbuscular mycorrhizal (AM) fungal communities. Soil Biol. Biochem. 2016, 97, 63–70. [Google Scholar] [CrossRef]
- Maia, L.C.; Passos, J.H.; Silva, J.A.; Oehl, F.; Assis, D.M.A. Species diversity of Glomeromycota in Brazilian biomes. Sydowia 2020, 72, 181–205. [Google Scholar] [CrossRef]
- Vieira, L.C.; Silva, D.K.A.; Escobar, I.E.C.; Silva, J.M.; Moura, I.A.; Oehl, F.; Silva, G.A. Changes in an Arbuscular Mycorrhizal Fungi Community Along an Environmental Gradient. Plants 2020, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, J.; Moora, M.; Öpik, M.; Adholeya, A.; Ainsaar, L.; Bâ, A.; Burla, S.; Diedhiou, A.G.; Hiiesalu, I.; Jairus, T.; et al. Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism. Science 2015, 349, 970–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stürmer, S.L.; Bever, J.D.; Morton, J.B. Biogeography of arbuscular mycorrhizal fungi (Glomeromycota): A phylogenetic perspective on species distribution patterns. Mycorrhiza 2018, 28, 587–603. [Google Scholar] [CrossRef]
- Cofré, M.N.; Soteras, F.; Iglesias, M.R.; Velázquez, S.; Abarca, C.; Risio, L.; Ontivero, E.; Cabello, M.N.; Domínguez, L.S.; Lugo, M.A. Biodiversity of arbuscular mycorrhizal fungi in South America: A review. In Mycorrhizal Fungi in South America; Pagano, M.C., Lugo, M.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 49–72. [Google Scholar] [CrossRef]
- Dotzler, N.; Walker, C.; Krings, M.; Hass, H.; Kerp, H.; Taylor, T.N.; Agerer, R. Acaulosporoid glomeromycotan spores with a germination shield from the 400-million-year-old Rhynie chert. Mycol. Prog. 2009, 8, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Jobim, K.; Błaszkowski, J.; Niezgoda, P.; Kozłowska, A.; Zubek, S.; Mleczko, P.; Chachuła, P.; Ishikawa, N.K.; Goto, B.T. New sporocarpic taxa in the phylum Glomeromycota: Sclerocarpum amazonicum gen. et sp. nov. in the family Glomeraceae (Glomerales) and Diversispora sporocarpia sp. nov. in the Diversisporaceae (Diversisporales). Mycol. Prog. 2019, 18, 369–384. [Google Scholar] [CrossRef] [Green Version]
- Błaszkowski, J.; Jobim, K.; Niezgoda, P.; Meller, E.; Malinowski, R.; Milczarski, P.; Zubek, S.; Magurno, F.; Casieri, L.; Bierza, W.; et al. New Glomeromycotan Taxa, Dominikia glomerocarpica sp. nov. and Epigeocarpum crypticum gen. nov. et sp. nov. From Brazil, and Silvaspora gen. nov. From New Caledonia. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-H.; Eo, J.-K.; Ka, K.-H.; Eom, A.-H. Diversity of Arbuscular Mycorrhizal Fungi and Their Roles in Ecosystems. Mycobiology 2013, 41, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Oehl, F.; Sieverding, E.; Palenzuela, J.; Ineichen, K.; Silva, G.A. Advances in Glomeromycota taxonomy and classification. IMA Fungus 2011, 2, 191–199. [Google Scholar] [CrossRef]
- Choia, J.J.; Kim, S.-H. A genome Tree of Life for the Fungi kingdom. Proc. Natl. Acad. Sci. USA 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [Green Version]
- De Souza, F.A.; da Silva, I.R.; Barreto, M.B.B.D.B.; Oehl, F.; Goto, B.T.; Maia, L.C. Racocetra crispa (Glomeromycotina) delimited by integrative evidence based on morphology, long continuous nuclear rDNA sequencing and phylogeny. Mycol. Prog. 2018, 17, 999–1011. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; de Paiva, J.N.; da Silva, K.J.G.; Theodoro, R.C.; Jobim, K.; Orfanoudakis, M.; Goto, B.T. Sieverdingia gen. nov., S. tortuosa comb. nov., and Diversispora peloponnesiaca sp. nov. in the Diversisporaceae (Glomeromycota). Mycol. Prog. 2019, 18, 1363–1382. [Google Scholar] [CrossRef] [Green Version]
- Błaszkowski, J.; Niezgoda, P.; Zubek, S.; Meller, E.; Milczarski, P.; Malicka, M.; Goto, B.T.; Woźniak, G.; Moreira, H.; Magurno, F. Dominikia bonfanteae and Glomus atlanticum, two new species in the Glomeraceae (phylum Glomeromycota) with molecular phylogenies reconstructed from two unlinked loci. Mycol. Prog. 2021, 20, 131–148. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef]
- Gerdemann, J.W.; Trappe, J.M. The Endogonaceae in the Pacific Northwest. Mycol. Mem. 1974, 5, 76. [Google Scholar] [CrossRef] [Green Version]
- Turrini, A.; Giovannetti, M. Arbuscular mycorrhizal fungi in national parks, nature reserves and protected areas worldwide: A strategic perspective for their in situ conservation. Mycorrhiza 2011, 22, 81–97. [Google Scholar] [CrossRef]
- Pereira, C.M.R.; da Silva, D.K.A.; Ferreira, A.C.D.A.; Goto, B.T.; Maia, L.C. Diversity of arbuscular mycorrhizal fungi in Atlantic forest areas under different land uses. Agric. Ecosyst. Environ. 2014, 185, 245–252. [Google Scholar] [CrossRef]
- Welemariam, M.; Kebede, F.; Bedadi, B.; Birhane, E. Effect of community-based soil and water conservation practices on arbuscular mycorrhizal fungi types, spore densities, root colonization, and soil nutrients in the northern highlands of Ethiopia. Chem. Biol. Technol. Agric. 2018, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Yano-Melo, A.M.; Saggin, O.J.; Maia, L.C. Tolerance of mycorrhized banana (Musa sp. cv. Pacovan) plantlets to saline stress. . Agric. Ecosyst. Environ. 2003, 95, 343–348. [Google Scholar] [CrossRef]
- Tian, C.Y.; Feng, G.; Li, X.L.; Zhang, F.S. Different effects of arbuscular mycorrhizal fungal isolates from saline or non-saline soil on salinity tolerance of plants. Appl. Soil Ecol. 2004, 26, 143–148. [Google Scholar] [CrossRef]
- Akib, M.A.; Mustari, K.; Kuswinanti, T.; Syaiful, S.A. The effect of application Acaulospora sp on the root growth of Canavaliaensiformis l at nickel post-mine land. Pak. J. Biotechnol. 2018, 15, 583–591. Available online: https://pjbt.org/index.php/pjbt/article/view/434 (accessed on 10 January 2020).
- Schneider, J. Ocorrência e Ação Fitoprotetora de Fungos Micorrízicos Arbusculares em Solos Contaminados Com Arsênio; UFLA: Lavras, Brazil, 2011; 181p, Available online: http://repositorio.ufla.br/handle/1/3724 (accessed on 10 January 2020).
