

Morpho-Phylogenetic Evidence Reveals Novel Species and New Records of Botryosphaeriaceae in China and Thailand

1
School of Life Science and Technology, Center for Informational Biology, University of Electronic Science and Technology of China, Chengdu 611731, China
2
Center of Excellence in Fungal Research, School of Science, Mae Fah Luang University, Chiang Rai 57100, Thailand
3
School of Pharmacy, Guizhou University of Traditional Chinese Medicine, Guiyang 550025, China
4
Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(11), 1051; https://doi.org/10.3390/jof9111051
Submission received: 29 August 2023
/
Revised: 23 October 2023
/
Accepted: 24 October 2023
/
Published: 26 October 2023
(This article belongs to the Special Issue Recent Advances in Taxonomy, Phylogeny and Evolution of Fungi)
Abstract
:Species in the Botryosphaeriaceae are common plant pathogens, endophytes, and saprobes found on a variety of mainly woody hosts. Botryosphaeriaceae is a high-profile fungal family whose genera have been subjected to continuous revisions in recent years. Surveys conducted during 2019 and 2020 on several decaying woody hosts (from dead arial twigs, branches, stems, bark, and seed pods) in China and Thailand revealed a high diversity of Botryosphaeriaceae fungi. Identification of 16 Botryosphaeriaceae isolates was carried out based on both morphological characteristics and phylogenetic analyses of combined ITS, LSU, tef1-α, and tub2 sequence data. Four novel species (Dothiorella ovata, Do. rosacearum, Do. septata, and Lasiodiplodia delonicis) and seven previously known species (Botryosphaeria fujianensis, Diplodia mutila, Di. seriata, L. crassispora, L. mahajangana, Macrophomina euphorbiicola and Sphaeropsis eucalypticola) were identified while new hosts and geographical records were reported. This study indicates that the fungal family Botryosphaeriaceae seems to be common and widespread on a broad range of hosts in China and Thailand.
1. Introduction
The order Botryosphaeriales (Dothideomycetes) was established by Schoch et al. [1], including a single family Botryosphaeriaceae. A recent comprehensive study by Phillips et al. [2] accepted six families, viz. Aplosporellaceae, Botryosphaeriaceae, Melanopsaceae, Phyllostictaceae, Planistromellaceae, and Saccharataceae are in this order based on morphological and phylogenetic analysis. Members of Botryosphaeriales have worldwide distribution on many different host plants [3,4,5,6,7,8] and occur as endophytes, pathogens, and saprobes. As opportunistic pathogens, they are of considerable importance to agriculture, horticulture, and forestry [9,10] while causing severe diseases of economically important crops and plants, leading to huge economic losses [11,12].
The family Botryosphaeriaceae was established by Theissen and Sydow [13] to accommodate three genera, Botryosphaeria, Dibotryon, and Phaeobotryon, with Botryosphaeria as the type genus [1]. Subsequently, Kirk et al. [14] estimated 26 genera within the family Botryosphaeriaceae, while Liu et al. [4] reevaluated the family and recognized 29 genera. Up to date, 22 genera with more than 200 species are accepted within the family, based on morphology and molecular evidence [2,15,16,17,18].
During an investigation of Botryosphaeriaceae diversity in China and Thailand, a collection of 16 Botryosphaeriaceae-like isolates was obtained from several arial parts of the decaying woody hosts. A multi-gene phylogeny based on combined ITS, LSU, tef1-α, and tub2, coupled with morphological comparisons, was carried out to confirm the taxonomic placement. Additionally, this study extended the taxonomic framework of Botryosphaeriaceae by discovering new species, new host records, and new geographical records in China and Thailand.
2. Materials and Methods
2.1. Specimen Collection, Examination, and Single Spore Isolation
Samples of decaying woody plants were collected from Chiang Rai and Chiang Mai Provinces in Thailand and from Sichuan Province in China between June 2019 and November 2020. Samples were brought to the laboratory by placing them in brown craft paper bags, and the sampling information (date, host, place, GPS, etc.) was recorded.
