
Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review


1
Department of Plant and Animal Production, Vocational School of Kızıltepe, Mardin Artuklu University, Mardin 47000, Turkey
2
Department of Plant Protection, Faculty of Agriculture, Bolu Abant Izzet Baysal University, Bolu 14030, Turkey
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(11), 1048; https://doi.org/10.3390/jof9111048
Submission received: 6 September 2023
/
Revised: 16 October 2023
/
Accepted: 23 October 2023
/
Published: 26 October 2023
(This article belongs to the Special Issue Plant Fungal Pathogens: Isolation, Characterization and Control Strategies, 2nd Edition)
Abstract
:Neoscytalidium dimidiatum, a plant- and human-associated fungus, has emerged as a substantial global ecological and agricultural threat aggravated by global warming. It inflicts various diseases, including canker, blight, dieback, leaf spot, root rot, and fruit rot, across a wide spectrum of fruit trees, field crops, shrubs, and arboreal species, with a host range spanning 46 plant families, 84 genera, and 126 species, primarily affecting eudicot angiosperms. Six genera are asymptomatic hosts. Neoscytalidium dimidiatum exhibits worldwide distribution, with the highest prevalence observed in Asia and North America, notably in Iran, Turkey, and California. Rising disease prevalence and severity, aggravated by climate change, particularly impact tropical arid places across 37 countries spanning all 7 continents. This comprehensive review encapsulates recent advancements in the understanding of N. dimidiatum, encompassing alterations in its taxonomic classification, host range, symptoms, geographic distribution, epidemiology, virulence, and strategies for effective management. This study also concentrates on comprehending the taxonomic relationships and intraspecific variations within N. dimidiatum, with a particular emphasis on N. oculus and N. hylocereum, proposing to consider these two species as synonymous with N. dimidiatum. Furthermore, this review identifies prospective research directions aimed at augmenting our fundamental understanding of host—N. dimidiatum interaction.
1. Introduction
In the face of escalating environmental changes and global climate fluctuations, the emergence of specific plant pathogens and the diseases they induce have become a pressing concern. Within this context, the monospecific plant pathogenic genus Neoscytalidium, represented by N. dimidiatum, has garnered substantial attention due to its remarkable adaptability and aggressive nature [1,2]. This pathogen poses a formidable threat as it gives rise to epidemics affecting a diverse array of plant species, manifesting as canker and dieback diseases that impact economically, industrially, forestally, and ornamentally important trees and shrubs. This adaptability, coupled with its aggressive tendencies, renders it a significant menace to the global agriculture and horticulture sectors. While Neoscytalidium is predominantly recognized as a phytopathogen, it also engenders a range of clinical conditions in humans. It affects individuals with underlying predispositions as well as those seemingly devoid of health concerns [3,4].
The pathogen enters host plant tissues through pre-existing wounds, giving rise to a spectrum of symptoms. Notably, host plants undergoing abiotic stress tend to exhibit intensified disease manifestations post-infection. In the realm of perennial plants like dragon fruits [5,6,7,8,9,10,11,12,13,14,15,16,17,18], citruses [19,20], grapevines [21,22,23,24,25,26], pines [27,28], stone fruits [29,30,31,32,33], Ficus spp. [34,35,36,37,38,39,40,41], pistachios [42], and willows [43,44], the symptomatic expression is particularly conspicuous, leading to yield reduction and shortened lifespans. In the context of dragon fruit canker, a devastating affliction affecting dragon fruit plants worldwide, the initiation of fungal infection ensues with the development of appressoria on the surface, followed by direct penetration into epidermal cells [45]. The transmission mechanisms of N. dimidiatum encompass seeds, propagation materials, soil, and airborne dispersal, with a marked tendency to persist within soil alongside infected debris [39,45,46,47,48,49].
Characterized by Scytalidium-like arthric chains of dark conidia and Fusicoccum-like conidia in conidiomata, N. dimidiatum is distinct [2,50,51,52,53]. Since the presence of ascospore-producing ascoma has not been documented, both coelomycetous and hyphomycetous morph conidia from diseased plant parts may serve as the primary source of Neoscytalidium-caused diseases [39,45,49]. However, the epidemiology, including the seasonal dynamics of various spore types, remains insufficiently elucidated. The taxonomic journey of N. dimidiatum has been marked by significant discoveries and revisions. Initially recognized as distinct species, such as N. novaehollandiae and N. orchidacearum, recent research endeavors have led to their consolidation under the species of N. dimidiatum [2]. However, the taxonomic landscape continues to evolve, with the emergence of novel fungal species such as N. oculus [54] and N. hylocereum [55], underscoring the intricate genetic diversity within this fungal group.
The adaptability of thermotolerant Neoscytalidium to diverse environmental conditions, including elevated temperatures and drought periods, likely contributes to its heightened virulence and extensive global distribution. To provide a comprehensive understanding of N. dimidiatum, this review amalgamates existing research while delving into novel insights across its attributes, epidemiology, virulence, and prospective management strategies. By enriching our comprehension of N. dimidiatum’s significance within the Botryosphaeriales order, this review augments our knowledge of this pathogen and its multifaceted impacts.
2. Taxonomy and Classification of Neoscytalidium dimidiatum
2.1. Historical Changes in Neoscytalidium dimidiatum Taxonomy
The taxonomic classification of N. dimidiatum has a turbulent history due to the fact that it generates two distinct asexual states known as synanamorphs, which have been described by multiple authors. The coelomycetous morph produces pycnidia with conidia that resemble Fusicoccum-like conidia produced in pycnidia, while the hyphomycetous morph produces arthric chains of conidia in the aerial mycelium, giving it a powdery appearance that resembles Scytalidium-like conidia [56,57,58]. Derived from the arthric synanamorph, the initial description of N. dimidiatum occurred when Penzig named it Torula dimidiata Penz. in 1882 [59]. Subsequently, another synanamorph of this species, categorized as a coelomycete, was identified by Nattrass [56] in 1933 and designated as Hendersonula toruloidea. This recognition came approximately fifty years after the initial classification by Penzig and was observed in pome and stone fruit trees. Wilson [60] proposed the name Exosporina fawcetti as the causal agent of a sudden wilt affecting Persian walnut trees (Juglans regia) in California. This assignment was grounded in the resemblance to a comparable incident reported by Fawcett [RAM., xv, p. 574] in 1923, wherein the incident took place within the same county and involved grapefruit and orange trees displaying similar symptomatic manifestations. Campbell and Mulder [61] introduced the new species Scytalidium hyalinum as the causal agent for the same clinical lesions previously associated with H. toruloidea. In the research conducted by Sutton and Dyko [57], a taxonomic reorganization was undertaken. They reclassified Dothiorella mangiferae, Fusicoccum eucalypti, Hendersonula agathidis, Hendersonula cypria, and Hendersonula toruloidea into Nattrassia mangiferae. Additionally, they introduced the name ‘Scytalidium dimidiatum’ to designate the corresponding mycelial synanamorph, drawing from the nomenclature of Torula dimidiata. This study also encompassed Torula dimidiata, Exosporina fawcettii, and Scytalidium lignicola, which were incorporated into the synonymy of Scytalidium dimidiatum based on shared mycelial synanamorphic characteristics. Subsequently, Farr et al. [58] conducted phylogenetic analysis using internal transcribed spacer region (ITS) and beta-tubulin gene (tub2) sequences, which led to the determination that Nattrassia mangiferae and Scytalidium dimidiatum should be reclassified under Fusicoccum, resulting in the name Fusicoccum dimidiatum. Additionally, they assigned the specific isolates of N. mangiferae responsible for inducing cankers on Pacific madrone trees as Fusicoccum arbuti. Through DNA sequencing, a definitive differentiation has been established between genuine N. mangiferae isolates and F. arbuti. However, Slippers et al. [62] conducted a study on isolates identified as Dothiorella mangiferae (Nattrassia mangiferae) obtained from mango trees in Australia and determined that they actually belonged to Fusicoccum. Consequently, they introduced the name Fusicoccum mangiferae (now classified as Neofusicoccum mangiferae) for this group of isolates. They also did not observe the Scytalidium-like synanamorph, which is consistent with the observations made by Sutton and Dyko [57] and Sydow et al. [63]. Therefore, the synonymy of H. toruloidea (which possesses a Scytalidium-like synanamorph) with F. mangiferae (which does not exhibit a Scytalidium-like synanamorph) was rejected. In a taxonomic revision of the Botryosphaeriaceae, Crous et al. [50] concluded that Scytalidium is polyphyletic and proposed the genus Neoscytalidium, accommodating S. dimidiatum as N. dimidiatum. Their study additionally unveiled the distinct classification of S. dimidiatum and Scytalidium’s type species, Scytalidium lignicola, as they belong to separate classes. Pavlic et al. [51] identified a fungal species as N. novaehollandiae, which was collected in Western Australia in July 2006, particularly from Crotalaria medicaginea. There have been suggestions that S. dimidiatum and S. hyalinum may be conspecific, proposing the name N. dimidiatum var. hyalinum [64]. In their study, Phillips et al. [52] classified the species as N. hyalinum and employed a multi-locus tree encompassing ITS, translation elongation factor 1-alpha gene (tef1), tub2, large subunit (of ribosomal RNA) gene (LSU), and small subunit (of ribosomal RNA) gene (SSU) sequences for its analysis within the Botryosphaeriaceae family. Huang et al. [53] identified a coelomycete in Thailand resembling asexual morphs within Botryosphaeriaceae. Through morphological and phylogenetic analysis, the strain was revealed as a new species, Neoscytalidium orchidacearum. Considering the phylogenetic similarity between S. hyalinum and N. dimidiatum and the fact that S. hyalinum is the older name, they proposed transferring S. hyalinum to the new genus Neoscytalidium and synonymizing N. dimidiatum [53]. Within the Neoscytalidium genus, Zhang et al. [2] conducted a comprehensive analysis by sequencing ITS, tef1, and tub2 genes across various Neoscytalidium spp. As a result, all examined species were determined to belong to N. dimidiatum (Penz.) Crous & Slippers. Additionally, N. novaehollandiae and N. orchidacearum were found to be synonymous with N. dimidiatum. To support this taxonomic assignment, the nucleotide similarities between the ex-type culture of N. dimidiatum and the sequences derived from the ex-type cultures of N. novaehollandiae and N. orchidacearum were compared. Specifically, the ITS gene showed 489 out of 492 matching positions (99.39%) with N. novaehollandiae and 486 out of 492 matching positions (98.78%) with N. orchidacearum. Regarding the tef1 gene, a similarity of 185 out of 187 positions (98.93%) was observed with N. novaehollandiae, while no tef1 sequence was available for N. orchidacearum. Moreover, the tub2 gene exhibited complete identity, with 100% sequence similarity at all 358 positions between N. dimidiatum and N. novaehollandiae, while no tub2 sequence was available for N. orchidacearum. Zhang et al. [2] also highlight the significant utility of the tef1 gene, which encodes translation elongation factor 1-alpha, in effectively distinguishing between various Neoscytalidium species.
2.2. Significance of Resolving Neoscytalidium Species Concepts
Resolving Neoscytalidium species concepts is crucial for addressing essential concerns related to polyphagous pathogen identification and control. It facilitates the implementation of control measures like crop rotation and mixed plantations; enhances our understanding of disease dynamics, allows for resistance monitoring; and supports epidemiological studies and research in fungal biology, ecology, and genetics. Ultimately, taxonomic clarity empowers comprehensive disease management and deepens our insights into disease dynamics.
The designation of a specific population or taxonomic group as a new fungal species is a central topic in taxonomic discussions [65]. Currently, the scientific community recognizes more than 30 distinct species concepts, although only a subset of these concepts is widely used in fungal taxonomy [66]. Each concept has its own advantages and disadvantages, and there is no universally agreed-upon standard. The process of taxonomic consolidation observed here mirrors the intricate historical complexities surrounding the classification of Neoscytalidium. This history has been marked by taxonomic disagreements and shifts in species boundaries, leading to challenges in understanding the species’ unique biology.
The use of both morphological and DNA-based methodologies has sometimes resulted in the excessive subdivision of N. dimidiatum into multiple taxa [50,51,52,53,56,57,58,59,60,61,64]. Conversely, other methodologies have grouped various taxa into a single species category [2,50,57,58,62]. Addressing these challenges requires the application of multiple genetic markers and coalescence-based methodologies, offering the potential for more precise and reliable insights into the complex taxonomic relationships within the Neoscytalidium genus.
In conclusion, the genetic investigation conducted by Zhang et al. [2] holds significant relevance in elucidating the taxonomic intricacies within the Neoscytalidium genus. Their integration of multiple gene loci brings clarity to the consolidation of species, with an emphasis on the role of tef1 in distinguishing Neoscytalidium species. Collectively, these studies provide invaluable insights into the taxonomy and delineation of species within Neoscytalidium. The taxonomic classification within the Neoscytalidium genus remains an ongoing area of investigation, and a more comprehensive understanding necessitates further sequencing of additional genetic markers for representative strains of different species. Achieving a conclusive taxonomic resolution for this genus requires continued research efforts.
2.3. Molecular Evidence for Synonymy and Intraspecific Variation in Neoscytalidium dimidiatum
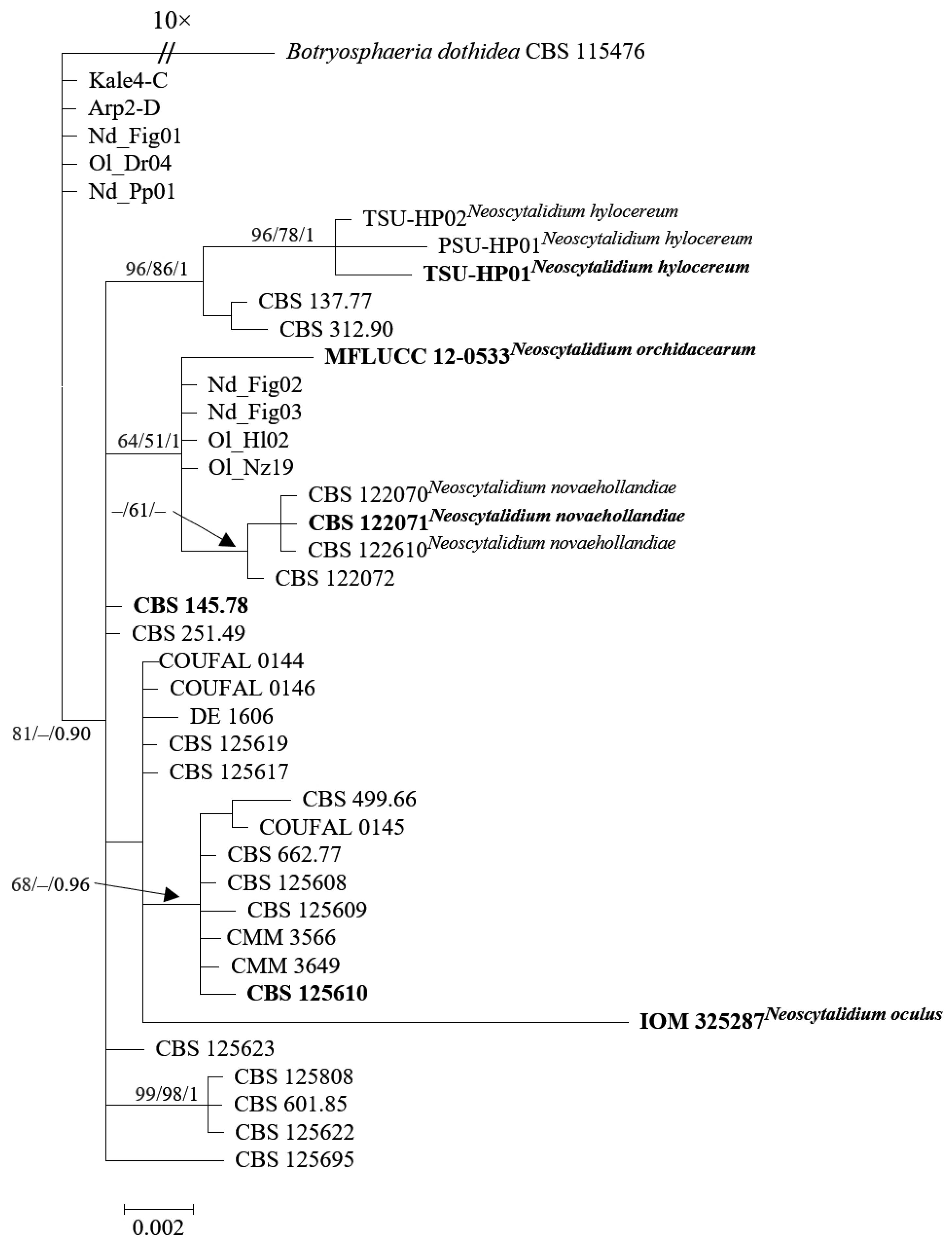
Precise taxonomic classification hinges upon a thorough comprehension of the genetic inter-relationships within closely related fungal species. Evidently, previously acknowledged taxonomic entities, such as N. novaehollandiae and N. orchidacearum, have undergone consolidation into N. dimidiatum, as elucidated by Zhang et al. [2]. Nevertheless, the taxonomic reassignment of N. oculus and N. hylocereum to N. dimidiatum remains pending and necessitates an in-depth inquiry for resolution.
In this context, it is pertinent to mention the work of Calvillo-Medina et al. [54], who introduced N. oculus, a novel fungal species linked to human keratitis in Latin America. Their taxonomic assignment, however, did not incorporate the tef1 and tub2 genes. Instead, they relied on a phylogenetic analysis of concatenated ITS and LSU ribosomal DNA sequences, along with morphological observations, to establish this new species. Another notable instance is the work of Wonglom et al. [55], where a distinct fungal pathogen, N. hylocereum, was identified. This discovery was achieved through a combined approach involving morphological assessments and molecular analyses. The DNA sequences derived from the ITS, tef1, and tub2 loci distinctly positioned these newly discovered isolates in a separate clade.
This part of the review aims to elucidate the taxonomic relationships and intraspecific variations within Neoscytalidium species, with a particular focus on N. oculus and N. hylocereum. These two species exhibit morphological similarities while posing challenges for precise species differentiation.
To address this challenge, we employed molecular analyses to clarify their taxonomic designations (Figure 1). The analysis of ITS, tef1, and tub2 loci (Table S1) was conducted using sequences obtained from the GenBank nucleotide database. Multiple sequence alignments were performed using the MAFFT v.7 online interface [67] and subsequently manually edited in MEGA X [68]. Phylogenetic trees were constructed using Maximum Likelihood (ML), Maximum Parsimony (MP), and Bayesian analysis (BA). ML analysis utilized RAxML-HPC BlackBox v. 8.2.10 [69], MP analysis was conducted with PAUP v. 4.0b10 [70], and Bayesian analysis employed MrBayes 3.2.7 [71]. The resulting trees were visualized using FigTree v. 1.4.2 (http://tree.bio.ed.ac.uk/software/figtree, accessed on 22 August 2023). Botryosphaeria dothidea (CBS 115476) served as an outgroup to root the tree.
The exploration of N. oculus and N. dimidiatum isolates began with a detailed investigation of their genetic makeup through LSU and ITS sequencing. Surprisingly, the LSU sequence demonstrated a remarkable genetic similarity of up to 100%, suggesting an intimate genetic connection. This finding, however, is juxtaposed by discernible variations in the ITS sequence. Particularly, disparities emerged within specific regions: the terminal 11 bases of the SSU, the first 24 bases of ITS1, and the terminal 43 bases of the ITS2 region. The discrepancies, encompassing 65 differing bases, also include the critical 22 bases crucial for binding to LSU rRNA. It is noteworthy that BLAST analysis identified self-overlapping occurrences exclusively within the GenBank database for these specific regions, implying potential inaccuracies in the sequence data input.
Further exploration focused on the tef1 sequences of N. hylocereum isolates, revealing a substantial similarity ranging from 99.61% to 100% when compared to tef1 sequences of N. dimidiatum. This genetic resemblance establishes a foundational criterion for their taxonomic classification. However, the diversity observed in ITS sequences, specifically within the terminal 13 bases of the SSU and the initial 24 bases of ITS1, suggests that relying solely on a single gene sequence might be insufficient for precise classification.
Delving deeper into the genetic makeup, analysis of the tub2 sequences revealed genetic disparities among the isolates, primarily rooted in variances within the initial 16 nucleotides of these sequences. The outcomes of BLAST analysis conducted on these sequences brought forth instances of self-matching solely within the NCBI GenBank nucleotide database, further supporting the notion of potential errors in sequence inputs.
In a collective summary, the comprehensive molecular analyses presented in this study provide strong evidence to support the proposition of synonymy between N. oculus and N. hylocereum with N. dimidiatum. While genetic similarities in specific regions are apparent, the highlighted differences within key gene regions underscore the intricate nature of their genetic relationships. The revelation of a unique ITS clade within the Turkish isolates, notably distant from reference isolates, highlights the critical need for precise taxonomic classification, especially in light of differences across multiple gene regions.
In conclusion, the comprehensive assessments of LSU, ITS, tef1, and tub2 sequences offer strong evidence supporting the classification of N. oculus and N. hylocereum as synonyms of N. dimidiatum. The observed genetic resemblances are accompanied by significant differences within crucial gene regions. The case of N. oculus further emphasizes the necessity of employing diverse gene regions for accurate species identification, shedding light on the intricate taxonomic complexity in the fungal realm.
Moreover, the inclusion of previously documented Turkish isolates from diverse hosts enhances the depth of our phylogenetic exploration concerning N. dimidiatum isolates, revealing a distinctive ITS clade embedded within the species. Through their application of molecular phylogenetic methodologies involving ITS, tef1, and tub2 sequence alignments, Zhang et al. [2] marked off the examined Turkish isolates, specifically Kale4-C and Arp2-D, as a distinct and recognizable group separate from the broader population of Neoscytalidium isolates, designating this particular cluster as Neoscytalidium sp.1. Even though Güney et al. [49]’s results agreed with the observation of this phylogenetic divergence among isolates from different chickpea cultivation sites, their subsequent statistical analysis did not show any significant differences in the size of spores produced both as arthroconidia and within conidiomata among these isolates that were in a separate clade. The clear separation of specific Turkish isolates into a separate cluster, evident when comparing them to the larger collection of recognized Neoscytalidium isolates in phylogenetic analyses relying on multi-locus sequences, strengthens the understanding that this divergence is best explained as a characteristic of intraspecies polymorphism. This interpretation holds more weight than considering it as sufficient evidence to classify these isolates as distinct taxonomic entities.