- Stürmer, S.L.; Kemmelmeier, K. The Glomeromycota in the Neotropics. Front. Microbiol. 2021, 11, 553679. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, A.S.; Goto, B.T.; Ganade, G. Ecological restoration methods influence the structure of arbuscular mycorrhizal fungal communities in degraded drylands. J. Soil Ecol. 2021, 84, 150690. [Google Scholar] [CrossRef]
- Giovannetti, M. Janusz Błaszkowski (ed); Glomeromycota. Mycorrhiza 2013, 23, 251–252. [Google Scholar] [CrossRef]
- Walker, C. Taxonomic concepts in the Endogonaceae: Spore-wall characteristics in species descriptions. Mycotaxon 1983, 18, 443–455. [Google Scholar]
- Goto, B.T.; Maia, L.C. Glomerospores: A new denomination for the spores of Glomeromycota, a group molecularly distinct from the Zygomycota. Mycotaxon 2006, 96, 129–132. [Google Scholar]
- Morton, J.B.; Benny, G.L. Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): A new order Glomales and Gigasporineae and two new families Acaulosporaceae and Gigasporaceae with an emendation of Glomaceae. Mycotaxon 1990, 37, 471–491. [Google Scholar]
- Morton, J.B.; Redecker, D. Two new families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia 2001, 93, 181–195. [Google Scholar] [CrossRef]
- Walker, C.; Vestberg, M.; Demircik, F.; Stockinger, H.; Saito, M.; Sawaki, H.; Nishmura, I.; Schüßler, A. Molecular phylogeny and new taxa in the Archaeosporales (Glomeromycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycol. Res. 2007, 111, 137–153. [Google Scholar] [CrossRef]
- Palenzuela, J.; Ferrol, N.; Boller, T.; Azcón-Aguilar, C.; Oehl, F. Otospora bareai, a new fungal species in the Glomeromycetes from a dolomitic shrub land in Sierra de Baza National Park (Granada, Spain). Mycologia 2008, 100, 296–305. [Google Scholar] [CrossRef]
- Oehl, F.; Sánchez-Castro, I.; Palenzuela, J.; Silva, G.A. Palaeospora spainii, a new arbuscular mycorrhizal fungus from Swiss agricultural soils. Nova Hedwig. 2015, 101, 89–102. [Google Scholar] [CrossRef]
- Ames, R.N.; Schneider, R.W. Entrophospora, a new genus in the Endogonaceae. Mycotaxon 1979, 8, 347–352. [Google Scholar]
- Sieverding, E.; Oehl, F. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. J. Appl. Bot. 2006, 80, 69–81. [Google Scholar]
- Oehl, F.; Da Silva, G.A.; Sánchez-Castro, I.; Goto, B.T.; Maia, L.C.; Vieira, H.E.E.; Barea, J.M.; Sieverding, E.; Palenzuela, J. Revision of Glomeromycetes with entrophosporoid and glomoid spore formation with three new genera. Mycotaxon 2011, 117, 297–316. [Google Scholar] [CrossRef]
- Kaonongbua, W.; Morton, J.B.; Bever, J.D. Taxonomic revision of arbuscular mycorrhizal fungal genus Acaulospora Gerd. & Trappe to include Kuklospora and a description of Acaulospora colliculosa sp. nov. from field collected spores. Mycologia 2010, 102, 1497–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, B.T.; Pereira, C.M.; Nobre, C.P.; Zatorre, N.P.; Covacevich, F.; Berbara, R.L.L.; Maia, L.C. Acaulospora endographis (Glomeromycetes), a new fungus with a complex spore wall. Mycotaxon 2013, 123, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Pawłowska, J.; Letcher, P.M.; Kirk, P.M.; Humber, R.A.; Schüßler, A.; Wrzosek, M.; Muszewska, A.; Okrasińska, A.; Istel, Ł.; et al. Notes for genera: Basal clades of Fungi (including Aphelidiomycota, Basidiobolomycota, Blastocladiomycota, Calcarisporiellomycota, Caulochytriomycota, Chytridiomycota, Entomophthoromycota, Glomeromycota, Kickxellomycota, Monoblepharomycota, Mortierellomyc. Fungal Divers. 2018, 92, 43–129. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA Genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfrand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Oehl, F.; Sykorova, Z.; Redecker, D.; Wiemken, A. Acaulospora alpina, a new arbuscular mycorrhizal fungal species characteristic for high mountainous and alpine regions of the Swiss Alps. Mycologia 2006, 98, 286–294. [Google Scholar] [CrossRef]
- Corazon-Guivin, M.A.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Vallejos-Tapullima, A.; Silva, G.A.; Oehl, F. Acaulospora aspera, a new fungal species in the Glomeromycetes from rhizosphere soils of the inka nut (Plukenetiavolubilis L.) in Peru. J. Appl. Bot. Food Qual. 2019, 92, 250–257. [Google Scholar] [CrossRef]
- Palenzuela, J.; Azcón-Aguilar, C.; Barea, J.M.; Silva, G.A.; Oehl, F. Acaulospora baetica, a new arbuscular mycorrhizal fungal species from two mountain ranges in Andalucía (Spain). Nova Hedwig. 2015, 101, 463–474. [Google Scholar] [CrossRef]
- Rothwell, F.M.; Trappe, J.M. Acaulospora bireticulata sp.nov. Mycotaxon 1979, 8, 471–475. [Google Scholar]
- Goto, B.T.; Maia, L.C.; Oehl, F. Ambispora brasiliensis, a new ornamented species in the arbuscular mycorrhiza forming Glomeromycetes. Mycotaxon 2008, 105, 11–18. [Google Scholar]
- Krüger, M.; Walker, C.; Schüßler, A. Acaulospora brasiliensis comb. nov. and Acaulospora alpina (Glomeromycota) from upland Scotland: Morphology, molecular phylogeny and DNA-based detection in roots. Mycorrhiza 2011, 21, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J. Polish Endogonaceae 7. Acaulospora capsicula, sp. nov. Mycologia 1990, 82, 794–798. [Google Scholar] [CrossRef]
- Błaszkowski, J. Acaulospora cavernata—A new species with pitted spores. Cryptogam. Bot. 1989, 1, 204–207. [Google Scholar]
- Schultz, P.A.; Bever, J.D.; Morton, J.B. Acaulospora colossica sp. nov. from an old field in North Carolina and morphological comparisons with similar species, A. laevis and A. koskei. Mycologia 1999, 91, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.; Pfeiffer, C.M.; Bloss, H.E. Acaulospora delicata sp. nov.: An endomycorrhizal fungus from Arizona. Mycotaxon 1986, 25, 621–628. [Google Scholar]
- Sieverding, E.; Toro, S. Acaulospora denticulata sp. nov. and Acaulospora rehmii sp. nov. (Endogonaceae) with ornamented spore walls. Angew. Bot. 1987, 61, 217–223. [Google Scholar]
- Morton, J.B. Three new species of Acaulospora (Endogonaceae) from high aluminium, low pH soils in West Virginia. Mycologia 1986, 78, 641–648. [Google Scholar] [CrossRef]
- Velazquez, M.S.; Cabello, M.N.; Irrazabal, G.; Godeas, A. Acaulosparaceae from El Palmar National Park, Entre Ríos, Argentina. Mycotaxon 2008, 103, 171–187. [Google Scholar]
- Ingleby, K.; Walker, C.; Mason, P.A. Acaulospora excavata sp. nov.—An endomycorrhizal fungus from Cote d’Ivoire. Mycotaxon 1994, 50, 99–105. [Google Scholar]
- He, R.J.; Long, C.L.; Yao, L.M.; Jiang, L. Acaulospora fanjing (Glomeromycota), a new species of arbuscular mycorrhizal fungi from Fanjingshan in China. Phytotaxa 2021, 524, 191–198. [Google Scholar] [CrossRef]