Morphological observations of fungal structures were made using a LEICA EZ4 dissecting microscope following the method described in Chomnunti et al. [19]. The fungal structures were transferred to a small drop of double distilled water on a clean slide and covered with a glass coverslip. Photomicrographs of the fungal specimens were captured using a Nikon ECLIPSE Ni compound microscope fitted with a Nikon DS-Ri2 digital camera. Macro-morphological structures were photographed with a Discovery V.8 stereo microscope fitted with a CARL ZEISS Axio Cam ERc5S microscope camera. All measurements were made with the Tarosoft (R) Image Frame Work (IFW) program [20], and the images were processed with Adobe Photoshop CC extended version 21.1.2.
Single spore isolations were carried out as described by Chomnunti et al. [19], and fruiting body contents were transferred to a drop of sterile water on a sterilized spot plate. This spore suspension was spread over the Petri dishes containing potato dextrose agar (PDA) and incubated at 25 °C for 12 to 24 h. Germinated spores were transferred onto fresh PDA media plates. These culture plates were incubated at 25 °C in incubators, and colony characteristics were observed and recorded after one week following the method described in Rayner [21]. A total of 56 isolates have been obtained, among which 16 isolates belong to Botryosphaeriaceae. In this study, we focus only on the fungal taxa of Botryosphaeriaceae. To induce sporulation, cultures were transferred onto fresh PDA media plates using sterile toothpicks or pine needles. The induction results were observed after incubating under near-ultraviolet light for 14–30 d at 25 °C.
Herbarium specimens were deposited in the herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand, and Guizhou Academic of Agriculture Sciences (GZAAS), China. Axenic cultures were deposited in Mae Fah Luang University Culture Collection (MFLUCC) and Guizhou Culture Collection (GZCC). In deciding whether we have new species, we followed the papers of Chethana et al. [22] and Pem et al. [23]. The descriptions are added to the GMS database [24].
2.2. DNA Extraction, PCR Amplification and Sequencing
In a sterile environment, fungal mycelium (about 50–100 mg) was scraped using a sterilized toothpick from the colonies grown on PDA media at 25 °C for 2 weeks and then transferred to sterilized 1.5 mL microcentrifuge tubes and maintained at −20 °C for long term storage. Ezup Column Fungi Genomic DNA Purification Kit (Sangon Biotech, Shanghai, China) was used to extract DNA according to the manufacturer’s instructions. The amplifications were performed in a 25 μL reaction volume containing 8.5 μL ddH2O, 12.5 μL 2 × PCR Master Mix (Green) (TsingKe Co., Beijing, China), 2 μL of DNA template, and 1 μL of each primer. The genes, primers, and amplification conditions used in this study are listed in Table 1. The PCR products were analyzed by 1.2% agarose gels containing the Safeview DNA stain and sent to Tsingke Biotechnology Co., Ltd. (Chengdu, China) for sequencing.
2.3. Sequence Alignment and Phylogenetic Analysis
Sequences generated in this study were checked and assembled using BioEdit v.7.0.9 [29] to assure the sequence quality. The closest taxa to the strains obtained in this study were determined with standard nucleotide BLAST searches in NCBI (http://www.ncbi.nlm.nih.gov/, accessed on 20 July 2022). According to the BLAST results and previous literature, appropriate sequences were determined and downloaded from GenBank to construct phylogenetic analysis. Two phylogenetic trees were constructed, one for the whole family Botryosphaeriaceae (Figure 1) and the other for the genus Dothiorella (Figure 2). Details of the isolates used in this study are listed in Table 2, where two strains of Pseudofusicoccum adansoniae (CBS 122055, CBS 122056) and Neofusicoccum parvum (CBS 110301, CMW 9081) were selected as the outgroup taxa for Botryosphaeriaceae analyses and Dothiorella analysis respectively. The sequences were aligned using MAFFT v.7 online (https://mafft.cbrc.jp/alignment/server/, accessed on 1 August 2023) and AliView [30], and the results were checked using BioEdit [29] and manually edited where necessary. The concatenation of different genes was conducted using SequenceMatrix 1.8 [31]. The NEXUS and Phylip files for phylogenetic analyses were obtained using AliView [30]. Phylogenetic analyses of the combined sequence data were performed using maximum likelihood (ML), maximum parsimony (MP), and Bayesian inference (BI) methods, as detailed in Dissanayake et al. [32]. The best model of evolution was determined using MrModeltest v2 [33]. The ML analysis was accomplished using RAxML GUI v. 1.3.1 [34], the MP analysis was performed using PAUP v.4.0b10 [35], and the BI analysis was conducted in MrBayes v 3.2.6 [36]. Phylogenetic trees were visualized with FigTree v.1.4.0 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 11 August 2023) and further edited in Adobe Illustrator 2020 (Adobe Systems Inc., Lehi, UT, USA). The final alignment was submitted to Figshare (https://figshare.com, at https://doi.org/10.6084/m9.figshare.24187500, accessed on 22 September 2023).