2.4. Current Species Name
2.5. Classification
Kingdom Fungi; Phylum Ascomycota; Subphylum Pezizomycotina; Class Dothideomycetes; Subclass Incertae sedis; Order Botryosphaeriales; Family Botryosphaeriaceae; Genus Neoscytalidium.
2.6. Synonyms and Basionyms
- 1.
- Torula dimidiata Penz. [59].
- 2.
- Hendersonula toruloidea Nattrass [56].
- 3.
- Exosporina fawcettii E.E. Wilson [60].
- 4.
- Scytalidium hyalinum C.K. Campb. & J.L. Mulder [61].
- 5.
- Scytalidium dimidiatum (Penz.) B. Sutton & Dyko [57].
- 6.
- Fusicoccum dimidiatum (Penz.) D.F. Farr [58].
- 7.
- Neoscytalidium dimidiatum (Penz.) Crous & Slippers [50].
- 8.
- Neoscytalidium novaehollandiae Pavlic, T.I. Burgess & M.J. Wingf. [51].
- 9.
- Neoscytalidium dimidiatum var. hyalinum (C.K. Camp. & J.L. Mulder) Madrid, Cano, Stchigel & Guarro [64].
- 10.
- Neoscytalidium hyalinum (C.K. Campb. & J.L. Mulder) A.J.L. Phillips, Groenewald & Crous [52].
- 11.
- Neoscytalidium orchidacearum S. K. Huang, N. Tangthirasunun, J. C. Kang & K. D. Hyde [53].
- 12.
- Neoscytalidium oculus (Neoscytalidium oculi in MycoBank) J. Mena, Raymundo & Bautista-de-Lucio (the taxonomic status of this species was consolidated as a synonym in the present investigation).
- 13.
- Neoscytalidium hylocereum Kheawleng, Intaraa-nun & Rodkaew (the taxonomic status of this species was consolidated as a synonym in the present investigation).
2.7. Rejected Synonyms
- 1.
- Dothiorella mangiferae Syd. & P. Syd. (1916) [57].
- 2.
- Fusicoccum eucalypti Sousa da Câmara (1929) [57].
- 3.
- Hendersonula cypria Nattrass (1937) [57].
- 4.
- Hendersonula agathidis H.E. Young (1948) [57].
- 5.
- Nattrassia mangiferae (Syd. & P. Syd.) B. Sutton & Dyko [57].
- 6.
- Fusicoccum mangiferae (Syd. & P. Syd.) Johnson, Slippers & M.J. Wingf. [62].
- 7.
- Scytalidium lignicola Pesante [50].
Note: The first six species listed above are currently classified as Neofusicoccum mangiferae (Syd. & P. Syd.) Crous, Slippers & A.J.L. Phillips [50].
3. Identification
The isolation and cultivation of Neoscytalidium dimidiatum are accomplished using conventional methods on diverse media such as Potato Dextrose Agar (PDA), Malt Extract Agar (MEA), and Oatmeal Agar (OA). Initially, colonies appear colorless but undergo a transformation within seven days, transitioning from a pale brown or greenish olivaceous shade to citrine hues at the center (Figure 2a,b). Over time, the colonies become black both on the surface and beneath, accompanied by moderately fluffy mycelium that is suppressed and has smooth edges [2,50,51,52].
Neoscytalidium dimidiatum, with an unknown teleomorph, presents two distinct asexual forms known as synanamorphs. The hyphomycetous morphology generates powdery to touch-textured conidia in arthric chains through hyphal fragmentation (Figure 2c). These conidia exhibit diverse shapes—from cylindrical-truncate to oblong-obtuse and doliiform—resembling those seen in the Scytalidium genus. They measure 4–16.5 × 2.5–8.5 μm and have thick walls that shift from an initial hyaline state to a dark brown coloration with age [2,50,51,52,53].
The coelomycetous morph generates hyaline, ellipsoidal conidia, which may have zero to two septa and possess a darkened central cell, closely resembling those found in the Fusicoccum genus. These conidia measure approximately 10–16 × 3.5–6.5 μm and are enclosed within solitary or multilocular conidiomata, often referred to as pycnidia. These pycnidia can be observed on various substrates, including sterilized pine needles and corn straw, using water agar (Figure 2d,e). The conidiogenous cells responsible for conidia production are holoblastic, cylindrical, and hyaline, with dimensions of 6–14 × 1.5–4 μm (Figure 2f).
For identification purposes, specific gene regions such as ITS, tef1, and tub2 are employed, with tef1 playing a pivotal role in the accurate identification of Neoscytalidium [2,50,51,52,53]. Additionally, the optimum mycelial growth of this fungus occurs at temperatures ranging from 33 to 35 °C, with conidial germination reaching its peak between 38 and 40 °C [11,30,49,72,73,74,75,76].
4. Host Range, Symptoms, and Geographic Distribution of Neoscytalidium dimidiatum
4.1. Host Range and Geographical Distribution Diversity of Neoscytalidium dimidiatum
Neoscytalidium dimidiatum demonstrates a wide-ranging infective capacity, encompassing a diverse array of plant species, as substantiated by records that document its presence in 126 distinct host species. Yet, the reliability of these reports remains a challenge due to taxonomic intricacies linked with N. dimidiatum and its historically rejected synonyms. Throughout history, various checklists have been published as outcomes of extensive surveys of plant pathogens. In recent times, many of these inventories have been amalgamated into comprehensive databases, such as the United States Department of Agriculture (USDA) database. These databases, including the USDA fungal database and similar sources from other countries, stand as invaluable repositories for comprehending the host spectrum and geographic distribution of plant pathogenic fungi. Despite the temporary inactivity of the USDA fungal database, we utilized the list of sources saved in 2018 to illustrate the geographic distribution of N. dimidiatum and its basionyms and synonyms. To construct an exhaustive dataset, we systematically compiled plant host data from diverse literature sources and the USDA database. This compilation, consisting of 250 instances of N. dimidiatum and its synonymous taxa from natural habitats, serves as a foundation for analysis (Table 1). This table, meticulously organized, presents a comprehensive listing of hosts associated with N. dimidiatum. Host species, along with their common names and corresponding families, are alphabetically arranged. Additionally, the table showcases the basionym names of N. dimidiatum reported in each country, illustrating its global distribution. This well-structured table offers a valuable and up-to-date resource for plant pathologists and researchers delving into the intricate relationship between N. dimidiatum and its diverse host range across various countries. Leveraging this host information has enabled the validation of N. dimidiatum’s expansive host range. Presently, the confirmed host count stands at over 100 (126 to date), cementing the understanding of its diverse host interactions. These hosts span 46 distinct families, with 43 of them belonging to the category of seed plants. The list of host families, including Acanthaceae, Anacardiaceae, Apocynaceae, Araliaceae, Berberidaceae, Betulaceae, Boraginaceae, Cactaceae, Casuarinaceae, Combretaceae, Convolvulaceae, Cucurbitaceae, Ebenaceae, Ericaceae, Euphorbiaceae, Fabaceae, Fagaceae, Juglandaceae, Lamiaceae, Lythraceae, Malvaceae, Meliaceae, Moraceae, Myrtaceae, Oleaceae, Proteaceae, Rhamnaceae, Rhizophoraceae, Rosaceae, Rutaceae, Salicaceae, Solanaceae, Ulmaceae, and Vitaceae, collectively fall within the classification of eudicots. These families are categorized under angiosperms, also known as flowering plants. Monocot hosts, represented by the Amaryllidaceae, Araceae, Asparagaceae, Asphodelaceae, Bromeliaceae, Dioscoreaceae, Iridaceae, Musaceae, and Orchidaceae families, constitute a significant subdivision within angiosperms. In particular, within the spectrum of seed plants, the majority of host families are classified as eudicots, making monocot hosts relatively infrequent for N. dimidiatum. The presence of N. dimidiatum within Araucariaceae, Cupressaceae, and Pinaceae aligns with the gymnosperms classification. The full host range of this species is not known as, apparently, it can be an endophyte/saprophyte instead of a pathogen in some plant species. Reports lacking the integration of morphological analysis and multigene phylogenetic analysis for precise pathogen identification introduce uncertainty into the existing knowledge. Thus, N. dimidiatum showcases its ability to infect a broad spectrum of seed plant families and potentially extends to other plant groups, including gymnosperms, establishing itself as one of the most versatile fungal plant pathogens in terms of host range.
The geographic distribution of N. dimidiatum underscores its global presence, with varying prevalence. The highest number of recorded occurrences, totaling 132, is notably concentrated in Asia, indicating a significant hotspot for this pathogen. North America reports 50 occurrences, suggesting a substantial yet comparatively lower frequency. Africa documents 26 instances, demonstrating the pathogen’s notable presence on the continent. In South America and Australia, 16 and 15 recorded occurrences, respectively, indicate a balanced distribution across these regions. Europe, with 11 reported cases, points to a noteworthy but comparatively lower occurrence rate. In contrast, Oceania reports only a single recorded instance, signifying a rare presence in this region (for all relevant references throughout this chapter, see Table 1).

{kind=link}
{kind=link}
Table 1.
Comprehensive compilation of hosts associated with Neoscytalidium dimidiatum. The host species, along with their scientific nomenclature and familial categorization, are systematically organized in alphabetical order. Concurrently, the basionym designations of N. dimidiatum, as documented within each country, are presented, showcasing its worldwide dispersion.
Table 1.
Comprehensive compilation of hosts associated with Neoscytalidium dimidiatum. The host species, along with their scientific nomenclature and familial categorization, are systematically organized in alphabetical order. Concurrently, the basionym designations of N. dimidiatum, as documented within each country, are presented, showcasing its worldwide dispersion.
| Plant Species Family | Host Species | Common Host Name | Identified Species Name | Target Loci for IDENTIFICATION | Koch’s Postulates | Symptoms | Country | Continent | References |
|---|---|---|---|---|---|---|---|---|---|
| Acanthaceae | Avicennia marina | White mangrove | Nd | ITS | + | Canker and dieback | Iran | Asia | [77] |
| Amaryllidaceae | Hymenocallis littoralis | White spider lily | Nd | ITS | + | Leaf blight | Malaysia | Asia | [78] |
| Clivia miniata | Natal lily | Nd | ITS, tef1 | + | Leaf blight | Iran | Asia | [79] | |
| Anacardiaceae | Anacardium occidentale | Cashew | Nh | ITS, tef1 | + | Dieback and stem and branch cankers | Brazil | South America | [72] |
| Mangifera indica | Mango | Ht | − | + | Leaf spot | India | Asia | [80] | |
| Ht | − | + | Dieback | Niger | Africa | [81] | |||
| Ef | − | NA | Herbarium specimen records | South Africa | Africa | [57] | |||
| Ht | − | NA | NA | Brazil | South America | [82] | |||
| Fd | ITS, tub2 | − | NA | US—California | North America | [58] | |||
| Nd and Nn | ITS, tef1 | + | Dieback | Australia | Australia | [35] | |||
| Nd and Nn | ITS, tef1 | + | Canker | Australia | Australia | [83] | |||
| Nd | ITS, tef1 | + | Dieback and stem-end rot | Brazil | South America | [84] | |||
| Nh | ITS, tef1 | + | Dieback and stem and branch cankers | Brazil | South America | [72] | |||
| Pistacia vera | Pistachio | Nd | ITS, tef1, tub2 | + | Endophytic | Iran | Asia | [85] | |
| Nd | ITS, LSU | + | Canker, shoot blight, and root rot | Turkey | Asia | [42] | |||
| Nn | ITS, tef1 | + | Dieback | Turkey | Asia | [86] | |||
| Rhus typhina | Staghorn sumac | Td | ITS, tub2 | NA | NA | US—West Virginia | North America | [87,88] | |
| Apocynaceae | Nerium oleander | Oleander | Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] |
| Araceae | Thaumatophyllum bipinnatifidum (Philodendron bipinnatifidum) | Split-leaf philodendron | Ht | − | NA | NA | India | Asia | [89] |
| Araliaceae | Meryta denhamii | Meryta | Nd | ITS, tef1, tub2 | + | Branch canker and dieback | Italy | Europe | [90] |
| Araucariaceae | Agathis robusta (A.palmerstoni) | Kauri | Ef | − | NA | Herbarium specimen records | Australia | Australia | [57] |
| Araucaria sp. | Chilean pine | Ht | − | NA | NA | Malaysia | Asia | [91] | |
| Asparagaceae | Agave sp. | Century plant | Ht | − | NA | NA | Guinea | Africa | [92] |
| Agave americana | Century plant | Ef | − | NA | Herbarium specimen records | India | Asia | [57] | |
| Agave sisalana | Sisal | Nd | ITS, tef1, tub2 | + | Black leaf spot | China | Asia | [93] | |
| Furcraea foetida (F. gigantea) | Green aloe | Ht | − | NA | NA | Malaysia | Asia | [94] | |
| Sansevieria hyacinthoides (S. guineensis) | African Bowstring Hemp | Ht | − | NA | NA | Guinea | Africa | [92] | |
| Sansevieria trifasciata (Dracaena trifasciata) | Dracaena | Nd | ITS | + | Leaf blight | Malaysia | Asia | [95] | |
| Nd | ITS, tef1 | + | Leaf blight | Brazil | South America | [96] | |||
| Asphodeloideae | Aloidendron dichotomum | Quiver tree | Nd | ITS, LSU, tef1, tub2, chs−1 | − | Epiphyte on stems | South Africa | Africa | [97] |
| Berberidaceae | Berberis vulgaris | Barberry | Nd | ITS, tef1, tub2, act | + | Canker and dieback | Iran | Asia | [98] |
| Betulaceae | Alnus glutinosa | Common alder | Nn | ITS, tef1 | + | Branch and trunk cankers | Iran | Asia | [1] |
| Carpinus betulus | Common hornbeam | Nn | ITS, tef1 | + | Branch and trunk cankers | Iran | Asia | [1] | |
| Boraginaceae | Cordia myxa | Assyrian plum | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] |
| Bromeliaceae | Ananas comosus (A. sativa) | Pineapple | Ht | − | NA | Leaf spot; fruit rot | Malaysia | Asia | [94] |
| Ht | − | NA | Leaf spot; fruit rot | Tanzania | Africa | [99] | |||
| Ef | − | NA | Herbarium specimen records | Sierra Leone | Africa | [57] | |||
| Ef | − | NA | Herbarium specimen records | Solomon Islands—Rendova | Oceania | [57] | |||
| Nd | ITS, LSU | + | Postharvest stem end rot | Malaysia | Asia | [100] | |||
| Cactaceae | Nopalea cochenillifera | Pickly pear cactus | Nh | ITS, tef1, tub2 | + | Squamous cladode spots | Brazil | South America | [101] |
| Selenicereus (Hylocereus) undatus | Pitahaya (white-fleshed dragon fruit) | Nd | ITS | + | Stem canker | Taiwan | Asia | [5] | |
| Nd | ITS | + | Brown stem-spot-forming canker | China | Asia | [6] | |||
| Nd | ITS, tub2 | + | Internal black rot | Israel | Asia | [8] | |||
| Nd | ITS | + | Canker, internal brown rot | China | Asia | [7,102] | |||
| Nd | ITS | + | Stem and fruit canker | US—Florida | North America | [10] | |||
| Nd | ITS, tub2 | + | Stem and fruit canker | US—Florida | North America | [11] | |||
| Selenicereus (Hylocereus) undatus × S. polyrhizus | Red-fleshed dragon fruit | Nd | ITS, tub2 | + | Stem and fruit canker | US—Florida | North America | [11] | |
| Selenicereus (Hylocereus) polyrhizus | Red-fleshed dragon fruit | Nd | ITS | + | Stem canker | Taiwan | Asia | [5] | |
| Nd | ITS | + | Stem canker | Malaysia | Asia | [9] | |||
| Nd | ITS | + | Stem canker | China | Asia | [12] | |||
| Nd | ITS, LSU, tub2 | + | Stem canker | Thailand | Asia | [15] | |||
| Selenicereus (Hylocereus) megalanthus | Yellow pitahaya (dragon fruit) | Nd | ITS, tef1, tub2 | + | Stem canker | Malaysia | Asia | [17] | |
| Nd | ITS, tef1, tub2 | + | Stem canker | Ecuador | South America | [18] | |||
| Selenicereus (Hylocereus) monacanthus | Dragon fruit | Nd | tub2 | + | Stem and fruit canker | Philippines | Asia | [13] | |
| Selenicereus spp. (Hylocereus spp.) | Dragon fruit | Nd | ITS, tef1 | + | Stem canker | Puerto Rico | North America | [14] | |
| Nd | ITS, tef1, tub2 | + | Stem canker | India | Asia | [16] | |||
| Casuarinaceae | Casuarina sp. | Casuarina | Ht | − | NA | NA | Pakistan | Asia | [103] |
| Combretaceae | Conocarpus erectus | Buttonwood or button mangrove | Nn | ITS, LSU, tef1 | + | Lamination of the trunk bark | Iran | Asia | [40] |
| Convolvulaceae | Ipomoea batatas | Sweet potato | Ht | − | NA | NA | Malaysia | Asia | [94,104] |
| Nd | ITS, tef1 | + | Root rot | Brazil | South America | [105] | |||
| Nd | ITS, tef1 | + | Root and stem rot | Brazil | South America | [106] | |||
| Cucurbitaceae | Cucumis melo | Melon | Nh | ITS | + | Fruit rot | Iran | Asia | [107] |
| Cupressaceae | Cupressus sempervirens | Mediterranean cypress | Nn | ITS, LSU, tef1 | − | Canker and dieback | Iran | Asia | [40] |
| Sequoiadendron giganteum (Sequoia gigantea) | Giant redwood, giant sequoia | Ht | − | NA | NA | US—California | North America | [108] | |
| Dioscoreaceae | Dioscorea esculenta | Lesser yam | Nd | ITS, tef1, tub2 | + | Dieback | China | Asia | [109] |
| Dioscorea rotundata | Yam | Nd | ITS, tub2 | − | Tuber dry rot | Colombia | South America | [110] | |
| Ebenaceae | Diospyros kaki | Japanese persimmon | Nn | ITS, tef1 | + | Branch dieback | Turkey | Asia | [111] |
| Ericaceae | Arbutus menziesii | Madrone | Ht | − | NA | NA | US—Washington | North America | [112,113] |
| Ht | − | NA | NA | Canada | North America | [114] | |||
| Ht | − | NA | Canker | US—California | North America | [115] | |||
| Arbutus unedo | Strawberry tree | Ht | − | + | Leaf spotting and defoliation | Greece | Europe | [116] | |
| Euphorbiaceae | Jatropha curcas | Physic nut, a biofuel plant | Nh | ITS | + | Collar and root rot | Brazil | South America | [117] |
| Nd | ITS, tef1, tub2 | + | Collar and root rot | Brazil | South America | [118] | |||
| Manihot esculenta (M. utilissima) | Cassava | Ht | − | NA | NA | Ghana | Africa | [119] | |
| Ht | − | NA | NA | Kenya | Africa | [120] | |||
| Ef | − | NA | Herbarium specimen records | Ghana | Africa | [57] | |||
| Nh | ITS, tef1, tub2 | + | Black root rot | Brazil | South America | [121] | |||
| Nd | ITS, tef1, rpb2 | + | Black root rot and stem cutting dry rot | Brazil | South America | [122] | |||
| Nd | ITS, tef1 | + | Black stem and root rot | Thailand | Asia | [123] | |||
| Fabaceae | Acacia auriculiformis | Earleaf acacia | Ht | − | NA | NA | India | Asia | [89,124] |
| Ef | − | NA | Herbarium specimen records | India | Asia | [57] | |||
| Acacia melanoxylon | Australian blackwood | Ht | − | NA | NA | India | Asia | [124,125] | |
| Acacia synchronicia | Bardi bush | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] | |
| Nh | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Albizia lebbeck | Siris tree | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Bauhinia purpurea | Orchid tree | Nn | ITS, LSU, tef1 | + | Lamination of the trunk bark | Iran | Asia | [40] | |
| Cassia fistula | Golden shower tree | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] | |
| Cassia floribunda | Arsenic bush | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] | |
| Cicer arietinum | Chickpea | Nd | ITS, tef1, tub2 | + | Blight and root rot | Turkey | Asia | [49] | |
| Crotalaria medicaginea | Rattlepods | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] | |
| Nd | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Delonix regia | Royal poinciana | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Nd | ITS, LSU, tef1, tub2 | + | Stem canker | United Arab Emirates | Asia | [127] | |||
| Lysiphyllum cunninghamii | Kimberley bauhinia or jigal tree | Nn | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |
| Parkinsonia aculeata | Palo verde | Nn | − | − | Dieback (when used as a bipherbicide) | Australia | Australia | [128] | |
| Peltophorum petrocarpum | Copperpod | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Fagaceae | Castanea sativa | Sweet chestnut | Ef | − | NA | Canker | US—California | North America | [129] |
| Ht | − | NA | Canker | US—California | North America | [115] | |||
| Fagus orientalis | Oriental beech | Nn | ITS, tef1 | + | Branch and trunk cankers | Iran | Asia | [1] | |
| Quercus brantii | Persian oak | Nn | ITS, LSU, tef1 | + | Dieback | Iran | Asia | [74] | |
| Nd | ITS, LSU, SSU | + | Decline and sooty canker | Iran | Asia | [130] | |||
| Iridaceae | Gladiolus sp. | Gladiolus | Ht | − | NA | NA | US—California | North America | [115] |
| Juglandaceae | Juglans californica | California black walnut | Ht | − | NA | On twigs | US—California | North America | [131] |
| Juglans regia | English walnut | Ef | − | NA | Branch wilt | US—California | North America | [60,129,132] | |
| Ht | − | NA | NA | US—California | North America | [115,133] | |||
| Fd | ITS, tub2 | − | NA | US—California | North America | [58,134] | |||
| Nd | ITS, tef1, tub2 | + | Black canker and death of graft union | US—California | North America | [135,136] | |||
| Nd | ITS, LSU, tef1 | + | Black canker, root rot, decline | Turkey | Asia | [137,138] | |||
| Nd | Decline | Iran | Asia | [139] | |||||
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Lamiaceae | Lavandula angustifolia | Lavender | Nd | ITS, tef1 | + | Foliar and stem blight | Turkey | Asia | [140] |
| Melissa officinalis | Lemon balm | Nd | ITS, tef1 | + | Blight | Turkey | Asia | [141] | |
| Origanum onites | Turkish oregano | Nd | ITS, tef1 | + | Leaf blight | Turkey | Asia | [142] | |
| Salvia officinalis | Common sage | Nn | ITS, tef1 | + | Root rot and foliar blight | Turkey | Asia | [143] | |