- Corazon-Guivin, M.A.; Vallejos-Tapullima, A.; Sota-Ricaldi, A.M.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Santos, V.M.; Silva, G.A.; Oehl, F. Acaulospora flava, a new arbuscular mycorrhizal fungus from Coffea arabica and Plukenetia volubilis plantations at the sources of the Amazon river in Peru. J. Appl. Bot. Food Qual. 2021, 94, 116–123. [Google Scholar] [CrossRef]
- Corazon-Guivin, M.A.; Vallejos-Tapullima, A.; Sota-Ricaldi, A.M.; Cerna-Mendoza, A.; Guerrero-Abad, J.C.; Santos, V.M.; da Silva, G.A.; Oehl, F. Acaulospora flavopapillosa, a new fungus in the Glomeromycetes from a coffee plantation in Peru, with an updated key for the identification of Acaulosporaceae species. J. Appl. Bot. Food Qual. 2022, 95, 6–16. [Google Scholar] [CrossRef]
- Janos, D.P.; Trappe, J.M. Two new Acaulospora species from tropical America. Mycotaxon 1982, 15, 515–522. [Google Scholar]
- Crossay, T.; Cilia, A.; Cavaloc, Y.; Amir, H.; Redecker, D. Four new species of arbuscular mycorrhizal fungi (Glomeromycota) associated with endemic plants from ultramafic soils of New Caledonia. Mycol. Prog. 2018, 17, 729–744. [Google Scholar] [CrossRef]
- Błaszkowski, J. Four new species of the Endogonaceae (Zygomycotina) from Poland. Karstenia 1988, 27, 37–42. [Google Scholar] [CrossRef]
- Furrazola, E.; Goto, B.T.; Silva, G.A.; Torres-Arias, Y.; Morais, T.; Lima, C.E.P.; Ferreira, A.C.A.; Maia, L.C.; Sieverding, E.; Oehl, F. Acaulospora herrerae, a new pitted species in the Glomeromycetes from Cuba and Brazil. Nova Hedwig. 2013, 97, 401–413. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Chwat, G.; Góralska, A. Acaulospora ignota and Claroideoglomus hanlinii, two new species of arbuscular mycorrhizal fungi (Glomeromycota) from Brazil and Cuba. Mycol. Prog. 2015, 14, 18. [Google Scholar] [CrossRef]
- Lee, E.-H.; Park, S.-E.; Eo, J.-K.; Ka, K.-H.; Eom, A.-H. Acaulospora koreana, a New Species of Arbuscular Mycorrhizal Fungi (Glomeromycota) Associated with Roots of Woody Plants in Korea. Mycobiology 2018, 46, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Błaszkowski, J. Acaulospora koskei, a new species in Glomales from Poland. Mycol. Res. 1995, 99, 237–240. [Google Scholar] [CrossRef]
- Schenck, N.C.; Spain, J.L.; Sieverding, E.; Howeler, R.H. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 1984, 76, 685–699. [Google Scholar] [CrossRef]
- Oehl, F.; Palenzuela, J.; Sánchez-Castro, I.; Hountondji, F.; Tchabi, A.; Louis, L.; Barea, J.M.; Danny, C.; Silva, G.A. Acaulospora minuta, a new arbuscular mycorrhizal fungal species from sub-Saharan savannas of West Africa. J. Appl. Bot. Food Qual. 2011, 84, 213–218. [Google Scholar]
- Oehl, F.; Palenzuela, J.; Sánchez-Castro, I.; Kuss, P.; Sieverding, E.; Silva, G.A. Acaulospora nivalis, a new fungus in the Glomeromycetes, characteristic for high alpine and nival altitudes of the Swiss Alps. Nova Hedwig. 2012, 95, 122. [Google Scholar] [CrossRef]
- Pereira, C.M.; Maia, L.C.; Sánchez-Castro, I.; Palenzuela, J.; Silva, G.A.; Sudová, R.; Sýkorová, Z.; Rydlová, J.; Čtvrtlíková, M.; Goto, B.T.; et al. Acaulospora papillosa, a new mycorrhizal fungus from NE Brazil, and Acaulospora rugosa from Norway. Phytotaxa 2016, 260, 14–24. [Google Scholar] [CrossRef]
- Błaszkowski, J. Three new vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Poland. Bull. Pol. Acad. Sci. Biol. 1988, 36, 271–275. [Google Scholar]
- Oehl, F.; da Silva, G.A.; Palenzuela, J.; Sánchez-Castro, I.; Castillo, C.; Sieverding, E. Acaulospora punctata, a new fungal species in the Glomeromycetes from mountainous altitudes of the Swiss Alps and Chilean Andes. Nova Hedwig. 2011, 93, 353–362. [Google Scholar] [CrossRef]
- Palenzuela, J.; Azcón-Aguilar, C.; Barea, J.M.; Silva, G.A.; Oehl, F. Acaulospora pustulata and Acaulospora tortuosa, two new species in the Glomeromycota from Sierra Nevada (southern Spain). Nova Hedwig. 2013, 97, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.M.R.; Goto, B.T.; da Silva, D.K.A.; Ferreira, A.C.d.; de Souza, F.A.; da Silva, G.A.; Maia, L.C.; Oehl, F. Acaulospora reducta sp. nov. and A. excavata—Two glomeromycotan fungi with pitted spores from Brazil. Mycotaxon 2015, 130, 983–995. [Google Scholar] [CrossRef]
- Trappe, J.M. Three new Endogonaceae: Glomus constrictus, Sclerocystis clavispora and Acaulospora scrobiculata. Mycotaxon 1977, 6, 359–366. [Google Scholar]
- Vaingankar, J.D.; Rodrigues, B.F. Acaulospora soloidea, a new arbuscular mycorrhizal fungus from rhizosphere soils of Murraya paniculata. Mycotaxon 2011, 115, 323–326. [Google Scholar] [CrossRef]
- Walker, C.; Trappe, J.M. Acaulospora spinosa sp. nov. with a key to the species of Acaulospora. Mycotaxon 1981, 12, 515–521. [Google Scholar]
- Pontes, J.S.; Santos, V.M.; Pereira, C.D.; Silva, G.A.; Maia, L.C.; Oehl, F. Acaulospora spinulifera, a new arbuscular mycorrhizal fungal species from the Brazilian Cerrado and Atlantic Rain forest. Nova Hedwig. 2017, 105, 219–229. [Google Scholar] [CrossRef]
- Sieverding, E.; Chaverri, A.; Rojas, I. Acaulospora splendida, a new species in the Endogonaceae from Costa Rica. Mycotaxon 1988, 33, 251–256. [Google Scholar]
- Berch, S.M. Acaulospora sporocarpia, a new sporocarpic species and emendation of the genus Acaulospora (Endogonaceae, Zygomycotina). Mycotaxon 1985, 23, 409–418. [Google Scholar]
- Hu, H.T. Study on the endomycorrhizae of China fir (Cunnighamia lanceolata Hooker) and taiwania (Taiwania cryptomerioides). Quart. J. Chin. Forest. 1988, 21, 45–72. [Google Scholar]
- Lin, T.-C.; Silva, G.A.; Oehl, F. Acaulospora tsugae, a new species in the Glomeromycetes from Taiwan, and a key to species in Acaulosporaceae. Nova Hedwig. 2019, 108, 475–488. [Google Scholar] [CrossRef]
- Swarupa, R.S.; Kunwar, I.K.; Manoharachary, C. Acaulospora terricola sp. nov. from India. J. Mycol. Plant Pathol. 2003, 33, 146–147. [Google Scholar]
- Palenzuela, J.; Azcón-Aguilar, C.; Barea, J.M.; Silva, G.A.; Oehl, F. Acaulospora viridis, a new species in the Glomeromycetes from two moutain ranges in Andalucía (Spain). Nova Hedwig. 2014, 99, 71–82. [Google Scholar] [CrossRef]
- Kramadibrata, K.; Hedger, J.N. A new species of Acaulospora associated with cocoa in Java and Bali (Indonesia). Mycotaxon 1990, 37, 73–77. [Google Scholar]
- Cai, B.P.; Guo, L.D.; Chen, J.Y.; Zhang, Q.X. Glomus mume and Kuklospora spinosa: Two new species of Glomeromycota from China. Mycotaxon 2013, 124, 263–268. [Google Scholar] [CrossRef]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The online database MaarjAM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jaroszewski, L.; Godzik, A. Clustering of highly homologous sequences to reduce the size of large protein databases. Bioinformatics 2001, 17, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jaroszewski, L.; Godzik, A. Tolerating some redundancy significantly speeds up clustering of large protein databases. Bioinformatics 2002, 18, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 20 October 2020).