3. Results
3.1. Phylogenetic Analysis
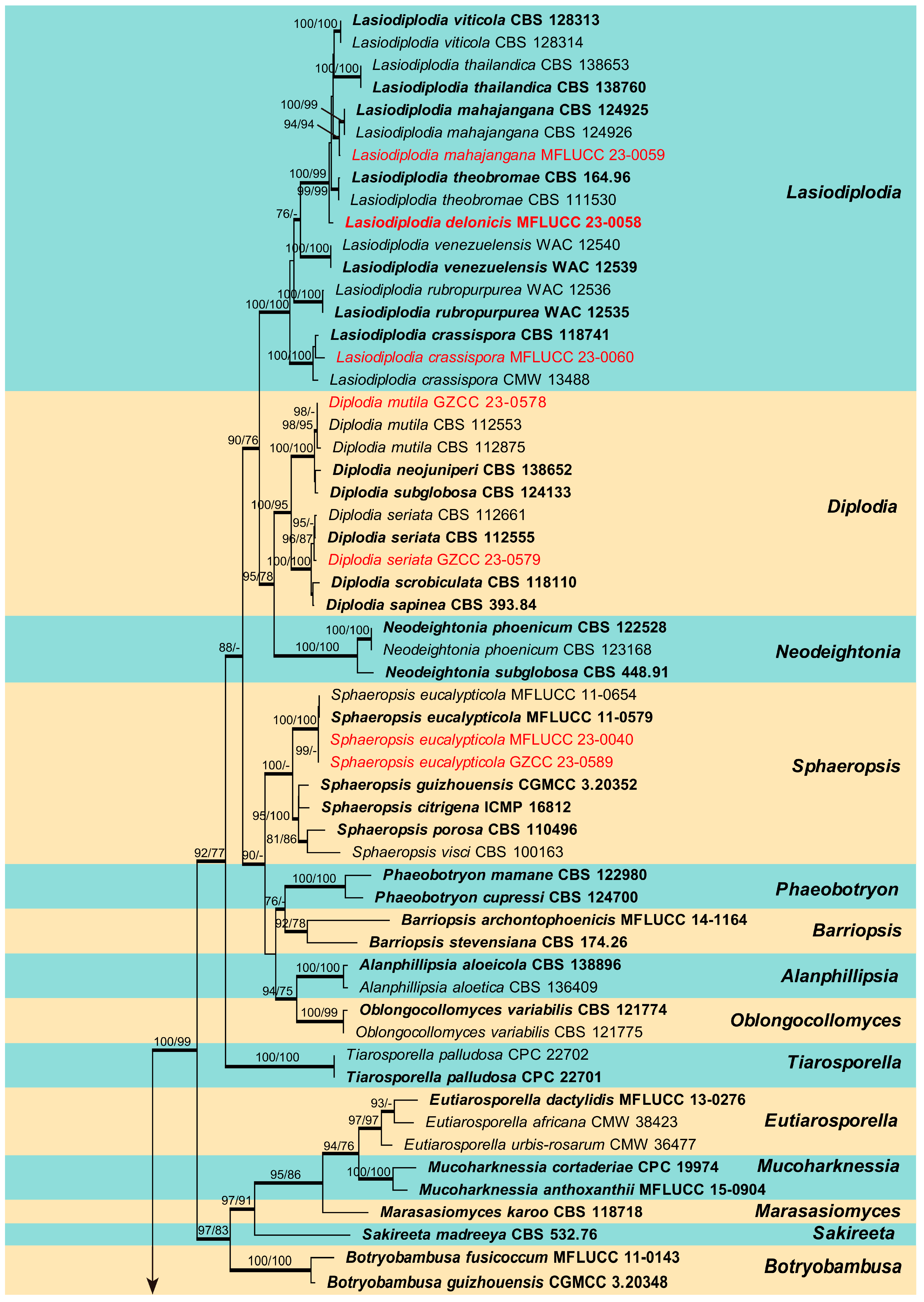
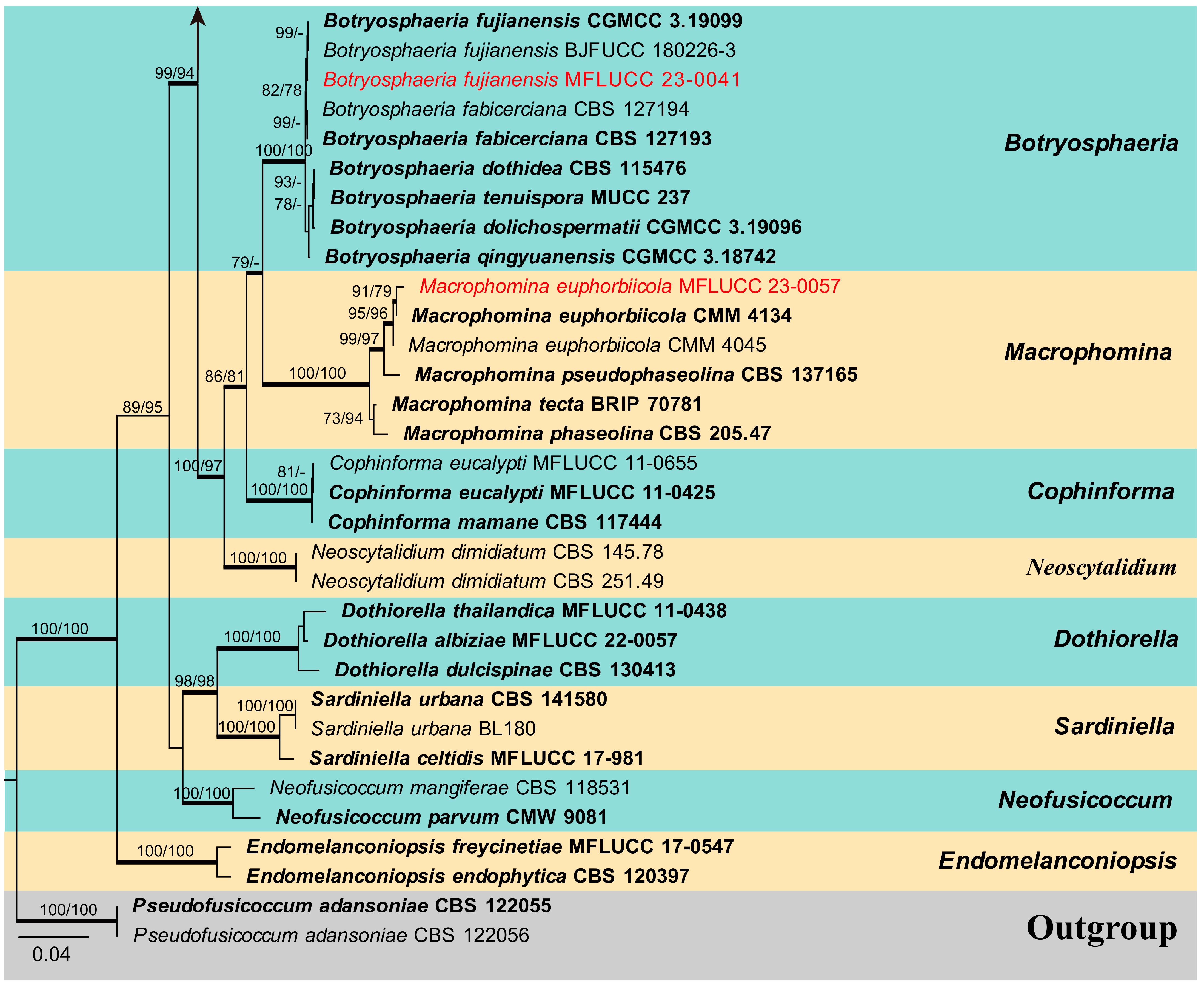
The combined ITS, LSU, tef1-α, and tub2 sequence dataset of Botryosphaeriaceae analysis comprises 89 taxa, including two outgroup taxa. The aligned dataset comprised 2241 characters (ITS: 1–540; LSU: 541–1394; tef1-α: 1395–1793; tub2: 1794–2241) including gaps. The maximum parsimonious dataset consisted of 2241 variable characters, of which 1470 were constant, 655 were parsimony-informative, and 115 were parsimony-uninformative. The MP analysis resulted in a tree length of 2775 steps [consistency index (CI) = 0.444, retention index (RI) = 0.843, relative consistency index (RC) = 0.374, homoplasy index (HI) = 0.556]. The RAxML analysis of the combined data set yielded a best-scoring tree (Figure 1) with a final ML optimization likelihood value of −17,381.405163. The matrix had 1017 distinct alignment patterns, with 25.04% of undetermined characters or gaps. Estimated base frequencies were: A = 0.218861, C = 0.278646, G = 0.277782, T = 0.224710; substitution rates AC = 1.166395, AG = 2.628375, AT = 1.104924, CG = 1.478465, CT = 4.949967, GT = 1.000000; gamma distribution shape parameter (alpha) = 0.207919. The maximum likelihood (ML), maximum parsimony (MP), and Bayesian methods (BI) for phylogenetic analyses resulted in trees with similar topologies. According to the results of phylogenetic analysis, 16 isolates obtained in this study were grouped into 11 clades and located in the genera Botryosphaeria, Diplodia, Dothiorella, Lasiodiplodia, Macrophomina and Sphaeropsis (Figure 1).
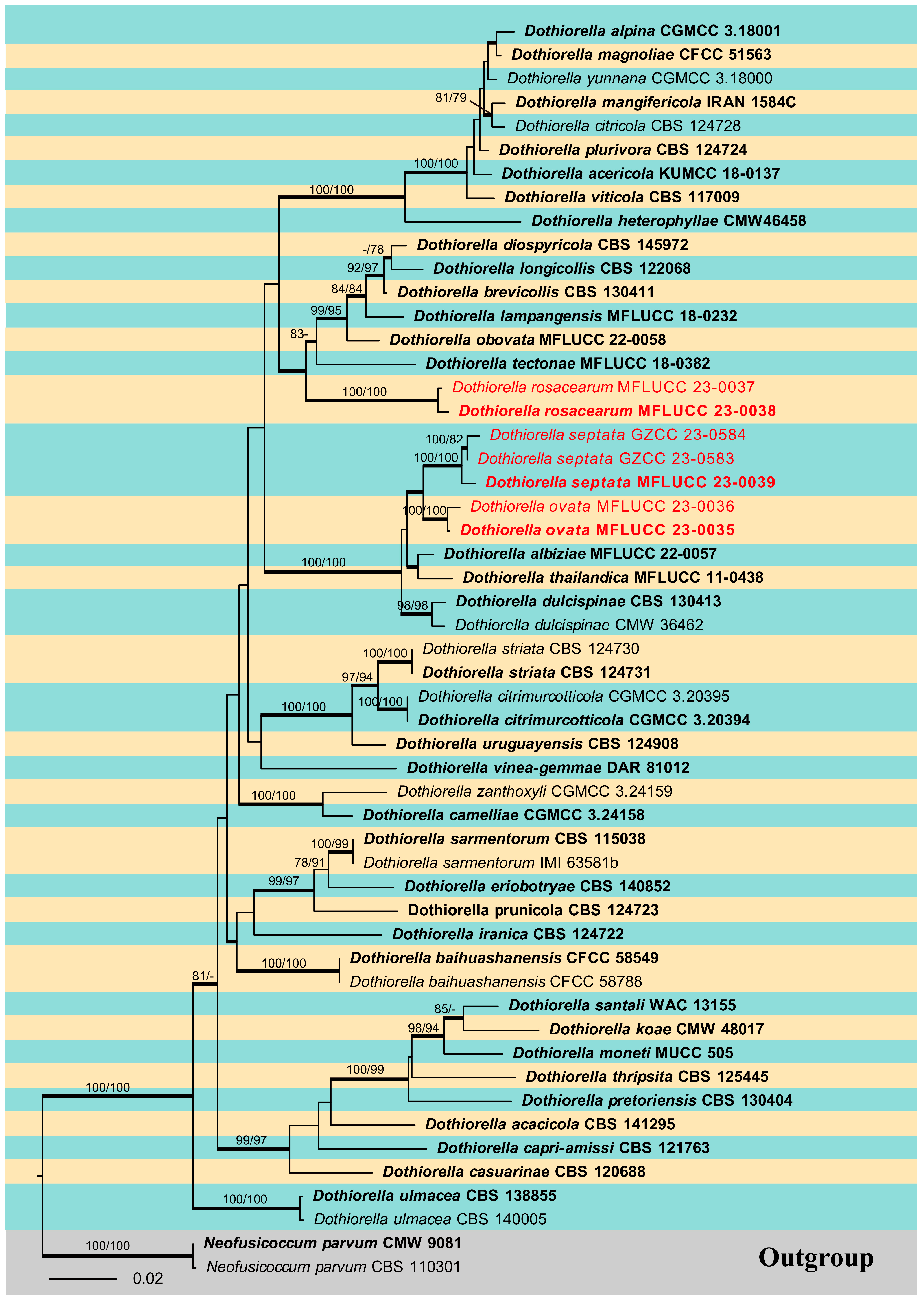
The phylogenetic analysis for the genus Dothiorella was carried out, and a phylogenetic tree combining ITS, LSU, tef1-α, and tub2 sequence data was also constructed (Figure 2). This dataset included 53 ingroup taxa and two outgroup taxa. The aligned dataset comprised 2062 characters (ITS: 1–514; LSU: 515–1331; tef1-α: 1332–1630; tub2: 1631–2062) including gaps. The maximum parsimonious dataset consisted of 2066 variable characters, of which 1637 were constant, 328 were parsimony-informative, and 101 were parsimony-uninformative. The MP analysis resulted in a tree length of 1223 steps [consistency index (CI) = 0.517, retention index (RI) = 0.760, relative consistency index (RC) = 0.393, homoplasy index (HI) = 0.483]. In the ML analyses, the best-scoring RAxML tree with a final likelihood value of −9705.573604 is presented. The matrix had 606 distinct alignment patterns, with 25.62% of undetermined characters or gaps. Estimated base frequencies were: A = 0.215885, C = 0.284638, G = 0.269261, T = 0.230216; substitution rates AC = 1.152119, AG = 2.141304, AT = 1.169569, CG = 1.149449, CT = 5.009593, GT = 1.000000; gamma distribution shape parameter (alpha) = 0.143638. The maximum likelihood (ML), maximum parsimony (MP), and Bayesian methods (BI) for phylogenetic analyses resulted in trees with similar topologies, and the result of ML analysis is shown in Figure 2. Phylogenetic results showed that the isolates obtained in this study were nested within the genus Dothiorella and grouped into three clades as distinct species.
3.2. Taxonomy
Index Fungorum number: IF826938.