| Lythraceae | Punica granatum | Pomegranate | Nd | ITS, tef1, tub2, act | + | Necrotic wood tissues | Iran | Asia | [144] |
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Malvaceae | Adansonia gibbosa | Baobab | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] |
| Adansonia gregorii | Boab | Nh | ITS, tef1 | + | Endophyte as a potential pathogen, canker | Australia | Australia | [126] | |
| Hibiscus rosa-sinensis | Chinese hibiscus | Nn | ITS, LSU, tef1 | − | Canker and dieback | Iran | Asia | [40] | |
| Thespesia populnea | Portia tree | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Meliaceae | Azadirachta indica | Neem | Nh | ITS, tef1, tub2, act | + | Decline | Iran | Asia | [145] |
| Melia azedarach | Chinaberry | Ht | − | − | NA | Pakistan | Asia | [146] | |
| Nd | ITS, LSU, tub2 | − | Canker and dieback | Iraq | Asia | [147] | |||
| Moraceae | Ficus benghalensis | Indian banyan | Sd | − | − | Dieback | Oman | Asia | [34] |
| Nd | Sooty canker | Iran | Asia | [39] | |||||
| Nn | ITS, LSU, tef1 | − | Dieback and sooty canker | Iran | Asia | [40] | |||
| Ficus benjamina | Weeping fig | Nd | ITS, LSU, rpb2 | Branch dieback | Mexico | North America | [36] | ||
| Nd | ITS, LSU, SSU | + | Sooty canker | Egypt | Africa | [37] | |||
| Ficus carica | Common fig | Ht | − | − | NA | Cyprus | Europe | [148] | |
| Ht | − | NA | NA | US—California | North America | [115,131,149] | |||
| Ht | − | NA | Herbarium specimen records | Cyprus | Europe | [57] | |||
| Sd | − | − | Dieback | Oman | Asia | [34] | |||
| Nd | ITS, tef1 | + | Dieback | Australia | Australia | [35] | |||
| Nd | ITS, tef1, tub2 | + | Dieback and canker | Turkey | Asia | [38] | |||
| Nd | − | Limb dieback | US—California | North America | [41] | ||||
| Ficus nitida | Chinese banyan | Nd | ITS, LSU, SSU | + | Sooty canker | Egypt | Africa | [37] | |
| Ficus religiosa | Bodhi tree | Nn | ITS, LSU, tef1 | + | Dieback and sooty canker | Iran | Asia | [40] | |
| Ficus retusa | Banyan tree | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Morus alba | White mulberry | Ef | − | NA | Herbarium specimen records | Pakistan | Asia | [57] | |
| Ef | − | NA | Herbarium specimen records | US | North America | [57] | |||
| Ht | − | − | NA | Pakistan | Asia | [146] | |||
| Nn | ITS, LSU, tef1 | + | Shoot and branch deaths | Turkey | Asia | [150] | |||
| Morus bombycis (Morus australis) | Korean mulberry | Nn | ITS, LSU, tef1 | + | Branch necrosis | Turkey | Asia | [150] | |
| Morus nigra | Black mulberry | Nn | ITS, LSU, tef1 | + | Shoot and branch deaths | Turkey | Asia | [150] | |
| Nn | ITS, LSU, tef1 | + | Sooty canker | Iran | Asia | [40] | |||
| Musaceae | Musa nana | Banana (dwarf) | Ht | − | − | Tip rot | Jamaica | North America | [151] |
| Musa spp. | Banana | Ht | − | − | Tip rot | Hawaii | North America | [88,152] | |
| Musa acuminata | Banana | Ht | − | − | Leaf spot | Hawaii | North America | [153] | |
| Myrtaceae | Callistemon viminalis | Weeping bottlebrush | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] |
| Eucalyptus camaldulensis | River red gum | Nd | ITS | + | Sooty canker | Iraq | Asia | [154] | |
| Nn | ITS, LSU, tef1 | + | Dieback, lamination of the trunk bark, and sooty canker | Iran | Asia | [40] | |||
| Eucalyptus sp. | Eucalyptus | Ef | − | NA | Herbarium specimen records | Portugal | Europe | [57] | |
| Nn | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Eucalyptus spp. | Eucalyptus | Nd | ITS, LSU, tub2 | − | Canker and dieback | Iraq | Asia | [147] | |
| Psidium guajava | Guava | Ht | − | NA | NA | India | Asia | [104] | |
| Nh | ITS, tef1 | + | Dieback and stem and branch cankers | Brazil | South America | [72] | |||
| Nd | ITS, tef1 | + | Postharvest fruit rot | Malaysia | Asia | [155] | |||
| Syzygium cumini | Java plum | Nh | ITS, tef1, tub2, act | + | Cankers and wedge-shaped wood necrosis | Iran | Asia | [156] | |
| Nh | ITS, tef1, tub2, act | + | Asymptomatic wood tissue | Iran | Asia | [156] | |||
| Nn | ITS, LSU, tef1 | + | Lamination of the trunk bark | Iran | Asia | [40] | |||
| Oleaceae | Olea europaea | Olive | Nd | ITS, tef1, tub2 | + | Canker and leaf scorch | Turkey | Asia | [76] |
| Orchidaceae | Arachnis sp. | Scorpion orchid | Ht | − | − | NA | Malaysia | Asia | [157] |
| NA | Orchid | No | ITS, LSU | − | From a fallen orchid leaf | Thailand | Asia | [53] | |
| Cattleya lueddemanniana var. lueddemanniana | Cattleya orchid | No | ITS, LSU | + | Leaf spot | Thailand | Asia | [158] | |
| Cattleya × hybrid | Orchids | Nd | ITS | + | Leaf blight | Taiwan | Asia | [159] | |
| Pinaceae | Picea pungens | Blue spruce | Nd | ITS, tef1, tub2 | + | Needle blight | Turkey | Asia | [160] |
| Pinus eldarica | Afghan pine | Nd | ITS, LSU | + | Shoot and needle blight | Turkey | Asia | [27] | |
| Nn | ITS, LSU, tef1 | + | Dieback | Iran | Asia | [28] | |||
| Pinus nigra | European black pine | Nd | ITS, LSU | + | Shoot and needle blight | Turkey | Asia | [27] | |
| Pinus sylvestris | Scots pine | Nd | ITS, LSU | + | Shoot and needle blight | Turkey | Asia | [27] | |
| Proteaceae | Grevillia agrifolia | Blue grevillea | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] |
| Nn | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Rhamnaceae | Ziziphus spina-christi | Christ’s thorn jujube | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] |
| Rhizophoraceae | Rhizophora mucronata | Red mangrove | Nd | ITS | + | Canker and dieback | Iran | Asia | [77] |
| Rosaceae | Malus domestica (M. pumila) | Apple | Ht | − | NA | Gummosis and dieback | Egypt | Africa | [56] |
| Ht | − | NA | NA | India | Asia | [89] | |||
| Ef | − | NA | Herbarium specimen records | India | Asia | [57] | |||
| Ef | − | NA | Herbarium specimen records | Iraq | Asia | [57] | |||
| Nd | Branch canker | Iran | Asia | [161] | |||||
| Nd | ITS, tef1 | + | Branch dieback and canker | Turkey | Asia | [32] | |||
| Nd | Cankers | China | Asia | [162] | |||||
| Prunus armeniaca | Apricot | Ht | − | NA | Gummosis and dieback | Egypt | Africa | [56] | |
| Ht | − | − | NA | Cyprus | Europe | [148] | |||
| Ht | − | NA | Herbarium specimen records | Cyprus | Europe | [57] | |||
| Ht | − | NA | Canker | US—California | North America | [115,131] | |||
| Nd | ITS, LSU, tef1, tub2 | + | Shoot blight, branch dieback, and canker | Turkey | Asia | [31] | |||
| Prunus avium | Cherry | Nn | ITS, tef1 | + | Canker and branch dieback | Turkey | Asia | [163] | |
| Prunus domestica | Plum | Ht | − | NA | Gummosis and dieback | Egypt | Africa | [56] | |
| Nd | ITS | + | Decline and dieback | Tunisia | Africa | [29] | |||
| Nn | ITS, tef1 | + | Branch dieback and stem cankers | Turkey | Asia | [164] | |||
| Prunus dulcis (P. amygdalus) | Almond | Ht | − | + | Secondary canker infection | US—California | North America | [165] | |
| Ht | − | NA | Canker | US—California | North America | [115,131] | |||
| Nd | ITS, tef1, tub2 | + | Trunk and branch cankers, spur and shoot blight, fruit rot | US—California | North America | [30] | |||
| Nn | ITS, tef1 | + | Stem canker and branch dieback | Turkey | Asia | [32] | |||
| Nd | ITS, tef1, tub2, GPD | + | Trunk and branch cankers | US—California | North America | [33] | |||
| Prunus persica | Peach | Ht | − | − | Canker | US—California | North America | [115,131] | |
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Prunus sp. | Prunus | Fd | ITS, tef1, tub2 | + | NA | Egypt | Africa | [58,134,166] | |
| Pyrus communis | Pear | Nn | ITS, LSU, tef1 | + | Shoot blight and branch canker | Turkey | Asia | [167] | |
| Rutaceae | Citrus aurantifolia | Acid lime | Ht | − | NA | NA | US—California | North America | [115,131] |
| Nd | ITS | + | Root rot | Oman | Asia | [46] | |||
| Nh | ITS, tef1, tub2 | + | Canker and dieback | Iran | Asia | [168] | |||
| Nn | ITS, LSU, tef1 | − | Canker and dieback | Iran | Asia | [40] | |||
| Citrus clementina | Clementine | Nd | ITS, tub2 | + | Shoot blight | Jordan | Asia | [169] | |
| Citrus latifolia | Persian lime | Ht | − | NA | NA | US—California | North America | [115,131] | |
| Citrus limetta | Sweet limetta | Nh | ITS, tef1, tub2 | + | Canker and dieback | Iran | Asia | [168] | |
| Nn | ITS, LSU, tef1 | − | Dieback | Iran | Asia | [40] | |||
| Citrus limettioides | Sweet lime | Nd | ITS | + | Root rot | Oman | Asia | [46] | |
| Citrus limon (C. limonium) | Lemon | Ht | − | − | NA | Cyprus | Europe | [148] | |
| Ef | − | NA | NA | US—California | North America | [129] | |||
| Ht | − | NA | NA | US—California | North America | [115,131] | |||
| Nh | ITS, tef1, tub2 | + | Branch canker | US—California | North America | [20] | |||
| Citrus maxima (C. grandis) | Pomelo | Ht | − | NA | NA | US—California | North America | [131] | |
| Ht | − | NA | NA | US—California | North America | [115] | |||
| Nd | ITS, tub2 | + | Shoot blight | Jordan | Asia | [169] | |||
| Citrus meyerii | Meyer lemon | Ht | − | NA | NA | US—California | North America | [131] | |
| Citrus paradisi | Grapefruit | Ht | − | NA | Canker, dieback | US—California | North America | [115,131,170] | |
| Ef | − | NA | NA | US—California | North America | [129] | |||
| Nd | ITS, tef1, tub2 | + | Bot gummosis | US—California | North America | [171] | |||
| Nh | ITS, tef1, tub2 | + | Branch canker | US—California | North America | [20] | |||
| Nd | ITS, tub2 | + | Shoot blight | Jordan | Asia | [169] | |||
| Citrus reticulata | Mandarin | Ht | − | NA | NA | US—California | North America | [115,131] | |
| Citrus sinensis | Sweet orange | Ht | − | NA | NA | South Africa | Africa | [172] | |
| Ht | − | NA | NA | US—California | North America | [115,131] | |||
| Ef | − | NA | Herbarium specimen records | Pakistan | Asia | [57] | |||
| Nd | ITS | + | Blight, canker, and gummosis | Italy | Europe | [19] | |||
| Nn | ITS, LSU, tef1 | − | Dieback | Iran | Asia | [40] | |||
| Citrus sp. | Citrus | Td | − | NA | NA | US—California | North America | [129] | |
| Ht | − | NA | NA | US—California | North America | [115,129] | |||
| Citrus tangelo | Tangelo | Ht | − | NA | NA | US—California | North America | [115,131] | |
| Salicaceae | Populus alba | Silver poplar | Ht | − | − | NA | Cyprus | Europe | [148] |
| Ht | − | NA | Herbarium specimen records | Cyprus | Europe | [57] | |||
| Populus fremontii | Frémont’s cottonwood | Ht | − | NA | NA | US—California | North America | [173] | |
| Populus nigra | Black poplar | Nh | tef1 | + | Decline | Iran | Asia | [43] | |
| Salix alba | White willow | Nh | tef1 | + | Decline, irregular and central wood necrosis | Iran | Asia | [43] | |
| Nd | ITS, LSU | + | Dieback, shoot blight, and branch canker | Turkey | Asia | [44] | |||
| Solanaceae | Capsicum annuum | Pepper | Ht | − | NA | NA | Tanzania | Africa | [99] |
| Solanum lycopersicum | Tomato | Nd | ITS, LSU, tef1 | + | Blight and root rot | Turkey | Asia | [47] | |
| Tomato | Nn | ITS, LSU, tef1 | + | Stem blight | Turkey | Asia | [174] | ||
| Solanum tuberosum | Potato | Nd | ITS, LSU, tef1 | + | Tuber rot | Turkey | Asia | [175] | |
| Ulmaceae | Ulmus sp. | Elm | Nh | ITS, tef1 | + | Decline | Iran | Asia | [176] |
| Vitaceae | Vitis vinifera | Grapevine | Ht | − | NA | Drying | India | Asia | [89,177,178] |
| Ht | − | NA | Branch wilt | Iraq | Asia | [179] | |||
| Nd | ITS | + | Dieback | Iraq | Asia | [21] | |||
| Nd | ITS, tub2 | + | Wood canker and decline | US—California | North America | [22] | |||
| Nh | ITS, tef1 | + | Dieback | Brazil | South America | [23] | |||
| Nd | ITS, LSU, tef1, tub2 | + | Canker and dieback | Turkey | Asia | [24] | |||
| Nn | ITS, tef1 | + | Wood canker | Turkey | Asia | [25] | |||
| Nh | ITS | + | Wood necrosis | Iran | Asia | [26] |
In the ‘Identified species name’ column: Td for Torula dimidiata, Ht for Hendersonula toruloidea, Ef for Exosporina fawcettii, Sh for Scytalidium hyalinum, Sd for Scytalidium dimidiatum, Fd for Fusicoccum dimidiatum, Nd for Neoscytalidium dimidiatum, Nn for Neoscytalidium novaehollandiae, Nh for Neoscytalidium hyalinum, and No for Neoscytalidium orchidacearum. In the ‘Target loci for identification’ column: ITS: Internal Transcribed Spacer region, tef1: Translation Elongation Factor 1-alpha gene, tub2: Beta-tubulin gene, LSU: large subunit (of ribosomal RNA) gene, SSU: small subunit (of ribosomal RNA) gene, chs-1: chitin synthase 1, act: Actin gene, rpb2: RNA Polymerase II Second Largest Subunit gene, and GPD: Glyceraldehyde-3-Phosphate Dehydrogenase gene. In the ‘Koch’s Postulates’ column, ‘−’ indicates unfulfilled Koch’s postulates, signifying a lack of demonstrated pathogenicity; ‘+’ indicates fulfilled Koch’s postulates, confirming demonstrated pathogenicity. In the context of this dataset, ‘NA’ indicates that the information is not available.
The distribution of N. dimidiatum extends across 37 countries, highlighting its adaptability and prevalence across diverse geographic regions. Iran records the highest number of occurrences, with 47 documented instances, emphasizing a significant presence in the region. Turkey closely follows with 30 reported occurrences, highlighting its substantial distribution. Brazil reports 14 instances, underlining the pathogen’s presence in South America. In Asia, Malaysia and China contributed 12 and 6 reports, respectively, indicating a significant distribution. The United States, particularly in California, displays a substantial presence, with 38 occurrences. Various other regions, including Canada, Colombia, Ecuador, Greece, Israel, Italy, Jamaica, Kenya, Mexico, Niger, the Philippines, Portugal, Puerto Rico, Sierra Leone, Solomon Islands—Rendova, Tanzania, Tunisia, the United Arab Emirates, US—Florida, US—Washington, and US—West Virginia, each report a limited number of cases. Notably, countries in the Middle East, such as Iraq and Oman, document six and nine occurrences, respectively, indicating a significant regional presence. This comprehensive distribution analysis underscores the global adaptability of N. dimidiatum and highlights the need for further research to understand the factors influencing its distribution and to develop effective management strategies.
4.2. Diversity of Host Responses and Geographic Distribution Patterns of Neoscytalidium dimidiatum across Plant Families and Countries
Neoscytalidium dimidiatum has the ability to infect various parts of its host plants, resulting in a wide range of symptoms, including decline, diebacks of branches and limbs, branch wilting, cankers on stems and branches, sooty cankers, wood cankers leading to necrosis, blights affecting spurs and shoots, needle blights, leaf blights, leaf spots, leaf scorching, blights affecting all above-ground plant parts, gummosis, loss of graft union viability, root rot, black root rot, dry rot in stem cuttings, collar rot, stem end rots after harvest, stem rots, fruit rots, internal brown or black stem and fruit rots, tuber rots, lamination of trunk bark, tip rot, as well as instances of asymptomatic conditions. However, the symptoms induced by the pathogen may resemble those caused by other biotic agents (fungi, bacteria, etc.) or abiotic factors. Identification of N. dimidiatum solely through visual examination of its host plants is unlikely, as the fruiting structures of the pathogen’s Fusicoccum-like (pycnidia with conidia) and/or Scytalidium-like synanamorphs (arthrocondia or phragmospores) found on symptomatic plant tissues exhibit morphological similarities to those of other fungal species within the family Botryosphaeriaceae. Moreover, the pathogen can exist in a quiescent or latent state within asymptomatic hosts. Consequently, alternative diagnostic methods beyond visual inspection are necessary for the accurate detection of N. dimidiatum. However, we include all synonym and basionym names of the pathogen to search the literature for the below host range since there is no similar extensive report to support our understanding of host diversity. In order to comprehensively understand the diversity of hosts, we included all synonymous and basionym names of the pathogen while performing literature searches for the host range. This is necessary as there is a lack of thorough reports on this topic. In recent publications, all identified species have undergone phylogenetic analyses based on DNA datasets.
Cankers and the development of internal brown or black rot caused by N. dimidiatum represent significant concerns affecting the stems (cladodes) and fruits of dragon fruits (pitahayas) within the Selenicereus genus (including S. megalanthus, S. monacanthus, S. polyrhizus, and S. undatus) belonging to the Cactaceae family. This phenomenon has been observed across various regions, encompassing China, India, Israel, Malaysia, the Philippines, Taiwan, Thailand, Florida, Puerto Rico, and Ecuador. Plants affected by N. dimidiatum canker exhibit poor recovery as the initial canker spots progress to stem rot, as reported by Chuang et al. [5]. The presence of N. dimidiatum in fruits leads to decay, resulting in unsatisfactory fruit pulp quality, according to Ezra et al. [8]. These collective studies contribute to a comprehensive understanding of the challenges posed by cankers and internal rot in dragon fruit cultivation. These challenges have garnered significant attention due to their influence on the overall health and vitality of these dragon fruit species. Feijo et al. [101] also identified N. dimidiatum as the cause of squamous cladode spots on Nopalea cochenillifera cactus (Cactaceae), representing the first worldwide record of its presence on this host (for all relevant references throughout this chapter, see Table 1).
Neoscytalidium dimidiatum causes canker-related symptoms on a variety of Ficus species belonging to the Moraceae family, including F. benghalensis, F. benjamina, F. carica, F. nitida, F. religiosa, and F. retusa. When cankers appear, the overall health of the plant declines rapidly, as evidenced by the death of leaves and branches. Sooty cankers are observed to form on banyans (F. benghalensis and F. nitida), bodhi trees (F. religiosa), and weeping fig (F. benjamina). These cankers can be distinguished by their dark and encrusted external appearance [37,39,40]). Bark necrosis, the presence of cankers on branches and aerial roots, yellowing of foliage, defoliation, and branch dieback are further signs that have been identified in these plants. In the case of common figs (F. carica), it is observed that the occurrence of cankers on branches is a frequent phenomenon, leading to the manifestation of dieback symptoms and a subsequent reduction in the overall health of fig trees. These symptoms, if left unaddressed, can ultimately result in the demise of the affected trees. Neoscytalidium dimidiatum on Ficus spp. exhibits a broad geographical zone of influence, encompassing many regions, including Egypt, Oman, Iran, Turkey, Brazil, California, and Mexico. Neoscytalidium dimidiatum also leads to shoot and branch deaths as well as sooty canker in Morus spp. (M. alba, M. australis, and M. nigra) of the same family (Moraceae), spanning across regions including Iran, Pakistan, Turkey, and the US.
Symptoms attributed to N. dimidiatum across diverse citrus hosts encompass a spectrum of manifestations, including canker formation, dieback, shoot blight, branch canker development, bot gummosis, gummosis occurrences, as well as root rot, affecting acid lime (C. aurantifolia), clementine (C. clementine), sweet limetta (C. limetta), sweet lime (C. limettioides), lemons (C. limon), pomelo (C. maxima), grapefruit (C. paradise), and sweet orange (C. sinensis) within the Citrus spp. domain of the Rutaceae family. These symptoms of Citrus spp. have been documented across various geographic regions, including Oman, Iran, Jordan, Pakistan, California, Cyprus, Italy, and Southern Africa.
Symptoms associated with N. dimidiatum on apple trees (Malus domestica), a member of the Rosaceae family, encompass branch dieback, branch canker, and gummosis [180]. These manifestations have been observed within distinct geographical locales, namely Egypt, Iran, India, Iraq, China, and Turkey. Within the sphere of stone fruit trees (Prunus spp.) in the Rosaceae family, a manifold spectrum of pathological manifestations becomes apparent. These include spur and shoot blights, branch dieback, trunk and branch cankers, decline, sooty canker, as well as secondary canker infections. Notably, this intricate array of symptoms is observed in apricots (P. armeniaca), cherries (P. avium), almonds (P. amygdalus), and plums (P. dulcis). These observations have been exhaustively documented across a geographically extensive scope spanning Egypt, Tunisia, Turkey, Iran, California, and Cyprus. The presence of N. dimidiatum on pear trees (Pyrus communis) within the Rosaceae family has been documented in Turkey, where it has been found to be responsible for the development of shoot blight and branch canker.