- European Molecular Biology Laboratory—European Bioinformatics Institute. Available online: https://www.ebi.ac.uk/ (accessed on 20 October 2020).
- Global Biodiversity Information Facility. Available online: https://www.gbif.org/search?q=Acaulospora (accessed on 20 October 2020).
- Barcode of Life Data System. Available online: http://boldsystems.org/index.php (accessed on 20 October 2020).
- Maar, J.A.M. Available online: https://maarjam.botany.ut.ee (accessed on 20 October 2020).
- MYCOBANK Database. Available online: https://www.mycobank.org/page/Basic%20names%20search (accessed on 20 October 2020).
- Furrazola, E. Research on arbuscular mycorrhizae in Cuba: A historical review and future perspectives. Stud. Fungi 2021, 6, 240–262. [Google Scholar] [CrossRef]
- Friberg, S. Distribution and diversity of arbuscular mycorrhizal fungi in traditional agriculture on the Níger inland delta, Mali, west, Africa. CBMs Skr. 2001, 3, 53–80. [Google Scholar]
- Kariman, K.H.; Goltapeh, E.M.; Minassian, V. Arbuscular mycorrhizal fungi from Iran. J. Agric. Technol. 2005, 1, 301–313. [Google Scholar]
- Gupta, M.M.; Naqvi, N.S.; Singh, V.K. The state of arbuscular mycorrhizal fungal diversity in India: An analysis. Sydowia 2014, 66, 265–288. [Google Scholar] [CrossRef]
- Nafady, N.A.; Abdel-Azeem, A.M.; Salem, F.M. A checklist of Egyptian fungi: II. Glomeromycota. Microb. Biosyst. 2016, 1, 40–49. [Google Scholar] [CrossRef]
- Sudová, R.; Kohout, P.; Rydlová, J.; Čtvrtlíková, M.; Suda, J.; Voriskova, J.; Kolaříková, Z. Diverse fungal communities associated with the roots of isoetid plants are structured by host plant identity. Fungal Ecol. 2020, 45, 100914. [Google Scholar] [CrossRef]
- Yang, H.; Dai, Y.; Xu, M.; Zhang, Q.; Bian, X.; Tang, J.; Chen, X. Metadata-mining of 18S rDNA sequences reveals that “everything is not everywhere” for glomeromycotan fungi. Ann. Microbiol. 2015, 66, 361–371. [Google Scholar] [CrossRef]
- Marinho, F.; da Silva, I.R.; Oehl, F.; Maia, L.C. Checklist of arbuscular mycorrhizal fungi in tropical forests. Sydowia 2018, 70, 107–127. [Google Scholar] [CrossRef]
- Pereira, C.M.R.; García, A.L.; da Silva, D.K.A.; Maia, L.C.; Frøslev, T.G.; Kjøller, R.; Rosendahl, S. Tropical Forest type influences community assembly processes in arbuscular mycorrhizal fungi. J. Biogeogr. 2019, 47, 434–444. [Google Scholar] [CrossRef]
- Bruns, T.D.; John, W.; Taylor, J.W. Comment on “Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism”. Sci. Transl. Med. 2016, 351, 826. [Google Scholar] [CrossRef] [Green Version]
- Gladieux, P.; Guérin, F.; Giraud, T.; Caffier, V.; Lemaire, C.; Parisi, L.; Didelot, F.; LE Cam, B. Emergence of novel fungal pathogens by ecological speciation: Importance of the reduced viability of immigrants. Mol. Ecol. 2011, 20, 4521–4532. [Google Scholar] [CrossRef] [Green Version]
- Krüger, M.; Stockinger, H.; Krüger, C.; Schüßler, A. DNA-based species level detection of Glomeromycota: One PCR primer set for all arbuscular mycorrhizal fungi. New Phytol. 2009, 183, 212–223. [Google Scholar] [CrossRef]
- Husband, R.; Herre, E.A.; Turner, S.L.; Gallery, R.; Young, J.P.W. Molecular diversity of arbuscular mycorrhizal fungi and patterns of host association over time and space in a tropical forest. Mol. Ecol. 2002, 11, 2669–2678. [Google Scholar] [CrossRef] [Green Version]
- Montoliu-Nerin, M.; Sánchez-García, M.; Bergin, C.; Kutschera, V.E.; Johannesson, H.; Bever, J.D.; Rosling, A. In-depth Phylogenomic Analysis of Arbuscular Mycorrhizal Fungi Based on a Comprehensive Set of de novo Genome Assemblies. Front. Fungal Biol. 2021, 2, 716385. [Google Scholar] [CrossRef]
- Beaudet, D.; Chen, E.C.H.; Mathieu, S.; Yildirir, G.; Ndikumana, S.; Dalpé, Y.; Séguin, S.; Farinelli, L.; Stajich, J.E.; Corradi, N. Ultra-low input transcriptomics reveal the spore functional content and phylogenetic affiliations of poorly studied arbuscular mycorrhizal fungi. DNA Res. 2017, 25, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree, Version v.1.4.3; Computer Program Distributed by the Author. Institute of Evolutionary Biology, University of Edinburgh. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/.1.4.3 (accessed on 20 October 2020).