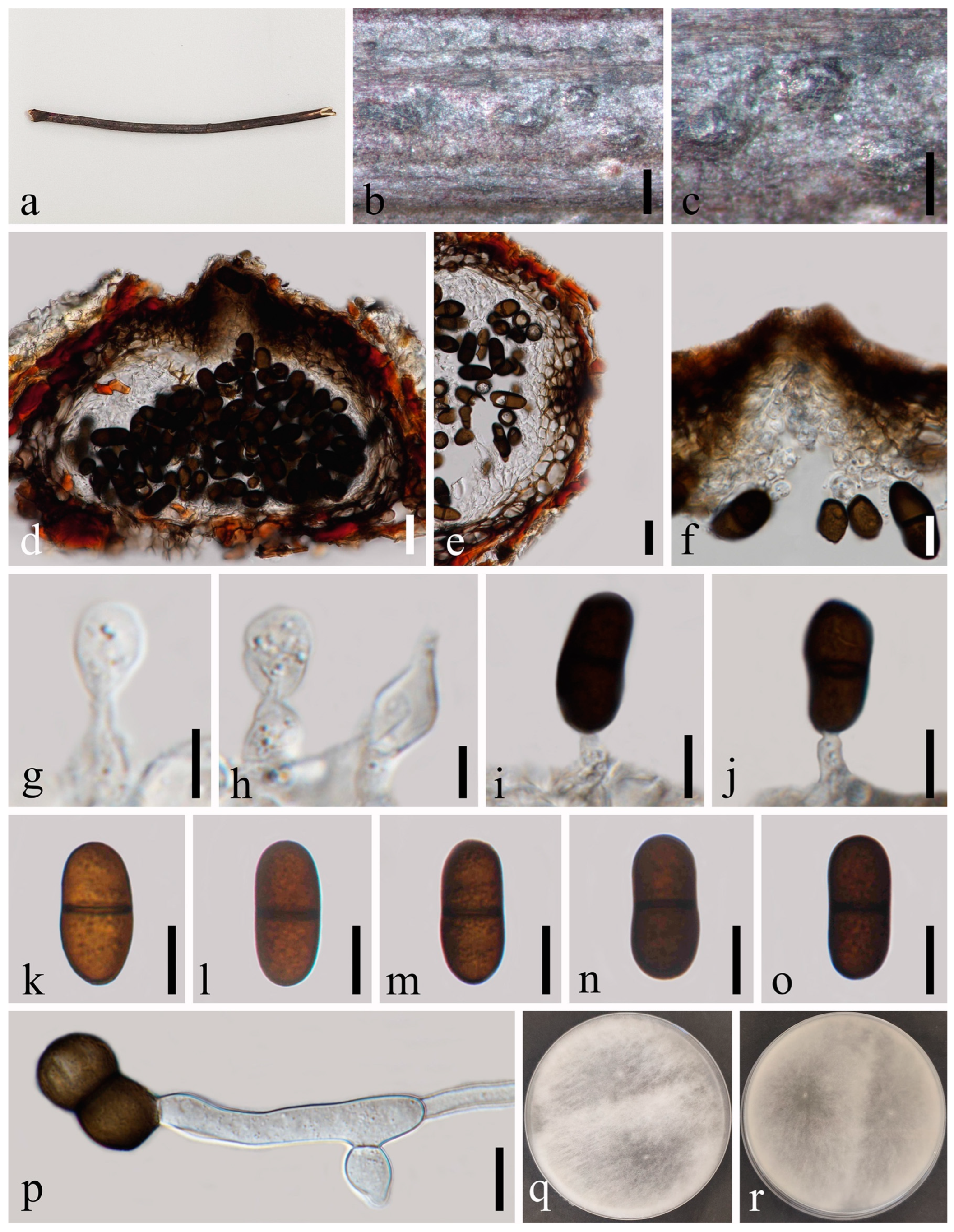
Saprobic on twigs of Tectona grandis L.F. Sexual morph: Ascomata 140–216 × 178–278 μm ( = 182 × 211 μm, n = 20), well-developed, bursting through the bark, generally emerged from the host surface, scattered or gregarious, black, subglobose to obpyriform. Peridium 16–56 μm wide, composed of five strata; an outer stratum has black, thick-walled cells, and the middle and inner layers have black, thin-walled cells. Hamathecium comprising up to 3–5 μm wide, dense, cellular pseudoparaphyses, anastomosing above and between the asci. Asci 76–125 × 20–33 μm (n = 20), bitunicate, eight-spored, broadly clavate, apically rounded and with a visible ocular chamber. Ascospores 20–27 × 7–11 μm ( = 24 × 10 μm, n = 50), initially rhomboid, hyaline, aseptate, elliptical to ovoid, thick-walled, smooth at maturity, apiculus at either end. Asexual morph: Not observed.