Neoscytalidium dimidiatum’s influence on walnuts (Juglans spp.), encompassing both J. regia and J. californica of the Juglandaceae family, includes black canker, root rot, graft union failure resulting in death, overall decline, sooty canker formation, as well as instances of branch wilt. These symptoms have been documented within geographical regions spanning Iran, Turkey, and California.
Within the Pinaceae family, N. dimidiatum has been associated with the occurrence of shoot and needle blight, concomitant with dieback, across diverse species of pine trees (Pinus spp.), including P. eldarica, P. nigra, and P. sylvestris. These occurrences have been observed in both Turkish and Iranian settings. The phenomenon of needle blight has been documented in blue spruce (Picea pungens) in Turkey.
Members of the Anacardiaceae family, namely mango (Mangifera indica) and pistachio (Pistacia vera), exhibit susceptibility to the influence of N. dimidiatum. Mango trees manifest a diverse spectrum of symptoms, encompassing dieback, the formation of stem and branch cankers, as well as the occurrence of leaf spots, evident across extensive geographic regions spanning Brazil, South Africa, Niger, Australia, California, and India. Similarly, pistachio trees within the territories of Turkey and Iran display a varied range of conditions, encompassing canker, shoot blight, and root rot, as well as instances of asymptomatic occurrences.
Neoscytalidium dimidiatum induces leaf blight in the white spider lily (Hymenocallis littoralis) and natal lily (Clivia miniata) of the Amaryllidaceae family and dracaena (Dracaena trifasciata) of the Asparagaceae family, with occurrences documented in Malaysia and Iran. Additionally, this fungus is associated with black leaf spots on sisal plants (Agave sisalana) of the Asparagaceae family, located in China. Notably, N. dimidiatum also prompts leaf blight and leaf spot on Cattleya orchids (Cattleya × hybrid, C. lueddemanniana var. lueddemanniana) within the Orchidaceae family in Taiwan and Thailand. Moreover, N. dimidiatum triggers leaf scorch in olive trees (Olea europaea) of the Oleaceae family in Turkey. This decline initiates with initial foliar scorching, which progresses to dieback in twigs, branches, and even entire trees. As the disease advances, necrosis becomes evident, accompanied by the formation of cankers on trunks, branches, and twigs. N. dimidiatum induces tip rot and leaf spot in bananas (Musa spp.: M. acuminata and M. nana) of the Musaceae family, noted in Hawaii and Jamaica but solely recorded in checklists. Additionally, foliar and stem blight, along with root rot, affect tomatoes (Solanum lycopersicum), and tuber rot impacts potatoes (S. tuberosum), both belonging to the Solanaceae family in Turkey.
Postharvest stem end rot and leaf spot in pineapple (Ananas comosus) within the Bromeliaceae family in Malaysia are among the symptoms of N. dimidiatum. Both Brazil and Malaysia have the challenge of dealing with root and stem rots that impact sweet potatoes (Ipomoea batatas) from the Convolvulaceae family. Fruit rot is observed in melons (Cucumis melo) belonging to the Cucurbitaceae family, with a particular occurrence in Iran. Yams belonging to the Dioscoreaceae family, such as Dioscorea esculenta and Dioscorea rotundata, experience dieback and tuber dry rot in Iran and Colombia caused by N. dimidiatum. In Turkey, it has been shown that Japanese persimmons (Diospyros kaki) belonging to the Ebenaceae family are susceptible to branch dieback caused by N. dimidiatum. Within the Euphorbiaceae family, N. dimidiatum has been observed to cause collar and root rot in the physic nut plant (Jatropha curcas) in Brazil. Cassava (Manihot esculenta), a member of the Euphorbiaceae family, is confronted with the challenges posed by black root rot and stem-cutting dry rot, which have been observed in several regions, such as Thailand, Kenya, Ghana, and Brazil. In Iran, barberry (Berberis vulgaris) shrubs in the Berberidaceae family exhibit symptoms of canker and severe dieback due to N. dimidiatum.
There are a significant number of studies that delve into the host diversity of N. dimidiatum within the Fabaceae family. The impact of this pathogen within the family is exemplified by the manifestation of dieback and sooty canker on the siris tree (Albizia lebbeck), observed in regions including Iran and Oman. Neoscytalidium dimidiatum’s effect extends to the orchid tree (Bauhinia purpurea) bark in India, where infection leads to the occurrence of lamination. Canker is a notable affliction affecting both the golden shower tree and arsenic bush (Cassia spp., particularly C. fistula and C. floribunda) in Iran. Chickpea plants (Cicer arietinum) in Turkey are susceptible to N. dimidiatum infection, resulting in blight and root rot. Dieback and stem canker are prevalent issues observed in the United Arab Emirates and Oman concerning the royal poinciana (Delonix regia). Similarly, documented instances of dieback occur in copperpod trees (Peltophorum petrocarpum) in Oman. The phenomenon of dieback also extends to Parkinsonia aculeata, commonly known as Palo verde trees, located in Australia. This occurrence follows the application of N. dimidiatum as a bioherbicide. Nevertheless, we firmly advocate against the utilization of this potentially hazardous pathogen as a herbicide.
In the context of the Fagaceae family, investigations into the host diversity of N. dimidiatum reveal distinct findings. Notable occurrences encompass branch and trunk cankers observed on oriental beech (Fagus orientalis) in Iran. Persian oak (Quercus brantii) in Iran demonstrates a complex presentation, including dieback, decline, and sooty canker. Moreover, canker affects sweet chestnut (Castanea sativa); however, its documentation is solely based on checklist inclusion.
Within the Lamiaceae family, N. dimidiatum exhibits diverse manifestations. Lavender (Lavandula angustifolia) in Turkey encounters foliar and stem blight. Lemon balm (Melissa officinalis) experiences blight, while Turkish oregano (Origanum onites) suffers from leaf blight in the same region. Common sage (Salvia officinalis) faces the combined threat of root rot and foliar blight in the Lamiaceae family context in Turkey.
Within the Myrtaceae family, N. dimidiatum exhibits a spectrum of host responses, including dieback, trunk bark lamination, canker, sooty canker, and asymptomatic occurrences within Eucalyptus species (E. camaldulensis and other unidentified species) in regions spanning Iran, Iraq, Portugal, and Australia. Guava (Psidium guajava) shows postharvest fruit rot, dieback, and stem and branch cankers across Malaysia, India, and Brazil. Similarly, Java plum (Syzygium cumini) manifests trunk bark lamination, cankers, and wedge-shaped wood necrosis alongside latent conditions in Iran. Furthermore, N. dimidiatum induces canker in the weeping bottlebrush (Callistemon viminalis) in Iran. This array of symptoms underscores the intricate influence of N. dimidiatum within the Myrtaceae family.
Within the Salicaceae family, N. dimidiatum induces decline in poplar trees (Populus spp.: P. alba, P. fremontii, and P. nigra), evident across regions including Iran, California, and Cyprus. Moreover, this pathogen is associated with a spectrum of symptoms in willow trees (Salix alba) within the same family, encompassing decline, irregular and central wood necrosis, dieback, shoot blight, and branch canker, primarily observed in Iran and Turkey.
Neoscytalidium dimidiatum’s impact is diverse across various plant species, resulting in a range of symptoms. For instance, it leads to elm decline (Ulmus sp.) within the Ulmaceae family in Iran. In the Vitaceae family, grapevine (Vitis vinifera) experiences canker, dieback, wood canker, necrosis, decline, and branch wilt, spreading through regions like Iran, Iraq, Turkey, India, California, and Brazil. Similarly, in the Rhizophoraceae family, red mangrove (Rhizophora mucronata) suffers from canker and dieback in Iran. N. dimidiatum triggers sooty canker and necrotic wood tissues in pomegranate (Punica granatum) of the Lythraceae family in Iran, canker and dieback in chinaberry (Melia azedarach) of the Meliaceae family in Iraq and Pakistan, and decline in neem (Azadirachta indica) of the Meliaceae family in Iran. Additionally, N. dimidiatum causes lamination of trunk bark in buttonwood (Conocarpus erectus) within the Combretaceae family in Iran. Moreover, cankers and dieback affect white mangrove (Avicennia marina) in the Acanthaceae family, evident in Iran, as well as dieback of the portia tree (Thespesia populnea) of the Malvaceae family in Oman. N. dimidiatum also induces branch and trunk cankers in both common alder (Alnus glutinosa) and common hornbeam (Carpinus betulus) of the Betulaceae family in Iran. Certain reports predominantly concentrate on the isolation and identification of field symptoms, while the investigation into pathogenicity aspects remains largely unexplored. In this context, N. dimidiatum was isolated from canker-infected plants, namely Assyrian plum (Cordia myxa) of the Boraginaceae family, oleander (Nerium oleander) of the Apocynaceae family, Christ’s thorn jujube (Ziziphus spina-christi) within the Rhamnaceae family, Mediterranean cypress (Cupressus sempervirens) of the Cupressaceae family, and Chinese hibiscus (Hibiscus rosa-sinensis) of the Malvaceae family, all originating from Iran. In Italy, there has been a documented case of branch canker and dieback in meryta (Meryta denhamii) (Araliaceae) attributed to N. dimidiatum. This diverse array of symptoms underscores the fungus’s varied impact.
Remarkably, some hosts exhibit an intriguing phenomenon of asymptomatic conditions, underscoring a distinctive interaction between the pathogen and the host. Notably, these conditions deviate from the conventional endophytic or epiphytic manifestations. Within this context, a noteworthy observation pertains to six genera that serve as symptomless hosts. This exclusive group encompasses acacias (Acacia spp.) (Fabaceae), rattlepods (Crotalaria medicaginea) (Fabaceae), Kimberley bauhinia (Lysiphyllum cunninghamii) (Fabaceae), baobabs (Adansonia spp.) (Malvaceae), and blue grevillea (Grevillea agrifolia) (Proteaceae) in Australia, alongside quiver tree (Aloidendron dichotomum) (Asphodelaceae) in Africa. Given the intricate dynamics and uncertainties surrounding the pathogen’s role, we have chosen to identify these hosts as symptomless, as the exact nature of their interaction with N. dimidiatum remains to be elucidated.
In summary, N. dimidiatum demonstrates extensive host diversity, involving 84 distinct plant genera and 126 species distributed across 46 families, which encompass 34 eudicot angiosperms, 9 monocot angiosperms, and 3 gymnosperms. It is noteworthy that among these genera, 10 (Rhus, Thaumatophyllum, Agathis, Araucaria, Furcraea, Casuarina, Sequoiadendron, Gladiolus, Arachnis, and Capsicum) lack descriptions regarding the induction of symptoms. Additionally, six genera have been identified as hosting the pathogen in an epiphytic or endophytic capacity without resulting in discernible symptoms. Conversely, detailed investigations have established the symptomatic relationship between N. dimidiatum and 68 genera, providing comprehensive insights into this aspect. Furthermore, 18 species (Rhus typhina, Thaumatophyllum bipinnatifidum, Agathis robusta, Araucaria sp., Agave sp., Furcraea foetida, Sansevieria hyacinthoides, Casuarina sp., Sequoiadendron giganteum, Gladiolus sp., Arachnis sp., Citrus latifolia, Citrus meyerii, Citrus reticulata, Citrus tangelo, Populus alba, Populus fremontii, and Capsicum annuum) have been recognized as potential hosts, primarily based on checklist inclusion, although specific symptom definitions remain unavailable for these species.
While Neoscytalidium species are commonly known as phytopathogens, they can manifest in various clinical conditions, affecting individuals with underlying factors or even those without apparent health issues. Despite typically causing superficial skin and nail infections, these species have been linked to more severe conditions like cerebral phaeohyphomycosis [4]. Notably, Heidari et al. [3] explored N. dimidiatum’s presence in human respiratory tracts, highlighting its adaptability to different environments and raising important questions about its impact on both human and plant health. This underscores the need for further research into the potential risks and implications associated with N. dimidiatum in various settings.
5. Understanding the Epidemiological Aspects of Neoscytalidium dimidiatum
5.1. Life Cycle of Neoscytalidium dimidiatum
The life cycle of N. dimidiatum presents intriguing dynamics, yet it remains a subject of ongoing research with several complex facets. While the epidemiology, including seasonal dynamics of different spore types of Neoscytalidium, is still inadequately understood, its impact on various crops and the occurrence of epidemics have been noted [11,27,77].
The interactions between this pathogen and its host plants are multifaceted and not fully elucidated [181]. Moreover, the virulence mechanisms employed by N. dimidiatum are still being explored.
As a plant pathogen, N. dimidiatum employs a dual strategy for asexual reproduction, as documented in various studies [2,50,51,52,53]. Firstly, it reproduces through hyphal fragmentation, where discrete fragments of the fungus detach, serving as vehicles for its dissemination to new host plants. Secondly, N. dimidiatum utilizes specialized structures known as fruiting bodies, within which it generates spores. These spores are subsequently released into the environment, poised for infecting new plant hosts. This intricate combination of asexual reproduction mechanisms equips Neoscytalidium with a remarkable capacity for efficient spread and establishment across diverse plant populations.
Crucially, Neoscytalidium’s adaptability underpins its propensity to initiate widespread diseases in agricultural and horticultural settings, presenting significant challenges in plant health management. It is worth noting that N. dimidiatum can infect its hosts through various means, including wounds and natural openings. Additionally, direct penetration can occur on specific host plants, such as dragon fruits, through the formation of appressoria, particularly on young tissues that are highly vulnerable to infection [45].
In conclusion, N. dimidiatum’s life cycle and pathogenicity represent intricate processes influenced by various factors, including host plants and environmental conditions. Further research is essential to fully comprehend the complexities of its interactions and the mechanisms underlying its adaptability as both a pathogen and a saprotroph.
5.2. Host Infection in a Unique Context of Dragon Fruit Canker
In the context of dragon fruit canker, acknowledged as the most destructive disease affecting dragon fruit plants, the fungal infection commences by generating appressoria on the surface, which is succeeded by a direct penetration into an epidermal cell. Initially, a swift collapse of chlorenchyma cells adjacent to the infection site occurs, accompanied by limited colonization of the tissue during the initial phases. In response to the infection, the host generates callus tissue, which forms around the impaired tissue, thereby curbing the progression of many infections. However, certain infections proceed to manifest extensive spreading lesions that contain embedded pycnidia. Internally, within the dead tissue, hyphal cells separate to give rise to arthrospores. Notably, lesion expansion is often preceded by the formation of prominent yellow halos, suggesting the production of a diffusible toxin by the pathogen. The existence of such a toxin is further corroborated by the chlorosis evident in cladode tissue subsequent to injection with cell-free culture filtrates of the fungus [45].
The most susceptible area for infection is the young tissue located directly behind the growing point of the cladode. The apexes of rapidly growing cladodes are particularly prone to infection. Throughout the inoculation phase, lesions are rarely observed beyond 3 cm from the apex. This specific susceptibility at certain sites explains the distinct bands of symptoms observed on mature cladodes in their natural environment. Each band represents infections that occurred during specific growth phases of the cladodes. Consequently, the highest vulnerability to infection in orchards corresponds to wet periods that coincide with the peak of vegetative growth. On the other hand, mature and aged cladodes demonstrate substantial resistance to infection [45].
Arthroconidia are confined within the necrotic tissues of the lesion and have limited dispersal capability, while pycniospores, released during wet weather and spread by rain, are the primary means of dispersal and infection. Therefore, the disease is more severe during the wet season when infection conditions are frequent [45]. While undocumented, there is a possibility that phragmospores formed on infected host surfaces could be disseminated by wind.
5.3. Factors Influencing Neoscytalidium dimidiatum Infection
5.3.1. Factors Influencing Host Response
Pathogens belonging to the Botryosphaeriaceae family employ distinct modes of infection in plants, exploiting openings such as wounds in both green and woody tissues, as well as utilizing natural entry points in flowers, fruits, leaves, and green shoots. These pathogens employ enzymatic and toxic agents to induce cellular and tissue demise within the afflicted plant organs. In the context of perennial woody hosts, incursions occurring within the inner layers of the wood or immediately beneath the bark can inhibit shoot development, culminating over time in the eventual withering of shoots, commonly referred to as ‘dieback’ [182].
Neoscytalidium dimidiatum, much like other species within the Botryosphaeriaceae family, is categorized as a stress-associated pathogen. Disease manifestation is frequently intertwined with the presence of abiotic stressors, including factors such as drought, physical damage, and unfavorable growing conditions, as substantiated by various studies [33,40,41,46,76,183,184]. The diseases induced by N. dimidiatum predominantly coincide with the onset of these stress factors, which are independent of the N. dimidiatum infection itself. In such scenarios, N. dimidiatum actively contributes to the emergence of observed symptoms, albeit to varying extents, contingent upon the severity and duration of the stress conditions. Notably, these disease symptoms can escalate swiftly, leading to extensive losses over expansive geographic regions when the causative stress factor is pervasive. Furthermore, the convergence of multiple factors, including biological pressures from pathogens and pests that are extending their geographic ranges, may collectively contribute to the conducive environment for the development of N. dimidiatum-induced diseases, as postulated by Slippers and Wingfield [185].
Across a diverse range of woody hosts in tropical and semi-tropical regions, N. dimidiatum functions as a wound-invading pathogen, with a tendency to primarily infect branches and trunks that have experienced damage from sunburn, freezing, or pruning [170]. Its primary focus of infection lies in the cambial region of the tree, leading to symptoms of wilting and branch dieback [186,187]. Subsequently, blisters and fissures develop on the bark, revealing black spore masses that are dispersed by wind. Concurrently, the affected wood exhibits a distinctive brown staining. When introduced into bark wounds extending to the vascular cambium or exposed xylem due to pruning, the pathogen demonstrates the capacity to induce canker formation [165]. These cankers primarily display an annual rather than perennial nature. The pathogen is primarily localized within the cell lumens of the xylem and phloem, with intercellular spread primarily occurring through pits. However, the excision of cankers, whether or not a wound protectant is applied, has proven ineffective in managing the disease [165]. However, pruning and grafting wounds are identified as the probable primary entry point for infection [76,138]. The application of appropriate fungicides or biological control agents to protect these wounds still may have a chance to prevent or minimize the occurrence of such infections. In the study conducted by Çiftçi et al. [138], it was observed that the regrafting of unproductive walnut trees with Chandler, when they were older than 8 to 10 years, resulted in the frequent and severe development of canker symptoms. This practice leads to tree loss in the affected orchards.
In their recent study, Güney et al. [76] observed that cracked tree trunks, branches, and limbs represent the most notable symptoms of canker in all examined olive cultivars. However, the severity and variety of symptoms were significantly influenced by multiple factors, including the cultivar, age of the tree, season, and biotic and abiotic stress factors. While trees of all ages are affected, younger trees were particularly impacted, resulting in higher mortality rates. Symptoms were most noticeable during the summer and fall seasons. Pruned trees, those with injury, ripe fruits, and mechanically injured plant parts appeared to be more susceptible to the pathogen, exhibiting large canker formations or black sporulation on exposed areas. Dying leaves, twigs, branch cankers, and fallen leaves and fruits all play a role in the disease cycle and the survival of the pathogen. Water deficiency and drought were recognized as significant stress factors that increase susceptibility, leading to the exacerbation of disease severity. Yeganeh and Mohammadi [39] investigated the production of loose spores by N. dimidiatum beneath the bark of infected banyan trees and in the pruning debris. This observation aligns with prior research that Botryosphaeriaceae species, known for inducing trunk diseases in fruit and forest trees, can also exist as saprophytes within wood debris [1]. These species can persist and produce spores in wood debris, serving as crucial sources of inoculum for infections in woody plants.
Gusella et al. [41] investigated various aspects of N. dimidiatum’s behavior. Their findings revealed that older branches (>3 years old) exhibit a longer timeline for canker development compared to their younger counterparts. They also made an intriguing observation about the absence of N. dimidiatum within fruit mummies, which are inhabited by various other fungal saprophytes.
5.3.2. Climatic and Edaphic Factors
Holland et al. [32] document that the timing of inoculation significantly impacts the development of N. dimidiatum-induced lesions on almond trees. Lesions are more pronounced when inoculation occurs during the summer compared to smaller lesions resulting from winter inoculation. These findings underscore the pathogen’s adaptability to elevated temperatures, as exposure to high temperatures can lead to substantial plant damage. Researchers such as Gusella et al. [75], Güney et al. [76], Hohenfeld et al. [73], Hong et al. [11], Nouri et al. [30], and Sabernasab et al. [74] have consistently reported the optimal temperature range for N. dimidiatum’s growth, sporulation, conidial germination, and pycnidia formation to be between 30 and 36 °C. Notably, Güney et al. [49] have conducted in vitro analyses, revealing that conidial germination reaches its zenith at 40 °C. Likewise, Coutinho et al. [72] demonstrated that Neoscytalidium is capable of thriving at a high temperature of 40 °C. Hohenfeld et al. [73] have particularly concentrated on the black root rot induced by N. dimidiatum on cassava, elucidating that the temperature range of 32–36 °C yields the highest disease severity. Hassan et al. [188] observed that the susceptibility of saplings to stem canker caused by N. dimidiatum is influenced by the duration of warm temperatures and the abundance of fungal inoculum. To further explore the impact of temperature conditions on canker development, a growth chamber was utilized with two temperature regimes: a very hot condition at 40 °C and a hot condition at 32 °C. Among the pre-inoculation regimes, the very hot and hot temperatures were identified as the most favorable for infection in eucalyptus saplings when compared to other host species. Mayorquin et al. [20] conducted a temperature study on lesion development caused by the pathogen, supporting these findings by showing that increased temperatures lead to more aggressive infection by this pathogen. Machado et al. [118] and Adesemoye et al. [171] observed that N. dimidiatum causes disease in semi-arid and desert regions, while its presence is absent in more humid areas. These findings and observations further accentuate the detrimental effects of elevated temperatures on plant vitality, indicating the pathogen’s thermotolerant nature.
Gusella et al. [41] demonstrated in laboratory experiments that N. dimidiatum efficiently colonizes desiccated fig fruits within a temperature range of 20 °C to 35 °C. Additionally, their in vitro studies on water potentials reveal a direct correlation between the mycelial growth rate and decreasing water potential (ranging from 1 to 3 MPa), with the specific type of salt used further influencing this reduction. In a separate study by Kuan et al. [184], they extensively explored the metabolic attributes of N. dimidiatum, including nutritional profiling, pH tolerance, and osmotolerance. Their research emphasized that the fungus exhibits optimal growth within a pH range of 4.5–5.5, highlighting the importance of future investigations into substrate utilization under varying growth conditions.