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Matthew, C.; Fisher, M. Phylogenetic Species Recognition and Species Concepts in Fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, M.; Krüger, C.; Walker, C.; Stockinger, H.; Schüßler, A. Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level. New Phytol. 2012, 193, 970–984. [Google Scholar] [CrossRef] [PubMed]
- Schüßler, A.; Walker, C. Archaeospora ecuadoriana sp. nov. from a mountainous biodiversity hotspot area in Ecuador, and transfer of Palaeospora spainiae to Archaeospora, as A. spainiae comb. nov. Mycorrhiza 2019, 29, 435–443. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Reference Number |
|---|---|
| Acaulospora alpina | UFRN-Fungos 3408 |
| Acaulospora ignota | UFRN-Fungos 3409 |
| Acaulospora denticulata | UFRN-Fungos 3410 |
| Acaulospora excavata | UFRN-Fungos 3411 |
| Acaulospora elegans | UFRN-Fungos 3412 |
| Acaulospora foveata | UFRN-Fungos 3413 |
| Acaulospora herrerae | UFRN-Fungos 3414 |
| Acaulospora laevis | UFRN-Fungos 3415 |
| Acaulospora lacunosa | UFRN-Fungos 3416 |
| Acaulospora mellea | UFRN-Fungos 3417 |
| Acaulospora morrowiae | UFRN-Fungos 3418 |
| Acaulospora reducta | UFRN-Fungos 3419 |
| Acaulospora rugosa | UFRN-Fungos 3420 |
| Acaulospora scrobiculata | UFRN-Fungos 3421 |
| Acaulospora spinosa | UFRN-Fungos 3422 |
| Acaulospora spinosissima | UFRN-Fungos 2965 |
| Acaulospora sporocarpia | OSC, Oregon, 47836 1 |
| Acaulospora tuberculata | UFRN-Fungos 3423 |
| Species | Ornate | Type | Shape | Original Description |
|---|---|---|---|---|
| Acaulospora alpina | Yes | Depressions | Pits | regular conical pits |
| Acaulospora aspera | Yes | Depressions | Pits | irregular depressions |
| Acaulospora baetica | Yes | Depressions | Pits | pitted |
| Acaulospora bireticulata | Yes | Double depressions | Depressed with central stratum | polygonal reticulum depressed central stratum; ridges occasionally branched forming irregular isolated projections |
| Acaulospora brasiliensis | Yes | Projections | Pustules | convex pustules irregular shape and size |
| Acaulospora capsicula | No | --- | --- | --- |
| Acaulospora cavernata | Yes | Depressions | Pits | evenly pitted deep depressions separated by ridges |
| Acaulospora colliculosa | Yes | Projections | Protuberances | protuberances |
| Acaulospora colombiana | No | --- | --- | --- |
| Acaulospora colossica | No | --- | --- | --- |
| Acaulospora delicata | No | --- | --- | --- |
| Acaulospora denticulata | Yes | Projections | Teeth | tooth-shaped projections circular or oblong |
| Acaulospora dilatata | No | --- | --- | --- |
| Acaulospora endographis | Yes | Projections | Spines | irregular spines |
| Acaulospora elegans | Yes | Projections | Spines | ornamented with crowded densely organized spines |
| Acaulospora entreriana | Yes | Projections | Teeth | ornamented with teeth |
| Acaulospora excavata | Yes | Depressions | Pits | concave round pits |
| Acaulospora fanjing | No | --- | --- | --- |
| Acaulospora flava | No | --- | --- | --- |
| Acaulospora flavopapillosa | Yes | Projections | Papillae | fine papillae |
| Acaulospora foveata | Yes | Depressions | Pits | round to oblong and concave depressions |
| Acaulospora fragilissima | No | --- | --- | --- |
| Acaulospora gedanensis | No | --- | --- | --- |
| Acaulospora herrerae | Yes | Depressions | Pits | rounded to elliptical pits some pits vermiform or regulate |
| Acaulospora ignota | Yes | Projections | Excrescences | granular excrescences |
| Acaulospora kentinensis | Yes | Depressions | Pits | pits circular to subcircular deep when observed in cross view |
| Acaulospora koreana | No | --- | --- | --- |
| Acaulospora koskei | No | --- | --- | --- |
| Acaulospora lacunosa | Yes | Depressions | Pits | irregularly distributed irregular saucer-shaped pits |
| Acaulospora laevis | No | --- | --- | --- |
| Acaulospora longula | No | --- | --- | --- |
| Acaulospora mellea | No | --- | --- | --- |
| Acaulospora minuta | Yes | Depressions | Pits | minute pit-like depressions |
| Acaulospora morrowiae | No | --- | --- | --- |
| Acaulospora nivalis | Yes | Depressions | Pits | irregular pits |
| Acaulospora papillosa | No | --- | --- | --- |
| Acaulospora paulinae | Yes | Depressions | Pits | concave round pits of widest diameter |
| Acaulospora punctata | Yes | Depressions | Pits | regular round pits |
| Acaulospora pustulata | Yes | Projections | Blister | pustulate projections |
| Acaulospora reducta | Yes | Depressions | Pits | irregularly-shaped small pits sometimes dumbbell-shaped pits |
| Acaulospora rehmii | Yes | Projections | Cerebriform | cerebriform folds |
| Acaulospora rugosa | No | --- | --- | --- |
| Acaulospora saccata | No | --- | --- | --- |
| Acaulospora scrobiculata | Yes | Depressions | Pits | ornamented with evenly distributed pits, circular ellipsoidal oblong triangular Y-shaped to irregular |
| Acaulospora sieverdingii | Yes | Depressions | Pits | irregular pits |
| Acaulospora soloidea | Yes | Projections | Bristle | ornamented with numerous acellular fibrillose hairy outgrowths forming a pile or thick coat |
| Acaulospora spinosa | Yes | Projections | Spines | densely organized spines |
| Acaulospora spinosissima | Yes | Projections | Spines | short spiny projections |
| Acaulospora spinulifera | Yes | Projections | Spines | fine spines |
| Acaulospora splendida | No | --- | --- | --- |
| Acaulospora sporocarpia | No | --- | --- | --- |
| Acaulospora taiwania | Yes | Depressions | Pits | side pits ridges form mesh |
| Acaulospora tsugae | No | --- | --- | --- |
| Acaulospora terricola | Yes | Depressions | Pits | minutely pitted |