5.4. Mode of Transmission
Similar to other members of Botryosphaeriaceae, the dissemination of this pathogen occurs through various means such as wind, rain, and diverse arthropod groups [39,165,176,186,187,189]. Jiménez Luna et al. [48] carried out a comparison of fungal community compositions obtained from spore traps and those acquired from plant samples collected from symptomatic almond and walnut trees. Their study consistently confirmed the identification of four distinct species, including N. dimidiatum, using both methodologies across various orchards. This discovery highlights the crucial role of airborne spores in the epidemiology of N. dimidiatum and strengthens the reliability and efficacy of these detection techniques. During our pathogenicity assays involving N. dimidiatum on various plant species, we made an unpublished observation of viable plant colonization by dust particles originating from air conditioner filters (personal communication with our research team member Şahimerdan Türkölmez). This occurrence is attributed to direct and prolonged exposure to air-conditioning and air-circulating systems within our growth rooms. Interestingly, Heidari et al. [3] conducted a study focusing on N. dimidiatum within human respiratory tracts. The ability of N. dimidiatum to thrive in such diverse environments, ranging from plants to human tissues, emphasizes its adaptability and raises questions about its potential impact on both human and plant health.
Al-Sadi et al. [46] emphasized the significance of nurseries and the potential spread of the pathogen through planting infected nursery plants. Türkölmez et al. [47] suggested that N. dimidiatum can potentially be transmitted through seeds, soil, and/or the air, and it may persist in the soil in association with infected tomato debris. While Mirtalebi et al. [107] and Güney et al. [49] have demonstrated the pathogen’s capacity to colonize the seed coat, the potential for N. dimidiatum to spread through the seeds of its host plants remains uncertain. Yeganeh and Mohammadi [39] have provided evidence that the pathogen’s conidia can also be transported by arthropods. In their study, Zhu et al. [189] successfully isolated N. dimidiatum from the beetle gallery system within infested elm (Ulmus densa) stems in China. This finding suggests the potential occurrence of N. dimidiatum within beetle galleries, a common habitat for various fungi, highlighting the ecological relevance of N. dimidiatum in this specific context. Considering that the beetle assists in carrying its conidia, it may indirectly aid in their dissemination. Avian species, rodents, and other small animals may serve as potential vectors for disseminating the pathogen through infected fruits and seeds [190].
During a survey on mango conducted by Ray et al. [35], N. hyalinum was also isolated from nearby fig shrubs (Ficus carica), indicating the potential movement of the pathogen between various plant hosts. Esmaeili et al. [98] observed canker and severe dieback symptoms on barberry trees in specific areas of Fars Province, Iran. Some of these trees were situated in vineyards that had been previously studied for grapevine trunk diseases. The researchers aimed to investigate whether barberry trees could serve as hosts for fungal trunk pathogens affecting grapevines. They collected wood samples from infected barberry trees and isolated fifteen fungal species, including Phaeoacremonium spp., D. seriata, N. dimitatum, and B. dothidea. The pathogenicity tests on both barberry and grapevine revealed that the isolates collected from barberry trees demonstrated cross-pathogenicity on grapevines. Their study underscores that barberry trees can act as alternative hosts for diverse grapevine fungal trunk pathogens, including N. dimidiatum.
6. Insights into the Virulence and Genomic Characteristics of Neoscytalidium dimidiatum
6.1. Host–Pathogen Interaction at Molecular Level
The virulence of Botryosphaeriaceae pathogens relies on an array of molecules known as effectors, including enzymes responsible for cell wall degradation, secondary metabolites, and peptidases [191]. Next-generation sequencing (NGS) has revolutionized fungal research within the Botryosphaeriaceae family, revealing unique gene and repeat distributions linked to co-expression patterns [192]. These pathogens employ a multifaceted biochemical arsenal to manipulate host physiology. This includes the synthesis of polyketide toxins like terremutin and mellein, known for their roles in defense mechanisms and necrosis induction [193,194]. They also rely on extracellular proteins [195], secreted polysaccharides [196], and jasmonic esters [197] to facilitate pathogenicity. Central to their arsenal are carbohydrate-active enzymes (CAZymes), particularly glycoside hydrolases (GH), which enable the degradation of wood components like cellulose and hemicelluloses [182,192,198]. While GH enzymes hold biotechnological promise and are potential phytoprotection targets, their precise in vivo mechanisms remain only partially understood. Other CAZyme categories include glycosyltransferases, carbohydrate esterases, polysaccharide lyases, and auxiliary activities, often associated with carbohydrate-binding modules [199]. The current understanding of proteases produced by Botryosphaeriaceae and their likely vital roles in fungal physiology and pathogenesis is very limited [192,198].
In recent years, N. dimidiatum has garnered increased attention through extensive research endeavors. Despite its importance, the population genetics of N. dimidiatum remains relatively unexplored, with limited studies providing insights into its virulence and genomic characteristics [182,184,191]. This pathogen exhibits significant variation in virulence across species within the Botryosphaeriaceae family, with unique attributes, such as mannanases, α-glucosidase enzymes, and pectate lyases, setting N. dimidiatum apart. These unique enzymatic traits, especially GH76 and PL3 proteins, are notably prevalent in N. dimidiatum compared to other species [182].
Genomic analyses have unveiled putative coding DNA sequences, including genes related to melanin biosynthesis, a known contributor to fungal virulence and adaptation to diverse environments. These analyses also reveal genes associated with iron uptake, which is crucial for various fungal cellular processes. The fungus displays a set of CAZymes linked to plant pathogenicity, involving the decomposition of hemicellulose and pectin [184].
Moreover, investigations into host–pathogen interactions have identified key genes associated with plant defense responses and calcium ion signaling pathways in red-fleshed pitaya [181,200]. These studies emphasize the potential roles of specific genes, including PR homologous proteins and WRKY transcription factors, in regulating pitaya’s defense mechanisms against N. dimidiatum infection. Transcriptomic analyses have also revealed the presence of Leucine-Rich Repeat (LRR) genes, primarily within the LRR-RLK, NBS-LRR, and FBXL subfamilies, underscoring their involvement in plant defense mechanisms against N. dimidiatum. These genetic insights contribute to our understanding of the molecular responses of pitaya to N. dimidiatum infection, highlighting potential key players in plant defense. Notably, research by Wang et al. [201] on reference genes for accurate gene expression analysis in N. dimidiatum offers valuable guidance for future studies, facilitating more precise investigations into the gene expression dynamics of this pathogen under varying conditions.
Collectively, these studies enhance our understanding of N. dimidiatum’s biology, genetics, and interactions with host plants. Continued research in this area holds the promise of further elucidating the mechanisms underpinning its pathogenicity and host interactions, ultimately aiding in the development of effective strategies for disease management and control.
6.2. Neoscytalidium dimidiatum Virulence in Diverse Host Plants
In the intricate realm of plant–pathogen interactions, understanding the virulence and impact of fungal species becomes paramount for effective disease management. N. dimidiatum, a member of the Botryosphaeriaceae family and other canker-causing fungi, has garnered considerable attention from diverse research teams for its aggressive tendencies and far-reaching consequences on various hosts. Through a series of studies, researchers have delved into the complexities of this fungal species, shedding light on its behavior, impact, and virulence. The virulence of N. dimidiatum stands out as a central theme across these investigations, positioning it as one of the most aggressive species within its family. The research by Marques et al. [84] into Botryosphaeriaceae fungi associated with mango dieback and stem-end rot in Northeastern Brazil yielded valuable insights. Their exploration identified seven taxa, including N. dimidiatum. Inoculation experiments on mango fruits highlighted the potential of various Botryosphaeriaceae species to induce damage, with N. dimidiatum and Neofusicoccum parvum exhibiting the largest lesions, underscoring their elevated virulence. Mayorquin et al. [20] conducted a study to identify the fungi associated with citrus branch canker and dieback disease in the desert areas of southern California. N. hyalinum, Eutypella citricola, E. microtheca, and an unidentified species of Eutypella were identified. N. hyalinum, the most often recovered fungus, together with Eutypella sp., induced much longer infections than other Eutypella species on ‘Lisbon’ lemon branches. Panahandeh et al. [156] contributed insights through their investigation into pathogenicity tests on different fungal species inoculated into Syzygium cumini shoots. Among the nine species examined, N. hyalinum emerged as a prominent contributor, inducing notably elongated lesions, thus unveiling its distinct impact on the host. A study by Kazemzadeh et al. [1] centered on pathogenicity assessments of nine Botryosphaeriaceae species in diverse forest trees in Iran. Their evaluation highlighted significant disparities in virulence among species, with N. novaehollandiae, Botryosphaeria dothidea, and Diplodia intermedia emerging as particularly aggressive agents. Notably, N. novaehollandiae exhibited heightened virulence on multiple host species, showcasing its adaptability to diverse environments. Ghasemi-Sardareh et al. [145] focused on the identification of fungal species associated with trunk diseases in neem trees (Azadirachta indica). Among an array of identified fungal taxa, N. hyalinum emerged as a potent contributor, displaying a high level of aggressiveness substantiated by the extent of vascular necrosis observed within the wood. In a study conducted by Çiftçi et al. [138] using walnut seedlings cv. Chandler, N. dimidiatum exhibited greater virulence than Lasiodiplodia theobromae, resulting in longer necrotic lesions.
However, contrasting outcomes have arisen concerning the most virulent species across distinct hosts. Chen et al. [136] conducted a pivotal study, isolating ten Botryosphaeriaceae family species, including N. dimidiatum, from varied walnut infections in California. Pathogenicity assessments involving English walnut cultivars (‘Chandler’, ‘Tulare’, and ‘Vim’) demonstrated the pathogenicity of all species, with L. citricola and N. parvum exhibiting the highest pathogenicity, followed by N. mediterraneum, N. dimidiatum, and B. dothidea. Exploring Bot gummosis in California citrus, Adesemoye et al. [171] investigated Bot gummosis infections in California’s citrus crops. In experiments using Eureka lemon shoots, lesion lengths varied based on the employed isolates. Particularly, Neofusicoccum luteum isolates induced longer lesions, while Dothiorella viticola isolates resulted in shorter lesions compared to other species, including N. dimidiatum. Hashemi and Mohammadi [43] studied the decline in willow and poplar trees in Iran, identifying nine fungal species linked to the decline symptoms, including N. hyalinum. Pathogenicity tests demonstrated that L. theobromae caused the longest lesions on willow, while Phaeoacremonium parasiticum induced the longest lesions on poplar. Further, Hashemi et al. [176] addressed elm tree decline, highlighting Dothiorella sarmentorum as the most virulent species based on wood necrosis extent. Coutinho et al. [72] examined Botryosphaeriaceae species associated with cashew, mango, and guava dieback, stem, and branch cankers. All isolates, including N. hyalinum, showed virulence, with Neofusicoccum strains being the most aggressive in mango, cashew, and ‘caja-umbu’ (Spondias mombin × S. tuberosa) plants. Espargham et al. [168] found that L. theobromae was the most virulent species on lime shoots, while Yeganeh and Mohammadi [39] reported the same for banyan trees compared to N. dimidiatum and other species. Brito et al. [122], in a Brazilian study on cassava investigating root and stem dry rot pathogens, N. dimidiatum and nine Lasiodiplodia species were analyzed. Lasiodiplodia laeliocattleyae exhibited the highest virulence, followed by N. dimidiatum and L. parva. Other species showed lower virulence levels, with no significant differences among them. Arkam et al. [202] investigated the Botryosphaeriaceae family, a fungal group with a global impact on grapevine production, under Algerian conditions. They identified eleven species from six genera, including N. dimidiatum. Pathogenicity trials involving grapevine green shoots showed that all identified species exhibited pathogenicity, with Neofusicoccum parvum and L. theobromae being the most aggressive. On barberry, L. theobromae exhibited a higher level of aggressiveness than N. dimidiatum on both barberry and grapevine [98]. In pomegranate, lesions induced by L. theobromae were significantly longer, whereas N. dimidiatum caused larger and deeper wood lesions compared to other species [144].
Ahmadpour et al. [40] reported notable variations in the response of different host species upon the reintroduction of N. novaehollandiae to its original substrates. It is worth mentioning that Conocarpus erectus, Eucalyptus camaldulensis, and Morus nigra had an increased predisposition towards severe wood discoloration accompanied by the appearance of a characteristic black powdery manifestation. In contrast, both Ficus religiosa and Bauhinia purpurea exhibited a somewhat reduced degree of these phenotypic modifications. These findings align harmoniously with the outcomes of concurrent field evaluations, thereby corroborating the presence of divergent patterns in disease incidence and severity within the ambit of the examined arboreal taxa.
In conclusion, the multifaceted investigations into N. dimidiatum and the Botryosphaeriaceae family have unveiled the complex nature of pathogenicity and virulence. The dynamic interactions between these fungi and their hosts, coupled with the intricate genetic makeup, provide a deeper understanding of their behavior and impact. These insights are crucial for devising effective disease management strategies and safeguarding the health of diverse plant species. As the body of research grows, the intricate web of interactions between fungi, hosts, and the environment continues to unravel, offering valuable perspectives on combating plant diseases and ensuring sustainable agricultural practices. The ongoing development of NGS platforms and their integration with other emergent technologies hold the promise of enhancing our understanding of its biology and evolution [192].
7. Management Strategies for Neoscytalidium dimidiatum
7.1. Antifungal Activity of Different Applications
Neoscytalidium dimidiatum poses risks to various crops, notably dragon fruit, causing stem canker and fruit rot. Traditional chemical solutions, though effective, have drawbacks. This prompts researchers to explore alternative approaches to combat this fungal threat and reduce its impact on agriculture. This review section delves into recent studies examining N. dimidiatum’s susceptibility to interventions such as fungistatic pesticides, essential oils, biopesticides, biocontrol agents, and nanoparticles. This exploration aims to uncover optimal methods for tackling this fungal challenge and minimizing its effects on crop health.
7.1.1. Fungicides
Xu et al. [12] conducted groundbreaking research that highlighted the susceptibility of N. dimidiatum to a range of fungistatic pesticides, notably including hexaconazole, tebuconazole, flusiazole, and pyraclostrobin. Of particular significance is the potency demonstrated by pyraclostrobin EC at concentrations of 250 g/L, resulting in commendable control rates reaching up to 85%, comparable to the efficacy of azoxystrobin SC and tebuconazole. This insight was further corroborated by Xian et al. [203], who confirmed that field applications of pyraclostrobin EC at similar concentrations effectively reduced disease incidence to 85%, emphasizing the practical utility of this approach for stem canker control.
Concurrently, Noegrahati et al. [204] introduced azoxystrobin (200 g/L) and difenoconazole (124 g/L) as potent agents for managing stem canker in red dragon fruit. Their research underscores the safety of these agents in terms of residue, ensuring their viability for consumption. Parallelly, Eraslan Sür and Oksal [205] delved into the effects of fungicide concentrations on N. dimidiatum isolated from apricots. The synergistic combination of Floupyram (200 g/L) with Tebuconazole (200 g/L), along with Cyprodinil + Fludioxonil (37.5 + 25%) fungicides, demonstrated robust suppression of mycelial growth, accentuating their pivotal role in mitigating the expansion of this pathogenic fungus.
Fullerton et al. [45], in collaboration with the Southern Horticultural Research Institute (SOFRI), identified efficacious fungicides for canker control. This includes protectant fungicides like mancozeb and copper, as well as active constituents such as difenoconazole and azoxystrobin. These agents, with their translaminar and curative attributes, effectively permeate cladodes and counter the fungus post-infection. Fullerton et al. [45] underscored the importance of adaptive strategies in response to climate, recommending curative fungicides during wet seasons marked by frequent infections and protective fungicides timed with forecasted rainfall in dry seasons.
In parallel, the rigorous study conducted by Al Raish et al. [127] unveiled the vulnerability of royal poinciana trees in the UAE to stem canker disease caused by N. dimidiatum. This affliction encompasses symptoms such as desiccated branches, wilting leaves, bark lesions, xylem discoloration, wood necrosis, and gumming. Through rigorous investigation, diverse chemical fungicides, including Protifert®, Cidely® Top, and Amistrar® Top, were assessed. Significantly, Cidely® Top emerged as the optimal treatment against N. dimidiatum in field conditions, demonstrating its potential significance in managing the impact of this pathogen on royal poinciana trees.
Riska et al. [206] conducted in vitro experiments revealing that sodium salt, particularly when concentrations exceeded 3%, displayed substantial potential in hindering the mycelial growth of Neoscytalidium isolates. Furthermore, through a field trial, the application of a sodium salt solution at 30 g/L twice a week, combined with the rotation of chemical fungicides (propineb, 80% mancozeb, 50% carbendazim, and difenoconazole 250 g/L) once a week, effectively reduced the severity of stem canker disease on dragon fruit plants. The collective findings from culture and field investigations provide support for the feasibility of utilizing alternating chemical fungicides as a prospective strategy for managing stem canker disease on dragon fruit.
7.1.2. Essential Oils and Biopesticides
The efficacy of essential oils in addressing N. dimidiatum remains relatively uncharted. Campos-Rivero et al. [207] introduced a novel strategy involving active sachets encapsulating oregano oil with starch/agave fructans. These sachets, endowed with antifungal attributes, aim to combat postharvest losses caused by phytopathogens in fruits and vegetables. Through in vitro experiments, their study demonstrated the efficacy of oregano oil sachets in significantly suppressing the growth of N. hyalinum (now N. dimidiatum) at a temperature of 30 °C over a span of 12 days. This investigation marks the inaugural exploration of the potential of gaseous oregano oil to impede mycelial expansion in N. dimidiatum.
In addition, the study conducted by Taguiam et al. [208] demonstrates the inhibitory capacity of citronella oil, exhibiting a range of effects from 60% to 85.42% against N. dimidiatum. The same authors highlight a biopesticide formulation containing Bacillus subtilis, mancozeb, isoprothiolane, and citronella essential oil as a promising solution for mitigating N. dimidiatum. Nevertheless, given the partial inhibition of fungal growth by citronella oil (1.25 µL/mL), a thorough assessment of its potential phytotoxicity on dragon fruits is imperative before considering field applications. In Ratanaprom et al.’s [209] study, their experiments demonstrated that the combination of B. subtilis supernatant and sodium bicarbonate effectively inhibited the growth of N. dimidiatum through both direct and indirect mechanisms. The direct effect involves the secretion of antibiotic metabolites into the growing medium. Simultaneously, this treatment triggers the defense mechanisms of dragon fruit plants, leading to increased levels of PR proteins and enzymes associated with phenylpropanoid biosynthesis. This dual-action approach offers a robust defense against stem brown spot disease in dragon fruit plants.
Furthermore, Balendres et al. [13] demonstrated that the in vitro growth of N. dimidiatum can be effectively inhibited by a bio-fungicide formulation comprising B. subtilis, isoprothiolane, and mancozeb. This finding underscores the potential of the bio-fungicide in managing the growth and spread of N. dimidiatum. Such efficacy could prove invaluable for devising disease-management strategies in agricultural contexts, with potential implications for postharvest fruit handling practices and future investigations into the management of dragon fruit diseases.
7.1.3. Biocontrol Agents
Wang et al. [210] investigated the potential of Penicillium rolfsii (strain Y17), an endophytic fungus isolated from papaya leaves, as a control agent against N. dimidiatum-induced pitaya fruit canker disease. They systematically assessed the impact of Y17 treatment on disease progression, internal defense enzymes, and malondialdehyde content within the fruit. The results unveiled that Y17 treatment prompts the activation of antioxidant enzymes, thereby ameliorating oxidative stress and mitigating disease-associated damage. Added to that, this treatment fortifies the fruit’s immune responses and resistance to infection, offering promising avenues for advancing novel strategies in countering pitaya fruit canker disease.
Al Hamad et al. [211] examined the potential of actinobacterial isolates as biological control agents (BCAs) for managing stem canker disease on royal poinciana, attributed to N. dimidiatum. Their investigation highlights the inhibitory prowess of Streptomyces griseorubens UAE2 and Streptomyces wuyuanensis UAE1. These strains showcase robust antifungal effects through the production of antifungal compounds and cell-wall-degrading enzymes. Notably, S. griseorubens exhibits an added dimension by reinstating the activity of 1-aminocyclopropane-1-carboxylate deaminase (ACCD). Greenhouse and container nursery trials demonstrate the efficacy of S. griseorubens in curtailing disease symptoms, evidenced by reduced conidia numbers and defoliated leaves among royal poinciana seedlings. Their study underscores the superiority of S. griseorubens as a BCA, attributed not only to its antifungal attributes but also its ACCD secretion, contributing to effective stem canker disease management.
Lin et al. [212] focused on the efficacy of biocontrol strains for combatting pitaya canker. Their investigation centered on bacterial strain AF01, identified as Paenibacillus polymyxa. AF01 displays potent antifungal effects against N. dimidiatum and other pitaya fungal pathogens, attributed to the production of 13 fusaricidins that impair mycelial growth and spore germination by disrupting fungal cell membranes and ultrastructure. Pot experiments and yield evaluations validate AF01′s capacity to curtail the disease index. RNA-seq analysis provides insights into AF01′s targeted disruption of cellular processes, including cell wall biosynthesis and protein biosynthesis. These revelations suggest that P. polymyxa AF01 holds promise as a biocontrol agent for pitaya canker.
Ma et al. [213] identified bacterial strain P42 as Lysinibacillus boronitolerans, demonstrating robust antagonistic effects (82.18%) against soilborne pathogens. Strain P42 showcased broad-spectrum antagonistic activity encompassing N. dimidiatum and other tropical agricultural fungal pathogens.
In Travadon et al.’s [214] study, they assessed Trichoderma-based Biological Control Agents (BCAs) in a laboratory using detached almond stems. These evaluations aimed to determine the BCAs’ effectiveness against four canker pathogens, including N. dimidiatum. Their findings highlighted the remarkable success of two specific Trichoderma strains, Trichoderma atroviride SC1 and T. paratroviride RTFT014, in significantly reducing infections caused by all canker pathogens in almond trees.
7.1.4. Nanoparticles
Cu2O-Cu nanoparticles stabilized by alginate, developed by Du et al. [215], serve as a potential plant fungicide against N. dimidiatum-induced brown spot disease in dragon fruit plants. Cu2O-Cu nanoparticles stabilized in alginate exhibit noteworthy antifungal efficacy (~100%) against N. dimidiatum at a copper concentration of 30 ppm. This points to the substantial potential of Cu2O-Cu nanoparticles in alginate as promising nanomaterials for plant fungicide applications.
Tuan et al. [216] investigated the management of brown spot disease in dragon fruit plants caused by N. dimidiatum. They explored the efficacy of plant elicitors, oligochitosan (OC) and nanosilica (nSiO2), and prepared OC and nSiO2 synthesized from rice husk. The combined nSiO2 and OC treatment, referred to as nSiO2-OC, demonstrated efficacy in enhancing chitinase induction and reducing disease severity. Their study highlights the potential of utilizing the nSiO2-OC hybrid material as an innovative elicitor for brown spot disease management in dragon fruit plants, offering environmentally sustainable alternatives to hazardous agrochemicals.
Acay et al. [217] introduced a cost-effective method to synthesize a chitosan bionanocomposite using Morchella esculenta extract. The synthesized M. esculenta-chitosan bionanocomposite exhibited antifungal properties against various plant pathogens, including N. dimidiatum.
Hashem et al. [218] assessed the efficacy of a clove essential oil nanoemulsion (CEONE) as a control agent against N. dimidiatum affecting Carum carvi plants. CEONE exhibited substantial antifungal activity (25.5% to 82.2% inhibition) against N. dimidiatum. In pot experiments, CEONE and clove essential oil (CEO) mitigated N. dimidiatum blight disease, enhancing root length, plant height, and leaf development. Their study also explored CEONE and CEO’s potential to induce plant resistance through changes in biochemical markers.
Duong et al. [219] evaluated the antifungal efficacy of green-synthesized Cu/Cu2O nanocomposites against N. dimidiatum using in vitro assays. These nanocomposites exhibited significant antifungal potency against N. dimidiatum, with a minimum inhibitory concentration (MIC) of 0.0625 g/L and a measured inhibition zone diameter of 18.00 ± 0.58 mm. These findings highlight their potential utility as effective antifungal agents against N. dimidiatum.
7.2. Strategies for Control
In order to ensure the successful control of N. dimidiatum, it is imperative to give careful attention to the numerous biological and technical factors that contribute to its multidimensional nature. Central to effective disease mitigation is the comprehension and resolution of latent infections, where N. dimidiatum invades host plants and products inconspicuously. Sole reliance on visual inspection for latent infection detection proves inadequate, complicating pathogen identification and management [45,122,220]. In addition, the visual similarity between symptoms attributed to N. dimidiatum and those caused by other species within the Fusicoccum or Scytalidium genera, along with Botryosphaeriaceae fungi, hinders precise differentiation and risks misdiagnosis [45,122]. The absence of rapid molecular diagnostic tools further compounds the challenges, with in planta identification methods currently lacking practicality [45].
The vast host range of N. dimidiatum magnifies the complexity of control efforts, necessitating tailored identification and management strategies due to the heterogeneous susceptibility across species [122]. Amidst these challenges, investing in comprehensive research and development emerges as paramount. The establishment of swift and reliable diagnostic methodologies rooted in molecular techniques stands to greatly enhance early pathogen detection [45]. Advancing our understanding of N. dimidiatum’s biology, host interactions, and disease progression holds promise for formulating precise control strategies tailored to specific host plants [45,122].
Fullerton et al. [45] emphasize the significance of rigorous orchard hygiene as a fundamental principle for the successful management of canker disease. Vital to successful control is the elimination of spore sources and the prompt removal of infected cladodes, while the time lag between symptom manifestation and sporulation offers an opportunity for pre-emptive excision of nascent lesions. Combining robust orchard hygiene with judicious fungicide application proves instrumental in expeditiously reducing disease burden, facilitating orchard recovery within a matter of months.
Brito et al. [122] accentuated the centrality of stem-cutting dry rot in the survival and dissemination of pathogenic Botryosphaeriaceae within cassava. Effective management strategies for black root rot and stem-cutting dry rot necessitate the utilization of high-quality propagating material and precise pretreatment protocols. Sakalidis et al. [83] identified N. novaehollandiae and N. dimidiatum among various Botryosphaeriaceae taxa with diverse roles as canker-associated fungi, endophytes, or potential pathogens. The unique growth conditions of the Kimberley region hinder pathogen establishment, while mechanical pruning wounds and excessive pruning contribute to plant stress.
In light of N. dimidiatum’s primary entry through wounds, especially those incurred during mechanical pruning, the mitigation of such wounds through careful practices or, where possible, avoiding pruning altogether becomes pivotal [76,165]. The application of fungicides or the introduction of effective biological control agents targeted at wound sites, such as pruning and grafting wounds, emerges as a promising preventive measure [76,138]. Targeted fungicide application during wet periods when infection conditions are conducive may further bolster control efforts [45]. Exploration of host cultivars exhibiting resistance or reduced susceptibility to N. dimidiatum canker provides valuable insight for breeding programs aimed at developing resistant varieties [122]. Ray et al. [35] and Esmaeili et al. [98] emphasized the significance of incorporating susceptible host functions into strategies for managing trunk diseases.
Owing to the need to minimize inoculum sources, orchard hygiene continues to be a pillar, necessitating the careful elimination and appropriate disposal of infected plant debris [45]. Given N. dimidiatum’s affinity for warmer temperatures, careful management of orchard temperature conditions, including irrigation and shading practices, warrants consideration [32]. By implementing rigorous protocols, conducting thorough inspections, and collaborating with experts, nurseries can significantly reduce the risk of propagating this pathogen and other latent infections in agricultural and horticultural settings [46]. The careful selection of seeds from reliable sources is also crucial in preventing the spread of N. dimidiatum [47,49,107].
A holistic approach to control involves vigilant monitoring for early signs of infection and employing rapid molecular-based diagnostic methods [45,48]. Research into the potential vectors, such as seeds and arthropods, aids in understanding disease dissemination [39,49,107,190]. Ongoing research into N. dimidiatum’s behavior, interactions, and disease progression contribute critical insights to developing refined control strategies [33,41,45].
With these research insights, investing in comprehensive research and development assumes pivotal importance for effective N. dimidiatum control. Molecular-based diagnostics and an enhanced understanding of pathogen–host dynamics offer the potential for targeted control strategies aligned with specific host plants. Integrating these strategies into disease management protocols promises to bolster crop yields and secure the sustainability of agricultural practices.
8. Future Challenges
In light of the comprehensive investigations into N. dimidiatum’s taxonomy, host range, biology, virulence traits, and management strategies, several intriguing avenues for future research emerge. While some progress has been made in understanding N. dimidiatum’s genetic makeup, there remains a need to unravel the species’ specific genetic traits linked to its adaptation to diverse environments. Uncovering the genetic mechanisms that allow N. dimidiatum to thrive across various hosts and ecological niches could shed light on its remarkable adaptability. Elucidating the intricate dynamics between N. dimidiatum and its host plants holds considerable potential. Exploring how the pathogen interacts with plants at the molecular, cellular, and physiological levels could unveil the underlying mechanisms driving infection, colonization, and disease development. Given the potential impact of climate change on N. dimidiatum, future research should delve into how shifting climatic conditions might influence the distribution, prevalence, and severity of N. dimidiatum infections. Understanding these dynamics can aid in developing targeted management strategies. As the management of N. dimidiatum and related pathogens remains a challenge, exploring novel, ecofriendly approaches could prove fruitful. Investigating the potential of biological control agents, as well as eco-friendly chemicals, could offer sustainable alternatives for disease management. Understanding how human-related factors, such as urbanization and agricultural practices, may influence N. dimidiatum’s ecology could provide insights into effective management strategies. Investigating any potential shifts in the pathogenicity or virulence of N. dimidiatum strains is essential, especially in light of changing environmental conditions and potential interactions with other pathogens. This research can enhance our ability to predict and mitigate disease outbreaks.
In conclusion, the intricate web of interactions involving N. dimidiatum necessitates continued research to unravel its genetic, ecological, and pathogenic intricacies. Addressing these future challenges will not only enhance our understanding of this enigmatic pathogen but also contribute to the development of effective strategies for disease management and mitigation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9111048/s1, Table S1: Collection details and GenBank accession numbers of isolates included in the phylogenetic analyses.
Author Contributions
S.D. and G.Ö., writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data generated or analyzed during this study are included in this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kazemzadeh, C.M.; Mohammadi, H.; Khodaparast, S.A. Diversity and pathogenicity of Botryosphaeriaceae species on forest trees in the north of Iran. Eur. J. For. Res. 2019, 138, 685–704. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef] [PubMed]
- Heidari, S.; Gheisari, M.; Abastabar, M.; Pourabdollah, M.; Mirenayat, M.S.; Basharzad, N.; Seifi, S.; Tavakoli, M.; Jafarzadeh, J.; Ansari, S.; et al. Genotyping and ın vitro antifungal susceptibility profile of Neoscytalidium species isolates from respiratory tract. Mycopathologia 2021, 186, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadi, S.; Nasirharandi, S.; Pourahmad, M.; Ahmadikia, K.; Mokhtari, M.; Najafzadeh, M.J.; Mohammadi, R. Phaeohyphomycosis caused by Neoscytalidium dimidiatum in a COVID-19 patient. Rev. Iberoam. Micol. 2022, 39, 72–76. [Google Scholar] [CrossRef]
- Chuang, M.F.; Ni, H.F.; Yang, H.R.; Shu, S.L.; Lai, S.Y.; Jiang, Y.L. First report of stem canker disease of pitaya (Hylocereus undatus and H. polyrhizus) caused by Neoscytalidium dimidiatum in Taiwan. Plant Dis. 2012, 96, 906. [Google Scholar] [CrossRef]
- Lan, G.B.; He, Z.F.; Xi, P.G.; Jiang, Z.D. First report of brown spot disease caused by Neoscytalidium dimidiatum on Hylocereus undatus in Guangdong, Chinese Mainland. Plant Dis. 2012, 96, 1702. [Google Scholar] [CrossRef]
- Yi, R.H.; Gan, L.J.; Yan, D.H.; Wu, Z.J.; Tong, Y.T.; Wu, F.F. Identification and biological characteristics of Neoscytalidium dimidiatum causing pitaya canker disease. Acta Phytophylacica Sin. 2013, 40, 102–108. [Google Scholar] [CrossRef]
- Ezra, D.; Liarzi, O.; Gat, T.; Hershcovich, M.; Dudai, M. First report of internal black rot caused by Neoscytalidium dimidiatum on Hylocereus undatus (pitahaya) fruit in Israel. Plant Dis. 2013, 97, 1513. [Google Scholar] [CrossRef]
- Mohd, M.H.; Salleh, B.; Zakaria, L. Identification and molecular characterizations of Neoscytalidium dimidiatum causing stem canker of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopathol. 2013, 161, 841–849. [Google Scholar] [CrossRef]
- Sanahuja, G.; Lopez, P.; Palmateer, A.J. First report of Neoscytalidium dimidiatum causing stem and fruit canker of Hylocereus undatus in Florida. Plant Dis. 2016, 100, 1499. [Google Scholar] [CrossRef]
- Hong, C.F.; Gazis, R.; Crane, J.H.; Zhang, S. Prevalence and epidemics of Neoscytalidium stem and fruit canker on pitahaya (Hylocereus spp.) in South Florida. Plant Dis. 2020, 104, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Peng, Y.; Qi, Z.; Yan, Z.; Yang, L.; He, M.D.; Li, Q.X.; Liu, C.L.; Ruan, Y.Z.; Wei, S.S.; et al. Identification of Neoscytalidium dimidiatum causing canker disease of pitaya in Hainan, China. Australas. Plant Pathol. 2018, 47, 547–553. [Google Scholar] [CrossRef]
- Balendres, M.A.; Taguiam, J.D.; Evallo, E.; Estigoy, J.; Cortaga, C. Fruit Brown Rot Caused by Neoscytalidium dimidiatum on Selenicereus monacanthus in the Philippines; MycoAsia: Visakhapatnam, India, 2022. [Google Scholar] [CrossRef]
- Serrato-Diaz, L.M.; Goenaga, R. First report of Neoscytalidium dimidiatum causing stem canker on dragon fruit (Hylocereus spp.) in Puerto Rico. Plant Dis. 2021, 105, 2728. [Google Scholar] [CrossRef] [PubMed]
- Dy, K.S.; Wonglom, P.; Pornsuriya, C.; Sunpapao, A. Morphological, molecular ıdentification and pathogenicity of Neoscytalidium dimidiatum causing stem canker of Hylocereus polyrhizus in Southern Thailand. Plants 2022, 11, 504. [Google Scholar] [CrossRef]
- Salunkhe, V.N.; Bhagat, Y.S.; Chavan, S.B.; Lonkar, S.G.; Kakade, V.D. First report of Neoscytalidium dimidiatum causing stem canker of dragon fruit (Hylocereus spp.) in India. Plant Dis. 2023, 107, 1222. [Google Scholar] [CrossRef]
- Khoo, Y.W.; Tan, H.T.; Khaw, Y.S.; Li, S.F.; Chong, K.P. First report of Neoscytalidium dimidiatum causing stem canker on Selenicereus megalanthus in Malaysia. Plant Dis. 2023, 107, 222. [Google Scholar] [CrossRef]
- Espinoza-Lozano, L.; Sumba, M.; Calero, A.; Jiménez, M.I.; Quito-Avila, D.F. First report of Neoscytalidium dimidiatum causing stem canker on yellow dragon fruit (Hylocereus megalantus) in Ecuador. Plant Dis. 2023, 107, 1949. [Google Scholar] [CrossRef]
- Polizzi, G.; Aiello, D.; Vitale, A.; Giuffrida, F.; Groenewald, J.Z.; Crous, P.W. First Report of shoot blight, canker, and gummosis caused by Neoscytalidium dimidiatum on citrus in Italy. Plant Dis. 2009, 93, 1215. [Google Scholar] [CrossRef]
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, distribution, and pathogenicity of Diatrypaceae and Botryosphaeriaceae associated with citrus branch canker in the Southern California desert. Plant Dis. 2016, 100, 2402–2413. [Google Scholar] [CrossRef]
- Al-Saadoon, A.H.; Ameen, M.K.M.; Hameed, M.A.; Al-Badran, A.; Ali, Z. First report of grapevine dieback caused by Lasiodiplodia theobromae and Neoscytalidium dimidiatum in Basrah, Southern Iraq. Afr. J. Biotechnol. 2012, 11, 16165–16171. [Google Scholar] [CrossRef]
- Rolshausen, P.E.; Akgul, D.S.; Perez, R.; Eskalen, A.; Gispert, C. First report of wood canker caused by Neoscytalidium dimidiatum on grapevine in California. Plant Dis. 2013, 97, 1511. [Google Scholar] [CrossRef] [PubMed]
- Correia, K.C.; Silva, M.A.; Netto, M.S.B.; Vieira, W.A.S.; Camara, M.P.S.; Michereff, S.J. First report of grapevine dieback caused by Neoscytalidium hyalinum in Brazil. Plant Dis. 2016, 100, 213. [Google Scholar] [CrossRef]
- Oksal, E.; Çelik, Y.; Özer, G. Neoscytalidium dimidiatum causes canker and dieback on grapevine in Turkey. Australas. Plant Dis. Notes 2019, 14, 33. [Google Scholar] [CrossRef]
- Akgül, D.S.; Savaş, N.G.; Özarslandan, M. First report of wood canker caused by Lasiodiplodia exigua and Neoscytalidium novaehollandiae on grapevine in Turkey. Plant Dis. 2019, 103, 1036–1037. [Google Scholar] [CrossRef]
- Bahmani, Z.; Abdollahzadeh, J.; Amini, J.; Evidente, A. Biscogniauxia rosacearum the charcoal canker agent as a pathogen associated with grapevine trunk diseases in Zagros region of Iran. Sci. Rep. 2021, 11, 14098. [Google Scholar] [CrossRef] [PubMed]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing shoot and needle blight of pines (Pinus spp.) in Turkey. Plant Dis. 2019, 103, 2960–2961. [Google Scholar] [CrossRef]
- Alizadeh, M.; Safaie, N.; Shams-Bakhsh, M.; Mehrabadi, M. Neoscytalidium novaehollandiae causes dieback on Pinus eldarica and its potential for infection of urban forest trees. Sci. Rep. 2022, 12, 9337. [Google Scholar] [CrossRef]
- Hajlaoui, M.R.; Nouri, M.T.; Hamrouni, N.; Trouillas, F.P.; Ben Yahmed, N.; Eddouzi, J.; Mnari-Hattab, M. First record of dieback and decline of plum caused by Neoscytalidium dimidiatum in Tunisia. New Dis. Rep. 2018, 38, 20. [Google Scholar] [CrossRef]
- Nouri, M.T.; Lawrence, D.P.; Yaghmour, M.A.; Michailides, T.J.; Trouillas, F.P. Neoscytalidium dimidiatum causing canker, shoot blight and fruit rot of almond in California. Plant Dis. 2018, 102, 1638–1647. [Google Scholar] [CrossRef]
- Oksal, E.; Yiğit, T.; Özer, G. First report of Neoscytalidium dimidiatum causing shoot blight, dieback and canker of apricot in Turkey. J. Plant Pathol. 2020, 102, 579–580. [Google Scholar] [CrossRef]
- Ören, E.; Koca, G.; Gencer, R.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with stem canker and branch dieback of almond trees. Australas. Plant Dis. Notes 2020, 15, 17. [Google Scholar] [CrossRef]
- Holland, L.A.; Trouillas, F.P.; Nouri, M.T.; Lawrence, D.P.; Crespo, M.; Doll, D.A.; Duncan, R.A.; Holtz, B.A.; Culumber, C.M.; Yaghmour, M.A.; et al. Fungal pathogens associated with canker diseases of almond in California. Plant Dis. 2021, 105, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, A.E.; Ba-Omar, T. First report of Albizia lebbeck dieback caused by Scytalidium dimidiatum in Oman. Mycopathologia 2002, 154, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.D.; Burgess, T.; Lanoiselet, V.M. First record of Neoscytalidium dimidiatum and N. novaehollandiae on Mangifera indica and N. dimidiatum on Ficus carica in Australia. Australas. Plant Dis. Notes 2010, 5, 48–50. [Google Scholar] [CrossRef]
- Fernández-Herrera, E.; Moreno-Salazar, S.F.; Rentería-Martínez, M.E.; Arratia-Castro, A.A.; Villar-Luna, E. Neoscytalidium dimidiatum: Causal agent of dieback in Ficus benjamina L. in Mexico. Rev. Chapingo Ser. Hortic. 2017, 23, 203–210. [Google Scholar] [CrossRef]
- Al-Bedak, O.A.; Mohamed, R.A.; Seddek, N.H. First detection of Neoscytalidium dimidiatum associated with canker disease in Egyptian Ficus trees. For. Pathol. 2018, 48, e12411. [Google Scholar] [CrossRef]
- Güney, İ.G.; Bozoğlu, T.; Özer, G.; Türkölmez, Ş.; Derviş, S. First report of Neoscytalidium dimidiatum associated with dieback and canker of common fig (Ficus carica L.) in Turkey. J. Plant Dis. Prot. 2022, 129, 701–705. [Google Scholar] [CrossRef]
- Yeganeh, S.; Mohammadi, H. Sooty canker, a destructive disease of banyan (Ficus benghalensis L.) trees in landscapes of Kish Island (Iran). Urban For. Urban Green. 2022, 72, 127573. [Google Scholar] [CrossRef]
- Ahmadpour, S.A.; Mehrabi-Koushki, M.; Farokhinejad, R.; Mirsoleymani, Z. Characterization and pathogenicity of Neoscytalidium novaehollandiae causing dieback and sooty canker in Iran. Trop. Plant Pathol. 2023. [Google Scholar] [CrossRef]
- Gusella, G.; Fiore, G.; Vitale, A.; Felts, D.G.; Michailides, T.J. New findings on the effects of different factors involved in fig limb dieback caused by Neoscytalidium dimidiatum in California. Eur. J. Plant Pathol. 2023, 167, 89–97. [Google Scholar] [CrossRef]
- Derviş, S.; Türkölmez, Ş.; Çiftçi, O.; Ulubaş Serçe, Ç.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing canker, shoot blight, and root rot of pistachio in Turkey. Plant Dis. 2019, 103, 1411. [Google Scholar] [CrossRef]
- Hashemi, H.; Mohammadi, H. Identification and characterization of fungi associated with internal wood lesions and decline disease of willow and poplar trees in Iran. For. Pathol. 2016, 46, 341–352. [Google Scholar] [CrossRef]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Ulubaş Serçe, Ç.; Türkölmez, C.G.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing dieback, shoot blight, and branch canker of willow trees in Turkey. Plant Dis. 2019, 103, 2139. [Google Scholar] [CrossRef]