| Acaulospora tortuosa | Yes | Projections | Excrescences | tortuous hyphae-like structures on the surface |
| Acaulospora thomii | No | --- | --- | --- |
| Acaulospora tuberculata | Yes | Projections | Spines/Tubercles | ornamented with evenly spines or tubercles |
| Acaulospora viridis | No | --- | --- | --- |
| Acaulospora verna | Yes | Depressions | Pits | ornamented with evenly distributed pits, circular to subcircular frequently ellipsoidal to oblong sometimes irregular |
| Acaulospora walkeri | Yes | Projections | Excrescences | finely ornamented |
| Kuklospora spinosa | Yes | Projections | Spines | fine spines |
| Original Name | Description | Current Name | Description | Authors |
|---|---|---|---|---|
| ≡Acaulospora appendicula | 1984 | Ambispora appendicula | 2008 | (Spain, Sieverd. & N.C. Schenck) C. Walker |
| ≡Acaulospora gerdemannii | 1979 | Ambispora jimgerdemannii | 2008 | (Spain, Oehl & Sieverd.) C. Walker |
| ≡Acaulospora myriocarpa | 1986 | Archaeospora myriocarpa | 2011 | (Spain, Sieverd. & N.C. Schenck) Oehl, G.A. Silva, B.T. Goto & Sieverd. |
| ≡Acaulospora nicolsonii | 1984 | Ambispora nicolsonii | 2012 | (C. Walker, L.E. Reed & F.E. Sanders) Oehl, G.A. Silva, B.T. Goto & Sieverd. |
| ≡Acaulospora trappei | 1976 | Archaeospora trappei | 2001 | (R.N. Ames & Linderman) J.B. Morton & D. Redecker |
| ≡Acaulospora undulata | 1988 | Archaeospora undulata | 2011 | (Sieverd.) Sieverd., G.A. Silva, B.T. Goto & Oehl |
| ≡Ambispora brasiliensis | 2008 | Acaulospora brasiliensis | 2011 | (B.T. Goto, L.C. Maia & Oehl) C. Walker, Krüger & Schüßler |
| ≡Acaulospora polonica | 1988 | Polonospora polonica | 2021 | (Błaszk.) Błaszk., Niezgoda, B.T. Goto & Magurno |
| N° | AMF Species | Occurrence | Sequence | Reference |
|---|---|---|---|---|
| 1 | Acaulospora alpina Oehl, Sýkorová & Sieverd. | England, Switzerland, India, Brazil | SSU-ITS-LSU | [52] |
| 2 | Acaulospora aspera Corazon-Guivin, Oehl & G.A. Silva | Peru | SSU-ITS-LSU | [53] |
| 3 | Acaulospora baetica Palenz., Oehl, Azcón-Aguilar & G.A. Silva. | Spain, Brazil | SSU-ITS-LSU | [54] |
| 4 | Acaulospora bireticulata F.M. Rothwell & Trappe | Brazil, Argentina, England, South Korea, Poland, Egypt, India, United States, China, Italy | --- | [37,55] |
| 5 | Acaulospora brasiliensis (B.T. Goto, L.C. Maia & Oehl) C. Walker, Krüger & Schüßler | Brazil, Scotland, Argentina, South Korea | SSU-ITS-LSU | [56,57] |
| 6 | Acaulospora capsicula Błaszk. | Australia, England, Poland, China, Brazil, Egypt, India, United States, Switzerland | --- | [58] |
| 7 | Acaulospora cavernata Błaszk. | Poland, Brazil, Benin, China, Switzerland | SSU-ITS-LSU | [59] |
| 8 | Acaulospora colliculosa Kaonongbua, J.B. Morton & Bever | United States, England | LSU | [48] |
| 9 | Acaulospora colombiana (Spain & N.C. Schenck) Kaonongbua, J.B. Morton & Bever | Colombia, Brazil, India, Philippines, Benin, Germany, Switzerland | SSU-ITS-LSU; ORF1 gene cox1; Beta Tubulin; gene CHS | [48] |
| 10 | Acaulospora colossica P.A. Schultz, Bever & J.B. Morton | United States, Brazil | SSU 1-ITS | [60] |
| 11 | Acaulospora delicata C. Walker, C.M. Pfeiffer & Bloss | Australia, Argentina, Brazil, China, United States, Spain, Philippines, Indonesia, Mexico, England, Senegal, Venezuela, Poland, India, Iceland | SSU-ITS-LSU | [61] |
| 12 | Acaulospora denticulata Sieverd. & S. Toro | Argentina, Colombia, Mexico, Brazil, South Korea, South Africa, China, India, Canada, Kenya | ITS | [62] |
| 13 | Acaulospora dilatata J.B. Morton | Argentina, United States, New Zealand, South Atlantic, Venezuela, Brazil, India | SSU; LSU; Beta tubulin | [63] |
| 14 | Acaulospora endographis B.T. Goto | Brazil | --- | [49] |
| 15 | Acaulospora elegans Trappe & Gerd. | United States, Indonesia, Poland, Brazil, South Korea, Argentina, India, South Africa, Switzerland | LSU | [27] |
| 16 | Acaulospora entreriana M.S. Velázquez & Cabello | Argentina, Brazil | SSU-ITS-LSU | [64] |
| 17 | Acaulospora excavata Ingleby & C. Walker | Australia, Argentina, Ivory Coast, Spain, New Zealand, Trinidad and Tobago, Brazil | ITS | [65] |
| 18 | Acaulospora fanjing R.J. He, L.M. Yao & L. Jiang | China | SSU-ITS-LSU | [66] |
| 19 | Acaulospora flava Corazon-Guivin, G.A. Silva & Oehl | Peru | SSU-ITS-LSU | [67] |
| 20 | Acaulospora flavopapillosa Corazon-Guivin, G.A. Silva & Oehl | Peru | SSU-ITS-LSU | [68] |
| 21 | Acaulospora foveata Trappe & Janos | Australia, Brazil, Costa Rica, Mexico, England, Congo, Poland, Panama, Indonesia, South Korea, Argentina, India | SSU-ITS-LSU | [69] |
| 22 | Acaulospora fragilissima D. Redecker, Crossay & Cilia | New Caledonia, Peru | SSU-ITS-LSU; Beta tubulin; ORF1 gene cox1 | [70] |
| 23 | Acaulospora gedanensis Błaszk. | Poland, Brazil, Austria, Switzerland | --- | [71] |
| 24 | Acaulospora herrerae Furrazola, B.T. Goto, G.A. Silva, Sieverd. & Oehl | Brazil, Cuba, Peru | LSU | [72] |
| 25 | Acaulospora ignota Błaszk., Góralska, Chwat & B.T. Goto | Brazil | SSU-ITS-LSU | [73] |
| 26 | Acaulospora kentinensis (Wu & Liu) Oehl & Sieverd. | Taiwan, Brazil, Benin | SSU-ITS-LSU; ORF1 gene cox1; Beta Tubulin | [48] |
| 27 | Acaulospora koreana E. H. Lee, S. H. Park & A. H | North Korea | SSU-ITS-LSU | [74] |
| 28 | Acaulospora koskei Błaszk. | Australia, Spain, Poland, Brazil, Estonia, Egypt, India, Cameroon, England | SSU-ITS-LSU; Beta tubulin | [75] |
| 29 | Acaulospora lacunosa J.B. Morton | Argentina, United States, New Zealand, Venezuela, Brazil, Poland, Denmark, India, Kenya, China | SSU-ITS-LSU; Beta tubulin; HSP60 gene | [63] |