- Fullerton, R.A.; Sutherland, P.A.; Rebstock, R.S.; Hieu, N.T.; Thu, N.N.A.; Linh, D.T.; Thanh, N.T.K.; Van Hoa, N. The life cycle of dragon fruit canker caused by Neoscytalidium dimidiatum and implications for control. In Proceedings of the Dragon Fruit Regional Network Initiation Workshop, Taipei, Taiwan, 23–24 April 2018; pp. 71–80. [Google Scholar]
- Al-Sadi, A.M.; Al-Ghaithi, A.G.; Al-Fahdi, N.; Al-Yahyai, R. Characterization and pathogenicity of fungal pathogens associated with root diseases of citrus in Oman. Int. J. Agric. Biol. 2014, 16, 371–376. [Google Scholar]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Serçe, Ç.U.; Dikilitas, M. New disease caused by Neoscytalidium dimidiatum devastates tomatoes (Solanum lycopersicum) in Turkey. Crop Prot. 2019, 118, 21–30. [Google Scholar] [CrossRef]
- Jiménez Luna, I.; Doll, D.; Ashworth, V.; Trouillas, F.P.; Rolshausen, P.E. Comparative profiling of wood canker pathogens from spore traps and symptomatic plant samples within California almond and walnut orchards. Plant Dis. 2022, 106, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Güney, İ.G.; Bozoğlu, T.; Özer, G.; Derviş, S. A novel blight and root rot of chickpea: A new host for Neoscytalidium dimidiatum. Crop Prot. 2023, 172, 106326. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.; Philips, A.J.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef]
- Pavlic, D.; Wingfield, M.J.; Barber, P.; Slippers, B.; Hardy, G.E.; Burgess, T.I. Seven new species of the Botryosphaeriaceae from baobab and other native trees in Western Australia. Mycologia 2008, 100, 851–866. [Google Scholar] [CrossRef]
- Phillips, A.J.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Huang, S.K.; Tangthirasunun, N.; Phillips, A.J.; Dai, D.Q.; Wanasinghe, D.N.; Wen, T.C.; Bahkali, A.H.; Hyde, K.D.; Kang, J.C. Morphology and phylogeny of Neoscytalidium orchidacearum sp. nov. (Botryosphaeriaceae). Mycobiology 2016, 44, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Calvillo-Medina, R.P.; Martinez-Neria, M.; Mena-Portales, J.; Barba-Escoto, L.; Raymundo, T.; Campos-Guillen, J.; Jones, G.H.; Reyes-Grajeda, J.P.; Gonzalez, Y.M.J.A.; Bautista-de Lucio, V.M. Identification and biofilm development by a new fungal keratitis aetiologic agent. Mycoses 2019, 62, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Pornsuriya, C.; Sunpapao, A. A new species of Neoscytalidium hylocereum sp. nov. causing canker on red-fleshed dragon fruit (Hylocereus polyrhizus) in Southern Thailand. J. Fungi 2023, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Nattrass, R.M. A new species of Hendersonula (H. toruloidea) on deciduous trees in Egypt. Trans. Br. Mycol. Soc. 1933, 18, 189–198. [Google Scholar] [CrossRef]
- Sutton, B.C.; Dyko, B.J. Revision of Hendersonula. Mycol. Res. 1989, 93, 466–488. [Google Scholar] [CrossRef]
- Farr, D.F.; Elliott, M.; Rossman, A.Y.; Edmonds, R.L. Fusicoccum arbuti sp. nov. causing cankers on Pacific madrone in western North America with notes on Fusicoccum dimidiatum, the correct name for Scytalidium dimidiatum and Natrassia mangiferae. Mycologia 2005, 97, 730–741. [Google Scholar] [CrossRef]
- Penzig, O. Funghi agrumicoli. Contribuzione allo studio dei funghi parassiti degli agrumi. Michelia 1882, 2, 385–508. [Google Scholar]
- Wilson, E.E. The branch wilt of Persian walnut trees and its cause. Hilgardia 1947, 17, 413–436. [Google Scholar] [CrossRef]
- Campbell, C.K.; Mulder, J.L. Skin and nail infection by Scytalidium hyalinum sp. nov. Sabouraudia 1977, 15, 161–166. [Google Scholar] [CrossRef]
- Slippers, B.; Johnson, G.I.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Phylogenetic and morphological re-evaluation of the Botryosphaeria species causing diseases of Mangifera indica. Mycologia 2005, 97, 99–110. [Google Scholar] [CrossRef]
- Sydow, H.; Sydow, P.; Butler, E.J. Fungi Indiae Orientalis. V. Ann. Mycol. 1916, 14, 177–220. [Google Scholar]
- Madrid, H.; Ruiz-Cendoya, M.; Cano, J.; Stchigel, A.; Orofino, R.; Guarro, J. Genotyping and in vitro antifungal susceptibility of Neoscytalidium dimidiatum isolates from different origins. Int. J. Antimicrob. Agents 2009, 34, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Fungal species concepts in the genomics era. Genome 2020, 63, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Maharachchikumbura, S.S.N.; Chen, Y.; Ariyawansa, H.A.; Hyde, K.D.; Haelewaters, D.; Perera, R.H.; Samarakoon, M.C.; Wanasinghe, D.N.; Bustamante, D.E.; Liu, J.; et al. Integrative approaches for species delimitation in Ascomycota. Fungal Divers. 2021, 109, 155–179. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony. (* and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Coutinho, I.B.L.; Cardoso, J.E.; Lima, C.S.; Lima, J.S.; Goncalves, F.J.T.; Silva, A.M.S.; Freire, F.C.O. An emended description of Neofusicoccum brasiliense and characterization of Neoscytalidium and Pseudofusicoccum species associated with tropical fruit plants in northeastern Brazil. Phytotaxa 2018, 358, 251–264. [Google Scholar] [CrossRef]
- Hohenfeld, C.S.; Santana, M.P.; Junior, L.R.C.; de Oliveira, E.J.; de Oliveira, S.A.S. Modelling growth characteristics and aggressiveness of Neoscytalidium hyalinum and Fusarium solani associated with black and dry root rot diseases on cassava. Trop. Plant Pathol. 2018, 43, 422–432. [Google Scholar] [CrossRef]
- Sabernasab, M.; Jamali, S.; Marefat, A.; Abbasi, S. Morphological and molecular characterization of Neoscytalidium novaehollandiae, the cause of Quercus brantii dieback in Iran. Phytopathol. Mediterr. 2019, 58, 347–357. [Google Scholar] [CrossRef]
- Gusella, G.; Morgan, D.P.; Michailides, T.J. Further ınvestigation on limb dieback of fig (Ficus carica) Caused by Neoscytalidium dimidiatum in California. Plant Dis. 2021, 105, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Güney, İ.G.; Özer, G.; Türkölmez, Ş.; Derviş, S. Canker and leaf scorch on olive (Olea europaea L.) caused by Neoscytalidium dimidiatum in Turkey. Crop Prot. 2022, 157, 105985. [Google Scholar] [CrossRef]
- Goudarzi, A.; Moslehi, M. Distribution of a devastating fungal pathogen in mangrove forests of southern Iran. Crop Prot. 2020, 128, 104987. [Google Scholar] [CrossRef]
- Nurul Nadiah, M.A.; Mohamed Nor, N.M.I.; Latiffah, Z.; Masratul Hawa, M. First report of leaf blight on white spider lily caused by Neoscytalidium dimidiatum in Malaysia. New Dis. Rep. 2017, 35, 16. [Google Scholar] [CrossRef]
- Zaeimian, Z.; Fotouhifar, K.B. First report of Neoscytalidium dimidiatum as the causal agent of leaf blight on Clivia miniata. Sci. Rep. 2023, 13, 16110. [Google Scholar] [CrossRef]
- Pandey, R.S.; Bhargava, S.N.; Shukla, D.N.; Khati, D.V.S. A new leaf spot disease of mango. Plant Dis. 1981, 65, 441–442. [Google Scholar] [CrossRef]
- Reckhaus, P.; Adamous, I. Hendersonula dieback of mango in Niger. Plant Dis. 1987, 71, 1045. [Google Scholar] [CrossRef]
- Mendes, M.A.S.; da Silva, V.L.; Dianese, J.C.; Ferreira, M.A.S.V.; dos Santos, C.E.N.; Urben, A.F.; Castro, C.; Gomes Neto, E. Fungos em Plants no Brasil; Embrapa-SPI/Embrapa-Cenargen: Brasilia, Brazil, 1998; 555p. [Google Scholar]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; St, J.; Hardy, G.E.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. Eur. J. Plant Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef]
- Marques, M.W.; Lima, N.B.; de Morais, M.A.; Michereff, S.J.; Phillips, A.J.; Câmara, M.P. Botryosphaeria, Neofusicoccum, Neoscytalidium and Pseudofusicoccum species associated with mango in Brazil. Fungal Divers. 2013, 61, 195–208. [Google Scholar] [CrossRef]
- Dolatabad, H.K.; Javan-Nikkhah, M.; Shier, W.T. Evaluation of antifungal, phosphate solubilisation, and siderophore and chitinase release activities of endophytic fungi from Pistacia vera. Mycol. Prog. 2017, 16, 777–790. [Google Scholar] [CrossRef]
- Kurt, Ş.; Uysal, A.; Soylu, E.M.; Kara, M.; Soylu, S. First record of Neoscytalidium novaehollandiae associated with pistachio dieback in the Southeastern Anatolia region of Turkey. Mycol. Iran 2019, 6, 55–57. [Google Scholar] [CrossRef]
- Williams, L.; Hayne, S.C. Index of Plant Diseases in West Virginia; West Virginia Department of Agriculture Plant Industries Division: Ripley, WI, USA, 1982; 115p. [Google Scholar]
- Farr, D.F.; Bills, G.F.; Chamuris, G.P.; Rossman, A.Y. Fungi on Plants and Plant Products in the United States; The American Phytopathological Society: St. Paul, MN, USA, 1989. [Google Scholar]
- Mathur, R.S. The Coelomycetes of India; Bishen Singh Mahendra Pal Singh: Delhi, India, 1979; 460p. [Google Scholar]
- Gusella, G.; Di Pietro, C.; Vecchio, L.; Campo, G.; Polizzi, G. Branch canker and dieback of Meryta denhamii caused by Neofusicoccum parvum and Neoscytalidium dimidiatum in Italy. Australas. Plant Dis. Notes 2023, 18, 31. [Google Scholar] [CrossRef]
- Peregrine, W.T.H.; Ahmad, K.B. Brunei: A first annotated list of plant diseases and associated organisms. Phytopathol. Pap. 1982, 27, 1–87. [Google Scholar]
- Kranz, J. Fungi collected in the Republic of Guinea, Collections from the Kindia area in 1962. Sydowia 1963, 17, 174–185. [Google Scholar]
- Xie, H.H.; Long, L.; Huang, S.; Mao, L.; Huang, Q.; Wang, L.; Li, J. First Report of Black Spot Caused by Neoscytalidium dimidiatum on Sisal in Guangxi, China. Plant Dis. 2021, 105, 701. [Google Scholar] [CrossRef]
- Johnston, A. A supplement to a host list of plant diseases in Malaya. Mycol. Pap. 1960, 77, 1–30. [Google Scholar]
- Kee, Y.J.; Suhaimi, N.N.; Zakaria, L.; Mohd, M.H. Characterisation of Neoscytalidium dimidiatum causing leaf blight on Sansevieria trifasciata in Malaysia. Australas. Plant Dis. Notes 2017, 12, 60. [Google Scholar] [CrossRef]
- Monteles, R.P.; Sousa, E.S.; da Silva Matos, K.; de Brito, V.S.T.; de Melo, M.P.; Beserra, J.E.A. Neoscytalidium dimidiatum causes leaf blight on Sansevieria trifasciata in Brazil. Australas. Plant Dis. Notes 2020, 15, 19. [Google Scholar] [CrossRef]
- Crous, P.W.; Hernández-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Kölling, G.; Marais, E.; Groenewald, J.Z. New and interesting fungi. Fungal Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef]
- Esmaeili, N.; Mohammadi, H.; Sohrabi, M. Barberry (Berberis vulgaris L.) as an alternative host of grapevine fungal trunk pathogens. Eur. J. Plant Pathol. 2023. [Google Scholar] [CrossRef]
- Ebbels, D.L.; Allen, D.J. A supplementary and annotated list of plant diseases, pathogens and associated fungi in Tanzania. Phytopathol. Pap. 1979, 22, 1–89. [Google Scholar]
- Kuruppu, M.; Siddiqui, Y.; Ahmad, K.; Ali, A. First report of postharvest stem end rot disease on MD2 pineapple fruits caused by Neoscytalidium dimidiatum in Malaysia. Plant Dis. 2021, 105, 1564. [Google Scholar] [CrossRef] [PubMed]
- Feijo, F.M.; Silva, M.J.S.; Nascimento, A.D.; Infante, N.B.; Ramos-Sobrinho, R.; Assuncao, I.P.; Lima, G.S.A. Botryosphaeriaceae species associated with the pickly pear cactus, Nopalea cochenillifera. Trop. Plant Pathol. 2019, 44, 452–459. [Google Scholar] [CrossRef]
- Yi, R.H.; Lin, Q.L.; Mo, J.J.; Wu, F.F.; Chen, J. Fruit internal brown rot caused by Neoscytalidium dimidiatum on pitahaya in Guangdong Province, China. Australas. Plant Dis. Notes 2015, 10, 13. [Google Scholar] [CrossRef]
- Boa, E.; Lenné, J. Diseases of Nitrogen Fixing Trees in Developing Countries. An Annotated List; Natural Resources Institute: Kent, UK, 1994; 82p. [Google Scholar]
- Watson, A.J. Foreign Bacterial and Fungus Diseases of Food, Forage, and Fiber Crops; U.S.D.A. Agricultural Research Service: Washington, DC, USA, 1971; Volume 111.
- de Mello, J.F.; Brito, A.C.Q.; Motta, C.M.S.; Vieira, J.C.B.; Michereff, S.J.; Machado, A.R. First report of Neoscytalidium dimidiatum causing root rot in sweet potato in Brazil. Plant Dis. 2019, 103, 373. [Google Scholar] [CrossRef]
- de Mello, J.F.; Brito, A.C.Q.; Vieira, J.C.B.; Camara, M.P.S.; Michereff, S.J.; de Souza-Motta, C.M.; Machado, A.R. Identification and pathogenicity of Botryosphaeriaceae species associated with root and stem rot of sweet potato in Brazil. Plant Pathol. 2021, 70, 1601–1615. [Google Scholar] [CrossRef]
- Mirtalebi, M.; Sabahi, F.; Banihashemi, Z. Fruit rot caused by Neoscytalidium hyalinum on melon in Iran. Australas. Plant Dis. Notes 2019, 14, 8. [Google Scholar] [CrossRef]
- Anonymous. California Fungi. Nos. 1-1325. Exsiccati Set 1931–1970, N/A Pages. Available online: https://data.nal.usda.gov/dataset/united-states-national-fungus-collections-fungus-host-dataset (accessed on 22 August 2023).
- Lin, C.H.; Chen, Y.X.; Liu, W.B.; Wu, W.Q.; Miao, W.G.; Zheng, F.C. First report of Dioscorea esculenta dieback caused by Neoscytalidium dimidiatum in China. Plant Dis. 2017, 101, 1320. [Google Scholar] [CrossRef]
- Arrieta-Guerra, J.J.; Díaz-Cabadiaz, A.T.; Pérez-Pazos, J.V.; Cadena-Torres, J.; Sánchez-López, D.B. Fungi associated with dry rot disease of yam (Dioscorea rotundata Poir.) tubers in Cordoba, Colombia. Agron. Mesoam. 2021, 32, 790–807. [Google Scholar] [CrossRef]
- Ören, E.; Koca, G.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with branch dieback on Japanese persimmon in Turkey. J. Plant Pathol. 2020, 102, 1311–1312. [Google Scholar] [CrossRef]
- Davison, A.D. Factors affecting development of madrone canker. Plant Dis. Rep. 1972, 56, 50–52. [Google Scholar]
- Shaw, C.G. Host Fungus Index for the Pacific Northwest-I. Hosts; Washington Agricultural Experiment Station: Washington, DC, USA, 1973; Volume 765, pp. 1–121. [Google Scholar]
- Ginns, J.H. Compendium of plant disease and decay fungi in Canada 1960–1980. Res. Br. Can. Agric. Publ. 1986, 1813, 416. [Google Scholar]
- French, A.M. California Plant Disease Host Index; California Department of Food and Agriculture: Sacramento, CA, USA, 1989; 394p.
- Tsahouridou, P.C.; Thanassoulopoulos, C.C. First report of Hendersonula toruloidea as a foliar pathogen of strawberry-tree (Arbutus unedo) in Europe. Plant Dis. 2000, 84, 487. [Google Scholar] [CrossRef]
- Machado, A.R.; Pinho, D.B.; Dutra, D.C.; Pereira, O.L. First report of collar and root rot of physic nut (Jatropha curcas) caused by Neoscytalidium dimidiatum in Brazil. Plant Dis. 2012, 96, 1697. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.R.; Pinho, D.B.; Pereira, O.L. Phylogeny, identification and pathogenicity of the Botryosphaeriaceae associated with collar and root rot of the biofuel plant Jatropha curcas in Brazil, with a description of new species of Lasiodiplodia. Fungal Divers. 2014, 67, 231–247. [Google Scholar] [CrossRef]
- Hughes, S.J. Fungi From the Gold Coast. I. Mycol. Pap. 1952, 48, 1–91. [Google Scholar]
- Nattrass, R.M. Host lists of Kenya fungi and bacteria. Mycol. Pap. 1961, 81, 1–46. [Google Scholar]
- Machado, A.R.; Pinho, D.B.; Oliveira, S.A.S.; Pereira, O.L. New occurrences of Botryosphaeriaceae causing black root rot of cassava in Brazil. Trop. Plant Pathol. 2014, 39, 464–470. [Google Scholar] [CrossRef]
- Brito, A.C.Q.; de Mello, J.F.; Camara, M.P.S.; Vieira, J.C.B.; Michereff, S.J.; Souza-Motta, C.M.; Machado, A.R. Diversity and pathogenicity of Botryosphaeriaceae species associated with black root rot and stem cutting dry rot in Manihot esculenta in Brazil. Eur. J. Plant Pathol. 2020, 157, 583–598. [Google Scholar] [CrossRef]
- Sangpueak, R.; Duchanee, S.; Saengchan, C.; Papathoti, N.K.; Hoang, N.H.; Thanh, T.L.; Phansak, P.; Buensanteai, N. Identification of cassava black stem and root rot agents in Thailand. Chil. J. Agric. Res. 2023, 83, 70–82. [Google Scholar] [CrossRef]
- Pande, A.; Rao, V.G. A Compendium Fungi on Legumes from India; Scientific Publishers: Jodhpur, India, 1998; 188p. [Google Scholar]
- Chandra, S. Some new leaf-spot diseases from Allahabad (India). Beih. Nova Hedwig. 1974, 47, 35–102. [Google Scholar]
- Sakalidis, M.L.; Hardy, G.E.S.; Burgess, T.I. Endophytes as potential pathogens of the baobab species Adansonia gregorii: A focus on the Botryosphaeriaceae. Fungal Ecol. 2011, 4, 1–14. [Google Scholar] [CrossRef]
- Al Raish, S.M.; Saeed, E.E.; Sham, A.; Alblooshi, K.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular characterization and disease control of stem canker on royal poinciana (Delonix regia) caused by Neoscytalidium dimidiatum in the United Arab Emirates. Int. J. Mol. Sci. 2020, 21, 1033. [Google Scholar] [CrossRef] [PubMed]
- Galea, V.J. Use of stem ımplanted bioherbicide capsules to manage an ınfestation of Parkinsonia aculeata in Northern Australia. Plants 2021, 10, 1909. [Google Scholar] [CrossRef]
- Anonymous. Index of Plant Diseases in the United States. In U.S.D.A. Agriculture Handbook; U.S. Department of Agriculture: Washington, DC, USA, 1960; Volume 165, pp. 1–531. [Google Scholar]
- Alidadi, A.; Kowsari, M.; Javan-Nikkhah, M.; Jouzani, G.R.S.; Rastaghi, M.E. New pathogenic and endophytic fungal species associated with Persian oak in Iran. Eur. J. Plant Pathol. 2019, 155, 1017–1032. [Google Scholar] [CrossRef]
- French, A.M. California Plant Disease Host Index. Part 1: Fruit and Nuts; California Department of Food and Agriculture: Sacramento, CA, USA, 1987; 39p.
- Wilson, E.E. The pycnidial stage of the walnut branch wilt fungus, Exosporina fawcettii. Phytopathology 1949, 53, 705–712. [Google Scholar]
- Sommer, N.F. Sunburn predisposes walnut trees to branch-wilt. Phytopathology 1955, 45, 607–613. [Google Scholar]
- Phillips, A.J.L.; Oudemans, P.V.; Correia, A.; Alves, A. Characterisation and epitypification of Botryosphaeria corticis, the cause of blueberry cane canker. Fung Divers. 2006, 21, 141–155. [Google Scholar]
- Chen, S.F.; Fichtner, E.; Morgan, D.P.; Michailides, T.J. First report of Lasiodiplodia citricola and Neoscytalidium dimidiatum causing death of graft union of English walnut in California. Plant Dis. 2013, 97, 993. [Google Scholar] [CrossRef]
- Chen, S.; Morgan, D.P.; Hasey, J.K.; Anderson, K.; Michailides, T.J. Phylogeny, morphology, distribution, and pathogenicity of Botryosphaeriaceae and Diaporthaceae from English walnut in California. Plant Dis. 2014, 98, 636–652. [Google Scholar] [CrossRef] [PubMed]
- Derviş, S.; Türkölmez, Ş.; Çiftçi, O.; Ulubaş Serçe, Ç.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing black canker and root rot of walnut in Turkey. Plant Dis. 2019, 103, 2129. [Google Scholar] [CrossRef]
- Çiftçi, O.; Özer, G.; Türkölmez, Ş.; Derviş, S. Lasiodiplodia theobromae and Neoscytalidium dimidiatum associated with grafted walnut (Juglans regia L.) decline in Turkey. J. Plant Dis. Prot. 2023, 130, 1117–1128. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Maharachchikumbura, S.S. Neoscytalidium dimidiatum as one of the fungal agents associated with walnut decline in Iran. J. Nuts 2022, 13, 31–39. [Google Scholar] [CrossRef]
- Güney, İ.G.; Özer, G.; Turan, İ.; Koşar, İ.; Derviş, S. First report of Neoscytalidium dimidiatum causing foliar and stem blight of lavender in Turkey. J. Plant Pathol. 2021, 103, 1347–1348. [Google Scholar] [CrossRef]
- Özer, G.; Günen, T.U.; Koşar, İ.; Güler, İ.G.; Derviş, S. First report of Neoscytalidium dimidiatum causing blight of Melissa officinalis in Turkey. J. Plant Dis. Prot. 2022, 129, 197–199. [Google Scholar] [CrossRef]
- Alkan, M.; Özer, G.; Koşar, I.; Güney, I.G.; Derviş, S. First report of leaf blight of Turkish oregano (Origanum onites) caused by Neoscytalidium dimidiatum in Turkey. J. Plant Pathol. 2022, 104, 471. [Google Scholar] [CrossRef]
- Derviş, S.; Güney, İ.G.; Koşar, İ.; Bozoğlu, T.; Özer, G. First report of Neoscytalidium novaehollandiae on common sage (Salvia officinalis). Australas. Plant Dis. 2021, 16, 19. [Google Scholar] [CrossRef]
- Kadkhoda-Hematabadi, S.; Mohammadi, H.; Sohrabi, M. Morphological and molecular identification of plant pathogenic fungi associated with necrotic wood tissues of pomegranate trees in Iran. J. Plant Pathol. 2023, 105, 465–479. [Google Scholar] [CrossRef]
- Ghasemi-Sardareh, R.; Mohammadi, H. Characterization and pathogenicity of fungal trunk pathogens associated with declining of neem (Azadirachta indica A. Juss) trees in Iran. J. Plant Pathol. 2020, 102, 1159–1171. [Google Scholar] [CrossRef]
- Ahmad, S.; Iqbal, S.H.; Khalid, A.N. Fungi of Pakistan. Sultan Ahmad Mycological Society of Pakistan; Department of Botany, University of the Punjab: Lahore, Pakistan, 1997; 248p. [Google Scholar]
- Abdulrahman, D.N.; Haleem, R.A. Morphological and molecular characterization of Neoscytalidium isolates that cause canker and dieback in eucalyptus and chinaberry trees in Iraq. Plant Prot. Sci. 2023, 59, 92–105. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Papadopoulos, C. A Second List of Cyprus Fungi; Government of Cyprus, Department of Agriculture: Nicosia, Cyprus, 1957; 38p.