| 30 | Acaulospora laevis Gerd. & Trappe | Australia, Argentina, China, Ecuador, United States, New Zealand, England, Brazil, South Korea, Egypt, India, Mexico, Iceland, France, Germany, Greece, Benin, Switzerland, Israel, South Africa | SSU-ITS-LSU; Beta tubulin; RPB1; mRNA gene Ste12; Alfa tubulin;ef1 Alpha | [27] |
| 31 | Acaulospora longula Spain & N.C. Schenck | Australia, Germany, Brazil, Colombia, Spain, Philippines, Japan, Indonesia, England, Uganda, Venezuela, South Korea, India, Switzerland | SSU; LSU; RPB1 | [76] |
| 32 | Acaulospora mellea Spain & N.C. Schenck | Germany, Argentina, Brazil, Colombia, China, United States, Mexico, Poland, India, Canada, Denmark, Benin, Cameroon, Switzerland | SSU-ITS-LSU; Beta tubulin; ORF1 gene cox1 | [76] |
| 33 | Acaulospora minuta Oehl, Tchabi, Hount., Palenz., I.C. Sánchez & G.A. Silva | Benin, Brazil | SSU-ITS-LSU | [77] |
| 34 | Acaulospora morrowiae Spain &N.C. Schenck | Colombia, United States, Mexico, Brazil, Poland, South Korea, Namibia, India, Niger, Mali, France, Germany, Switzerland, Benin, Cameroon, China | SSU; LSU; SSU-ITS; ITS-LSU 1; Beta tubulin; glomalin cDNA; HSP60 gene | [76] |
| 35 | Acaulospora nivalis Oehl, Palenz., I.C. Sánchez, G.A. Silva & Sieverd. | Switzerland, Brazil | SSU-ITS-LSU | [78] |
| 36 | Acaulospora papillosa C.M.R. Pereira & Oehl | Brazil, Norway | SSU-ITS-LSU | [79] |
| 37 | Acaulospora paulinae Błaszk. | Brazil, Poland, United States, India, Germany, France, Switzerland, Israel | ITS2-LSU; SSU-ITS1; SSU-ITS | [80] |
| 38 | Acaulospora punctata Oehl, Palenz., I.C. Sánchez, G.A. Silva, C. Castillo & Sieverd. | Brazil, Switzerland, Chile | SSU-ITS-LSU | [81] |
| 39 | Acaulospora pustulata Palenz., Oehl, Azcon-Aguilar & G.A. Silva | Spain, Switzerland | SSU-ITS-LSU | [82] |
| 40 | Acaulospora reducta Oehl, B.T. Goto & C.M.R. Pereira | Brazil | SSU 1-ITS-LSU 1 | [83] |
| 41 | Acaulospora rehmii Sieverd. & S. Toro | Brazil, Mexico, South Korea, Argentina, Poland, Colombia, Taiwan, Egypt, India, China | --- | [62] |
| 42 | Acaulospora rugosa J.B. Morton | Brazil, United States, South Korea, Poland, India, Iran | SSU-ITS-LSU | [63] |
| 43 | Acaulospora saccata D. Redecker, Crossay & Cilia | New Caledonia | SSU-ITS-LSU | [70] |
| 44 | Acaulospora scrobiculata Trappe | Brazil, England, Mexico, Australia, Indonesia, Trinidad and Tobago, Spain, Argentina, Venezuela, United States, Thailand, Japan, Philippines, Poland, Canada, Israel, Greece, Italy, Cameroon, China, Taiwan, South Korea, India, Benin, Finland, Kenya, Switzerland, Nepal | SSU-ITS-LSU; Beta tubulin; gene CHS | [84] |
| 45 | Acaulospora sieverdingii Oehl, Sýkorová & Błaszk. | Brazil, Germany, Poland, Italy, Northern Guinea, Southern Guinea, Benin, France, Sudan, Switzerland | ITS | [47] |
| 46 | Acaulosporasoloidea Vaingankar & B.F. Rodrigues | India, Egypt | --- | [85] |
| 47 | Acaulospora spinosa C. Walker & Trappe | United States, Mexico, Argentina, Brazil, South Korea, Ecuador, China, India, Canada, Venezuela, Colombia, Benin, Cameroon, Kenya, South Africa, Switzerland, Nepal | SSU-ITS-LSU | [86] |
| 48 | Acaulospora spinosissima Oehl, Palenz., Sánchez-Castro, Tchabi, Hount. & G. A. Silva | Sudan, South Africa, Northern Guinea, Southern Guinea, Switzerland, Benin, Brazil | SSU-ITS-LSU | [53] |
| 49 | Acaulospora spinulifera Oehl, V.M. Santos, J.S. Pontes & G.A. Silva | Brazil | LSU | [87] |
| 50 | Acaulospora splendida Sieverd., Chaverri & I. Rojas | Costa Rica, India, Mexico, Egypt, Brazil | --- | [88] |
| 51 | Acaulospora sporocarpia S.M. Berch | England, Switzerland, United States, Pakistan, Egypt, India, Brazil | --- | [89] |
| 52 | Acaulospora taiwania H.T. Hu | Taiwan | --- | [90] |
| 53 | Acaulospora tsugae T.C.Lin & Oehl | Taiwan, Switzerland | SSU 1-ITS-LSU 1 | [91] |
| 54 | Acaulospora terricola Swarupa, Kunwar & Manohar | India | --- | [92] |
| 55 | Acaulospora tortuosa Palenz., Oehl, Azcon-Aguilar & G.A.Silva | Spain, Switzerland | SSU-ITS-LSU | [82] |
| 56 | Acaulospora thomii Błaszk. | Poland, Egypt, India, Switzerland | --- | [71] |
| 57 | Acaulospora tuberculata Janos & Trappe | Brazil, Venezuela, Argentina, Costa Rica, Panama, China, Egypt, India | LSU | [69] |
| 58 | Acaulospora viridis Palenz., Oehl, Azcón-Aguilar & G.A.Silva | Spain | SSU-ITS-LSU | [93] |
| 59 | Acaulospora verna Błaszk. | Poland | --- | [37] |
| 60 | Acaulospora walker Kramad. & Hedger | Australia, Indonesia, Brazil | --- | [94] |
| Kuklospora spinosa B.P. Cai, Jun Y. Chen, Q.X. Zhang & L.D. Guo | China | --- | [95] |
| Access Number Sequence | Concatenated Analysis | SSU-ITS-LSU | ITS1-5.8S-ITS2 |
|---|---|---|---|
| KX355819_Sacculospora_baltica | x | x | x |
| KX355818_Sacculospora_baltica | x | x | x |
| KX355821_Sacculospora_baltica | x | x | x |
| KX345938_Sacculospora_felinovii | x | x | x |
| KX345939_Sacculospora_felinovii | x | x | x |
| KX345941_Sacculospora_felinovii | x | x | x |
| FR681927_Acaulospora_alpina | x | x | x |
| FR681928_Acaulospora_alpina | x | x | x |
| FR681930_Acaulospora_alpina | x | x | x |
| MN080998_Acaulospora_aspera | x | x | x |
| MN081001_Acaulospora_aspera | x | x | x |
| MN080999_Acaulospora_aspera | x | x | x |
| LN810999_Acaulospora_baetica | x | x | x |
| LN811001_Acaulospora_baetica | x | x | x |
| LN811002_Acaulospora_baetica | x | x | x |
| FN825910_Acaulospora_brasiliensis | x | x | x |
| FR681934_Acaulospora_brasiliensis | x | x | x |
| FR681933_Acaulospora_brasiliensis | x | x | x |
| FM876789_Acaulospora_cavernata | x | x | x |
| FM876790_Acaulospora_cavernata | x | x | x |
| FM876788_Acaulospora_cavernata | x | x | x |
| AF133764_Acaulospora_colossica | x | x | x |
| AF133768_Acaulospora_colossica | x | x | x |
| AF133776_Acaulospora_colossica | x | x | x |
| GU326339_Acaulospora_colliculosa | x | ||
| GU326346_Acaulospora_colliculosa | x | ||
| GU326352_Acaulospora_colliculosa | x | ||
| FR750063_Acaulospora_colombiana | x | x | x |
| FJ461804_Acaulospora_colombiana | x | ||
| KX168435_Acaulospora_colombiana | x | ||
| JF439093_Acaulospora_delicata | x | x | x |
| JF439203_Acaulospora_delicata | x | x | x |
| MT832212_Acaulospora_dilatata | x | ||
| FJ461792_Acaulospora_dilatata | x | ||
| AJ239115_Acaulospora_denticulata | x | x | |
| MT112118_Acaulospora_denticulata | x | x | |
| FR750173_Acaulospora_entreriana | x | x | x |
| FR750171_Acaulospora_entreriana | x | x | x |
| FR750169_Acaulospora_entreriana | x | x | x |
| KM057069_Acaulospora_excavata | x | x | |
| KM057074_Acaulospora_excavata | x | x | |
| KM057076_Acaulospora_excavata | x | x | |
| KY362433_Acaulospora_fragilissima | x | x | x |
| KY362432_Acaulospora_fragilissima | x | x | x |
| KY362431_Acaulospora_fragilissima | x | x | x |
| LN736022_Acaulospora_foveata | x | x | x |
| LN736026_Acaulospora_foveata | x | x | x |
| LN736025_Acaulospora_foveata | x | ||
| JX135571_Acaulospora_herrerae | x | ||
| JX135569_Acaulospora_herrerae | x | ||
| JX135573_Acaulospora_herrerae | x | ||
| KP191468_Acaulospora_ignota | x | x | x |
| KP191471_Acaulospora_ignota | x | x | x |
| KP191472_Acaulospora_ignota | x | x | x |
| FM876830_Acaulospora_kentinensis | x | x | x |
| FM876822_Acaulospora_kentinensis | x | x | x |
| FN547520_Acaulospora_kentinensis | x | x | x |
| KP191475_Acaulospora_koskei | x | x | x |
| KP191474_Acaulospora_koskei | x | x | x |
| KP191476_Acaulospora_koskei | x | x | x |
| KY565428_Acaulospora_koreana | x | x | x |
| KY565427_Acaulospora_koreana | x | x | x |
| KY565429_Acaulospora_koreana | x | x | x |
| KP756438_Acaulospora_laevis | x | x | x |
| KP756447_Acaulospora_laevis | x | x | x |
| FN547512_Acaulospora_laevis | x | x | x |
| KP756427_Acaulospora_lacunosa | x | x | x |
| KP756435_Acaulospora_lacunosa | x | x | x |
| KP756584_Acaulospora_lacunosa | x | x | x |
| AM040291_Acaulospora_longula | x | ||
| AM040292_Acaulospora_longula | x | ||
| AJ510228_Acaulospora_longula | x | ||
| AF389007_Acaulospora_longula | x | ||
| KP756453_Acaulospora_mellea | x | x | x |
| KP756456_Acaulospora_mellea | x | x | x |
| KP756471_Acaulospora_mellea | x | x | x |
| FR869691_Acaulospora_minuta | x | x | x |
| FR821675_Acaulospora_minuta | x | x | x |
| FR821674_Acaulospora_minuta | x | x | x |
| AJ242500_Acaulospora_morrowiae | x | x | |
| HE603641_Acaulospora_nivalis | x | x | x |
| HE603643_Acaulospora_nivalis | x | x | x |
| HE603644_Acaulospora_nivalis | x | x | x |
| AJ891120_Acaulospora_paulinae | x | x | |
| AJ891119_Acaulospora_paulinae | x | x | |
| AY639265_Acaulospora_paulinae | x | x | |
| LN884304_Acaulospora_papillosa | x | x | x |
| LN884303_Acaulospora_papillosa | x | x | x |
| LN884302_Acaulospora_papillosa | x | x | x |
| FR846382_Acaulospora_punctata | x | x | x |
| FR846384_Acaulospora_punctata | x | x | x |
| FR846385_Acaulospora_punctata | x | x | x |
| HF567941_Acaulospora_pustulata | x | x | x |
| HF567939_Acaulospora_pustulata | x | x | x |
| HF567938_Acaulospora_pustulata | x | x | x |
| KM057064_Acaulospora_reducta | x | x | |
| KM057066_Acaulospora_reducta | x | x | |
| KM057063_Acaulospora_reducta | x | x | |
| LN881566_Acaulospora_rugosa | x | x | x |
| LN881565_Acaulospora_rugosa | x | x | x |
| LN881564_Acaulospora_rugosa | x | x | x |
| KY362428_Acaulospora_saccata | x | x | x |
| KY362430_Acaulospora_saccata | x | x | x |
| KY362429_Acaulospora_saccata | x | x | x |
| FR692352_Acaulospora_scrobiculata | x | x | x |
| FR692354_Acaulospora_scrobiculata | x | x | x |
| FR692350_Acaulospora_scrobiculata | x | x | x |
| AM076384_Acaulospora_sieverdingii | x | x | |
| AM076382_Acaulospora_sieverdingii | x | x | |
| FR750153_Acaulospora_spinosa | x | x | x |
| FR750156_Acaulospora_spinosa | x | x | x |
| FR750152_Acaulospora_spinosa | x | x | x |
| HG422734_Acaulospora_spinosissima | x | x | x |
| HG422733_Acaulospora_spinosissima | x | x | x |
| HG422732_Acaulospora_spinosissima | x | x | x |
| KY413817_Acaulospora_spinulifera | x | ||
| KY413815_Acaulospora_spinulifera | x | ||
| KY413814_Acaulospora_spinulifera | x | ||
| HF567933_Acaulospora_tortuosa | x | x | x |
| HF567937_Acaulospora_tortuosa | x | x | x |
| HF567936_Acaulospora_tortuosa | x | x | x |
| MH045497_Acaulospora_tsugae | x | x | |
| MH045498_Acaulospora_tsugae | x | x | |
| MH333280_Acaulospora_tsugae | x | x | |
| AF378440_Acaulospora_tuberculata | x | ||
| FJ461799_Acaulospora_tuberculata | x | x | x |
| MT832207_Acaulospora_tuberculata | x | x | x |
| HG421736_Acaulospora_viridis | x | x | x |
| HG421738_Acaulospora_viridis | x | x | x |
| HG421737_Acaulospora_viridis | x | x | x |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, K.J.G.; Fernandes, J.A.L.; Magurno, F.; Leandro, L.B.A.; Goto, B.T.; Theodoro, R.C. Phylogenetic Review of Acaulospora (Diversisporales, Glomeromycota) and the Homoplasic Nature of Its Ornamentations. J. Fungi 2022, 8, 892. https://doi.org/10.3390/jof8090892
da Silva KJG, Fernandes JAL, Magurno F, Leandro LBA, Goto BT, Theodoro RC. Phylogenetic Review of Acaulospora (Diversisporales, Glomeromycota) and the Homoplasic Nature of Its Ornamentations. Journal of Fungi. 2022; 8(9):892. https://doi.org/10.3390/jof8090892
Chicago/Turabian Styleda Silva, Kássia J. G., José A. L. Fernandes, Franco Magurno, Larissa B. A. Leandro, Bruno T. Goto, and Raquel C. Theodoro. 2022. "Phylogenetic Review of Acaulospora (Diversisporales, Glomeromycota) and the Homoplasic Nature of Its Ornamentations" Journal of Fungi 8, no. 9: 892. https://doi.org/10.3390/jof8090892