- Paxton, J.D.; Wilson, E.E.; Davis, J.R. Branch wilt disease of fig caused by Hendersonula toruloidea. Plant Dis. Rep. 1964, 48, 142. [Google Scholar]
- Oksal, E. Prevalence, molecular characterization, and variety reactions of Neoscytalidium novaehollandiae on mulberries in Turkey. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12716. [Google Scholar] [CrossRef]
- Meredith, D.S. Tip rot of banana fruits in Jamaica. Trans. Br. Mycol. Soc. 1963, 46, 473–481. [Google Scholar] [CrossRef]
- Meredith, D.S. Fungal diseases of bananas in Hawaii. Plant Dis. Rep. 1969, 53, 63–66. [Google Scholar]
- Raabe, R.D.; Conners, I.L.; Martinez, A.P. Checklist of Plant Diseases in Hawaii; Information Text Series No. 22; College of Tropical Agriculture and Human Resources, University of Hawaii: Honolulu, HI, USA, 1981; 313p. [Google Scholar]
- Al-Tememe, Z.A.M.; Lahuf, A.; Abdalmoohsin, R.G.; Al-Amirry, A.T. Occurrence, identification, pathogenicity and control of Neoscytalidium dimidiatum fungus, the causal agent of sooty canker on Eucalyptus camaldulensis in Kerbala Province of Iraq. Plant Arch. 2019, 19, 31–38. [Google Scholar]
- Ismail, S.I.; Ahmad Dahlan, K.; Abdullah, S.; Zulperi, D. First Report of Neoscytalidium dimidiatum causing fruit rot on guava (Psidium guajava L.) in Malaysia. Plant Dis. 2021, 105, 220. [Google Scholar] [CrossRef]
- Panahandeh, S.; Mohammadi, H.; Gramaje, D. Trunk disease fungi associated with Syzygium cumini in Iran. Plant Dis. 2019, 103, 711–720. [Google Scholar] [CrossRef]
- Williams, T.H.; Liu, P.S.W. A host list of plant diseases in Sabah, Malaysia. Phytopathol. Pap. 1976, 19, 1–67. [Google Scholar]
- Suwannarach, N.; Kumla, J.; Lumyong, S. Leaf spot on cattleya orchid caused by Neoscytalidium orchidacearum in Thailand. Can. J. Plant Pathol. 2018, 40, 109–114. [Google Scholar] [CrossRef]
- Chang, C.W.; Chen, C.Y.; Wang, C.L.; Chung, W.H. First report of leaf blight on Cattleya × hybrid caused by Neoscytalidium dimidiatum in Taiwan. J. Plant Pathol. 2020, 102, 921. [Google Scholar] [CrossRef]
- Derviş, S.; Türkölmez, Ş.; Güney, İ.G.; Alkan, M.; Ozer, G. First report of needle blight of blue spruce (Picea pungens) caused by Neoscytalidium dimidiatum in Turkey. J. Plant Pathol. 2023, 105, 1195–1196. [Google Scholar] [CrossRef]
- Nourian, A.; Salehi, M.; Safaie, N.; Khelghatibana, F.; Abdollahzadeh, J. Fungal canker agents in apple production hubs of Iran. Sci. Rep. 2021, 11, 22646. [Google Scholar] [CrossRef] [PubMed]
- Sha, S.; Wang, Z.; Hao, H.; Wang, L.; Feng, H. First report of Neoscytalidium dimidiatum inducing canker disease on apple trees in China. J. Plant Pathol. 2022, 104, 1149–1150. [Google Scholar] [CrossRef]
- Ören, E.; Palacıoğlu, G.; Ozan, G.N.; Çelik, K.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with canker and branch dieback on cherry trees in Turkey. J. Plant Pathol. 2022, 104, 391. [Google Scholar] [CrossRef]
- Ören, E.; Palacıoğlu, G.; Ozan, G.N.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with branch dieback and stem cankers on plum in Turkey. J. Plant Pathol. 2022, 104, 1167. [Google Scholar] [CrossRef]
- English, H.; Davis, J.R.; Devay, J.E. Relationship of Botryosphaeria dothidea and Hendersonula toruloidea to a canker disease of almond. Phytopathology 1975, 65, 114–122. [Google Scholar] [CrossRef]
- Burgess, T.I.; Barber, P.A.; St., J.; Hardy, G.E. Botryosphaeria spp. associated with eucalypts in Western Australia, including the description of Fusicoccum macroclavatum sp. nov. Australas. Plant Pathol. 2005, 34, 557–567. [Google Scholar] [CrossRef]
- Oksal, E.; Özer, G. First report of shoot blight and branch canker of Pyrus communis by Neoscytalidium novaehollandiae in Turkey. J. Plant Pathol. 2021, 103, 673–674. [Google Scholar] [CrossRef]
- Espargham, N.; Mohammadi, H.; Gramaje, D. A Survey of Trunk Disease Pathogens within Citrus Trees in Iran. Plants 2020, 9, 754. [Google Scholar] [CrossRef]
- Alananbeh, K.M.; Al-Qasim, M.; Gharaibeh, A.; Al-Hiary, H.A. First report of shoot blight caused by Neoscytalidium dimidiatum on citrus in Jordan. Plant Dis. 2020, 104, 571. [Google Scholar] [CrossRef]
- Calavan, E.C.; Wallace, J.M. Hendersonula toruloidea Nattrass on Citrus in California. Phytopathology 1954, 44, 635–639. [Google Scholar]
- Adesemoye, A.O.; Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Lynch, S.C.; Eskalen, A. Identification of species of Botryosphaeriaceae causing bot gummosis in citrus in California. Plant Dis. 2014, 98, 55–61. [Google Scholar] [CrossRef]
- Doidge, E.M. The South African fungi and lichens to the end of 1945. Bothalia 1950, 5, 1–1094. [Google Scholar]
- Ogawa, J.M. The occurrence of Hendersonula toruloidea Nattrass on Populus species in California. Plant Dis. Rep. 1954, 38, 238. [Google Scholar]
- Derviş, S.; Özer, G.; Türkölmez, Ş. First report of Neoscytalidium novaehollandiae causing stem blight on tomato in Turkey. J. Plant Pathol. 2020, 102, 1339–1340. [Google Scholar] [CrossRef]
- Derviş, S.; Özer, G.; Türkölmez, Ş. First report of Neoscytalidium dimidiatum causing tuber rot of potato in Turkey. J. Plant Pathol. 2020, 102, 1295–1296. [Google Scholar] [CrossRef]
- Hashemi, H.; Mohammadi, H.; Abdollahzadeh, J. Symptoms and fungi associated with elm trees decline in Iran. Eur. J. For. Res. 2017, 136, 857–879. [Google Scholar] [CrossRef]
- Wangikar, P.D.; Raut, J.G.; Gopalkrishna, N. Drying of grape vines caused by Hendersonula toruloidea. Indian Phytopathol. 1969, 22, 403–404. [Google Scholar]
- Sarbhoy, A.K.; Lal, G.; Varshney, J.L. Fungi of India (1967–1971); Navyug Traders: New Delhi, India, 1971; 148p. [Google Scholar]
- Natour, R.M.; Ahmed, J.M. Control of branch wilt disease of grape. Plant Dis. Rep. 1969, 53, 152–153. [Google Scholar]
- Ören, E.; Palacıoğlu, G.; Koca, G.; Ozan, G.N.; Bayraktar, H. First report of Neoscytalidium dimidiatum causing branch dieback and canker on apple in Turkey. J. Plant Pathol. 2022, 104, 429. [Google Scholar] [CrossRef]
- Xu, M.; Liu, C.L.; Fu, Y.; Liao, Z.W.; Guo, P.Y.; Xiong, R.; Cheng, Y.; Wei, S.S.; Huang, J.Q.; Tang, H. Molecular characterization and expression analysis of pitaya (Hylocereus polyrhizus) HpLRR genes in response to Neoscytalidium dimidiatum infection. BMC Plant Biol. 2020, 20, 160. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.F.; Lawrence, D.P.; Morales-Cruz, A.; Travadon, R.; Minio, A.; Hernandez-Martinez, R.; Rolshausen, P.E.; Baumgartner, K.; Cantu, D. Phylogenomics of plant-associated Botryosphaeriaceae Species. Front. Microbiol. 2021, 12, 652802. [Google Scholar] [CrossRef]
- Türkölmez, Ş.; Eren, A.; Özer, G.; Derviş, S. The effect of Talaromyces funiculosus ST976 isolated from pistacia vera rhizosphere on phosphorus solubility in soil samples with different physicochemical properties. J. Agric. Nat. 2022, 25, 1077–1085. [Google Scholar] [CrossRef]
- Kuan, C.S.; Ng, K.P.; Yew, S.M.; Umar Meleh, H.; Seow, H.F.; How, K.N.; Yeo, S.K.; Jee, J.M.; Tan, Y.C.; Yee, W.Y.; et al. Comparative genomic and phenotypic analyses of pathogenic fungi Neoscytalidium dimidiatum and Bipolaris papendorfii isolated from human skin scraping. Braz. J. Microbiol. 2023, 54, 1351–1372. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Khan, A.H. Some new diseases of Citrus in Pakistan. Mycopathol. Mycol. Appl. 1959, 10, 289–302. [Google Scholar] [CrossRef]
- Paxton, J.D.; Wilson, E.E. Anatomical and physiological aspects of branch wilt disease of Persian walnut. Phytopathology 1965, 55, 21. [Google Scholar]
- Hassan, W.A.; Haleem, R.A.; Hassan, P.H. Effect of heat-stress predisposition on the development of sooty canker caused by Neoscytalidium dimidiatum (Penz.) Crous and Slippers. Acta Agrobot. 2011, 64, 207–212. [Google Scholar] [CrossRef]
- Zhu, X.M.; Liu, X.F. A new species and genus distribution record from China: Neoscytalidium novaehollandiae. Indian. J. Microbiol. 2012, 52, 565–568. [Google Scholar] [CrossRef]
- Corlett, R.T. Frugivory and seed dispersal by vertebrates in tropical and subtropical Asia: An update. Glob. Ecol. Conserv. 2017, 11, 1–22. [Google Scholar] [CrossRef]
- Belair, M.; Restrepo-Leal, J.D.; Praz, C.; Fontaine, F.; Remond, C.; Fernandez, O.; Besaury, L. Botryosphaeriaceae gene machinery: Correlation between diversity and virulence. Fungal Biol. 2023, 127, 1010–1031. [Google Scholar] [CrossRef] [PubMed]
- Nagel, J.H.; Wingfield, M.J.; Slippers, B. Increased abundance of secreted hydrolytic enzymes and secondary metabolite gene clusters define the genomes of latent plant pathogens in the Botryosphaeriaceae. BMC Genom. 2021, 22, 589. [Google Scholar] [CrossRef] [PubMed]
- Reveglia, M.; Masi, A.; Cimmino, T.; Cinelli, L.; Mugnai, A. Evidente phytotoxins produced by Lasiodiplodia laeliocattleyae involved in Botryosphaeria dieback of grapevines in Brazil. Phytopathol Mediterr. 2019, 58, 207–211. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Robert-Siegwald, G.; Fernandez, O.; Leal, C.; Villaume, S.; Guise, J.F.; Abou-Mansour, E.; Lebrun, M.H.; Fontaine, F. Diversity of Neofusicoccum parvum for the production of the phytotoxic metabolites (-)-terremutin and (R)-mellein. J. Fungi 2022, 8, 319. [Google Scholar] [CrossRef]
- Bénard-Gellon, M.; Farine, S.; Goddard, M.L.; Schmitt, M.; Stempien, E.; Pensec, F.; Laloue, H.; Mazet-Kieffer, F.; Fontaine, F.; Larignon, P.; et al. Toxicity of extracellular proteins from Diplodia seriata and Neofusicoccum parvum involved in grapevine Botryosphaeria dieback. Protoplasma 2015, 252, 679–687. [Google Scholar] [CrossRef]
- Cimmino, A.; Cinelli, T.; Evidente, M.; Masi, M.; Mugnai, L.; Silva, M.A.; Michereff, S.J.; Surico, G.; Evidente, A. Phytotoxic fungal exopolysaccharides produced by fungi involved in grapevine trunk diseases. Nat. Prod. Commun. 2016, 10, 1481–1484. [Google Scholar] [CrossRef]
- Andolfi, A.; Maddau, L.; Cimmino, A.; Linaldeddu, B.T.; Basso, S.; Deidda, A.; Serra, S.; Evidente, A. Lasiojasmonates A–C, three jasmonic acid esters produced by Lasiodiplodia sp., a grapevine pathogen. Phytochemistry 2014, 103, 145–153. [Google Scholar] [CrossRef]
- Yu, C.; Diao, Y.; Lu, Q.; Zhao, J.; Cui, S.; Xiong, X.; Lu, A.; Zhang, X.; Liu, H. Comparative genomics reveals evolutionary traits, mating strategies, and pathogenicity-related genes variation of Botryosphaeriaceae. Front. Microbiol. 2022, 13, 800981. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Xu, M.; Liu, C.L.; Luo, J.; Qi, Z.; Yan, Z.; Fu, Y.; Wei, S.S.; Tang, H. Transcriptomic de novo analysis of pitaya (Hylocereus polyrhizus) canker disease caused by Neoscytalidium dimidiatum. BMC Genom. 2019, 20, 10. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Z.; Wei, S.; Xie, J.; Huang, J.; Li, D.; Hu, W.; Li, H.; Tang, H. Identification of RT-qPCR reference genes suitable for gene function studies in the pitaya canker disease pathogen Neoscytalidium dimidiatum. Sci. Rep. 2022, 12, 22357. [Google Scholar] [CrossRef]
- Arkam, M.; Alves, A.; Lopes, A.; Cechova, J.; Pokluda, R.; Eichmeier, A.; Zitouni, A.; Mahamedi, A.E.; Berraf-Tebbal, A. Diversity of Botryosphaeriaceae causing grapevine trunk diseases and their spatial distribution under different climatic conditions in Algeria. Eur. J. Plant Pathol. 2021, 161, 933–952. [Google Scholar] [CrossRef]
- Xian, X.; Lin, S.; Zhu, G.; Wei, X.; Qin, W.; Zhong, Y. Indoor virulence and field effects of fungicides on pitaya canker. J. South. Agric. 2018, 49, 1338–1345. [Google Scholar]
- Noegrahati, S.; Sulasmi, S.; Hemadi, E.; Asviastuti, S. Dissipation pattern of azoxystrobin and difenoconazole in red dragon fruit (Hylocereus polyrhizus) cultivated in Indonesian highland (West Java) and coastal area (D.I. Jogyakarta) and its implication for dietary risk assessment. Food Qual. Saf. 2019, 3, 99–106. [Google Scholar] [CrossRef]
- Eraslan Sür, A.; Oksal, E. In vitro efficiency of some fungicides against Neoscytalidium dimidiatum (Penz.) Crous and Slippers causing sudden shoot dry on apricot trees. Turk. J. Agric.-Food Sci. Technol. 2021, 9, 797–802. [Google Scholar] [CrossRef]
- Riska, J.; Budiyanti, T.; Husada, E.D.; Indriyani, N.L.P.; Hadiati, S.; Muas, I.; Mansyah, E. Stem canker of dragon fruit (Hylocereus polyrhizus): Neoscytalidium sp. is a pathogen of the disease and its control using sodium salt. Plant Protect. Sci. 2023, 59, 245–255. [Google Scholar] [CrossRef]
- Campos-Rivero, G.; Sánchez-Teyer, L.F.; De la Cruz-Arguijo, E.A.; Ramírez-González, M.S.; Larralde-Corona, C.P.; Narváez-Zapata, J.A. Bioprospecting for fungi with potential pathogenic activity on leaves of Agave tequilana Weber var. Azul. J. Phytopathol. 2019, 167, 283–294. [Google Scholar] [CrossRef]
- Taguiam, J.D.; Evallo, E.; Bengoa, J.; Maghirang, R.; Balendres, M.A. Susceptibility of the three dragon fruit species to stem canker and growth inhibition of Neoscytalidium dimidiatum by chemicals. J. Plant Pathol. 2020, 102, 1077–1084. [Google Scholar] [CrossRef]
- Ratanaprom, S.; Nakkanong, K.; Nualsri, C.; Jiwanit, P.; Rongsawat, T.; Woraathakorn, N. Overcoming encouragement of dragon fruit plant (Hylocereus undatus) against stem brown spot disease caused by Neoscytalidium dimidiatum using Bacillus subtilis combined with sodium bicarbonate. Plant Pathol. J. 2021, 37, 205–214. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, R.; Yuan, Z.; Chen, P. Biological prevention and control of pitaya fruit canker disease using endophytic fungi isolated from papaya. Arch. Microbiol. 2021, 203, 4033–4040. [Google Scholar] [CrossRef]
- Al Hamad, B.M.; Al Raish, S.M.; Ramadan, G.A.; Saeed, E.E.; Alameri, S.S.A.; Al Senaani, S.S.; AbuQamar, S.F.; El-Tarabily, K.A. Effectiveness of augmentative biological control of Streptomyces griseorubens UAE2 depends on 1-aminocyclopropane-1-carboxylic acid deaminase activity against Neoscytalidium dimidiatum. J. Fungi 2021, 7, 885. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chen, X.; Xie, L.; Zhang, Y.; Zeng, F.; Long, Y.; Ren, L.; Qi, X.; Wei, J. Biocontrol potential of lipopeptides produced by Paenibacillus polymyxa AF01 against Neoscytalidium dimidiatum in pitaya. Front. Microbiol. 2023, 14, 1188722. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gao, Y.; Li, H.; Wang, D.; Li, J.; Hu, X.; Huang, X.; Lin, M.; Tang, Y.; Liu, Z. Identification and characterization of biocontrol agent Lysinibacillus boronitolerans P42 against Cerrena unicolor that causes root rot of arecanut palm. Arch. Microbiol. 2023, 205, 157. [Google Scholar] [CrossRef] [PubMed]
- Travadon, R.; Lawrence, D.P.; Li, S.; Trouillas, F. Evaluation of biological control agents for the protection of almond pruning wounds against infection by fungal canker pathogens. Phytopathology 2023, 113, 1417–1427. [Google Scholar] [CrossRef]
- Du, B.D.; Ngoc, D.T.B.; Thang, N.D.; Tuan, L.N.A.; Thach, B.D.; Hien, N.Q. Synthesis and in vitro antifungal efficiency of alginate-stabilized Cu2O-Cu nanoparticles against Neoscytalidium dimidiatum causing brown spot disease on dragon fruit plants (Hylocereus undatus). Vietnam J. Chem. 2019, 57, 318–323. [Google Scholar] [CrossRef]
- Tuan, L.N.A.; Du, B.D.; Ha, L.D.T.; Dzung, L.T.K.; Van Phu, D.; Hien, N.Q. Induction of chitinase and brown spot disease resistance by oligochitosan and nanosilica–oligochitosan in dragon fruit plants. Agric. Res. 2019, 8, 184–190. [Google Scholar] [CrossRef]
- Acay, H.; Yildirim, A.; Güney, I.G.; Derviş, S. Morchella esculenta-based chitosan bionanocomposites: Evaluation as an antifungal agent. J. Food Process. Preserv. 2022, 46, e17117. [Google Scholar] [CrossRef]
- Hashem, A.H.; Abdelaziz, A.M.; Hassanin, M.M.H.; Al-Askar, A.A.; AbdElgawad, H.; Attia, M.S. Potential impacts of clove essential oil nanoemulsion as bio fungicides against Neoscytalidium blight disease of Carum carvi L. Agronomy 2023, 13, 1114. [Google Scholar] [CrossRef]
- Duong, N.L.; Nguyen, V.M.; Tran, T.A.N.; Phan, T.D.T.; Tran, T.B.Y.; Do, B.L.; Phung Anh, N.; Nguyen, T.A.T.; Ho, T.G.; Nguyen, T. Durian shell-mediated simple green synthesis of nanocopper against plant pathogenic fungi. ACS Omega 2023, 8, 10968–10979. [Google Scholar] [CrossRef]
- Bragard, C.; Baptista, P.; Chatzivassiliou, E.; Di Serio, F.; Gonthier, P.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P. Pest categorisation of Neoscytalidium dimidiatum. EFSA J. 2023, 21, e08001. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic tree of Neoscytalidium resulting from a Bayesian analysis of the combined ITS, tef1, tub2 sequence alignment. Maximum Likelihood and Maximum Parsimony bootstrap support values (ML/MP-BS > 50%) and Bayesian posterior probabilities (PP > 0.90) are shown at the nodes. Ex-type strains and taxonomic novelties are indicated in bold font. The last accepted species names are shown in superscript where species were synonymized in this study; species names on which the name of the clade is based are in bold superscript. The tree was rooted to Botryosphaeria dothidea (CBS 115476).
Figure 1.
Phylogenetic tree of Neoscytalidium resulting from a Bayesian analysis of the combined ITS, tef1, tub2 sequence alignment. Maximum Likelihood and Maximum Parsimony bootstrap support values (ML/MP-BS > 50%) and Bayesian posterior probabilities (PP > 0.90) are shown at the nodes. Ex-type strains and taxonomic novelties are indicated in bold font. The last accepted species names are shown in superscript where species were synonymized in this study; species names on which the name of the clade is based are in bold superscript. The tree was rooted to Botryosphaeria dothidea (CBS 115476).

Figure 2.
Morphological characteristics of Neoscytalidium, 4-day-old colony on PDA (a) and 7-day-old colony on PDA (b), arthroconidia (c), black conidiomata (pycnidia) on wood chips (d), conidia formed in pycnidia (e), and conidiogenous cells (f).
Figure 2.
Morphological characteristics of Neoscytalidium, 4-day-old colony on PDA (a) and 7-day-old colony on PDA (b), arthroconidia (c), black conidiomata (pycnidia) on wood chips (d), conidia formed in pycnidia (e), and conidiogenous cells (f).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Derviş, S.; Özer, G. Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review. J. Fungi 2023, 9, 1048. https://doi.org/10.3390/jof9111048
AMA Style
Derviş S, Özer G. Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review. Journal of Fungi. 2023; 9(11):1048. https://doi.org/10.3390/jof9111048
Chicago/Turabian StyleDerviş, Sibel, and Göksel Özer. 2023. "Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review" Journal of Fungi 9, no. 11: 1048. https://doi.org/10.3390/jof9111048
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.