Culture characteristics: Ascospores germinating on PDA within 12 h. Colonies are fast growing on PDA, reaching 60 mm diam. after 7 d at 20–25 °C. Circular, white in first few days, becoming pale grey from the center after two weeks, and finally black after three weeks, felt-like, flattened, surface smooth.
Material examined: Thailand, Chiang Mai Province, Amphoe Mae Taeng, Tambon Pa Pae, 19°06′32.6″ N, 98°44′21.1″ E, on dead twigs of Tectona grandis (Lamiaceae), 8 August 2019, Na Wu, YW246 (MFLU 23-0012), living culture MFLUCC 23-0041.
Notes: The phylogenetic results (Figure 1) showed that our newly obtained isolate clustered together with Botryosphaeria fujianensis, and we identified it as B. fujianensis. There was only the asexual morph provided when Chu et al. [37] described this species, and we illustrate the sexual morph of B. fujianensis and reported it as the new record from Thailand in this study.
Index Fungorum number: IF201741; Facesoffungi number: FoF00147.
Saprobic on dead twigs of Prunus persica L. Sexual morph: Not observed. Asexual morph: Conidiomata 213–306 × 247–313 μm ( = 266 × 282 μm, n = 20), semi-immersed or immersed in the substrate, dark brown to black, solitary, globose to ovoid, centrally ostiolate. Ostiole 28–45 μm diam., centrally located, short papillate. Peridium up to 53–78 μm wide, 5–7 layers, consisting of brown and small-celled textura angularis. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 8–14 × 3–7 μm ( = 9 × 4 μm, n = 20), hyaline, cylindrical, discrete, smooth-walled, slightly swollen at the base, forming a single conidium at the tip. Conidia 23–29 × 12–15 μm ( = 27 × 14 μm, n = 50), hyaline, aseptate, externally smooth, internally verruculose, thick-walled, oblong to ovoid, straight, both ends broadly rounded.
Culture characteristics: Conidia germinating on PDA within 24 h. Colonies are fast growing at 20–25 °C, becoming ash-grey on the surface after 7 days and finally black after two weeks, felt-like, dense, convex with the papillate surface, aerial.
Material examined: China, Sichuan Province, Chengdu City, High Tech West Zone, Xi Yuan Avenue, University of Electronic Science and Technology of China campus, 30°45’9″ N, 103°55’31″ E, on dead twigs of Prunus persica L., 22 November 2020, H.Z. Du, YW290 (GZAAS 23-0582), living culture GZCC 23-0578.
Notes: We identified our isolate as Diplodia mutila based on morphology and phylogeny. This is the first record of Di. mutila found on Prunus persica in China.
Index Fungorum number: IF180468.
Saprobic on dead twigs of Wisteria sinensis L. Sexual morph: Not observed. Asexual morph: Conidiomata 138–245 × 206–240 μm ( = 175 × 228 μm, n = 20), semi-immersed or immersed in the substrate, dark brown to black, solitary, pyriform, initially intraepidermal, visible as black dots on the host when mature, thick-walled, glabrous, centrally ostiolate. Ostiole 16–30 μm diam., centrally located, short papillate. Peridium up to 12–30 μm wide, 3–5 layers, consisting of brown and small-celled textura angularis, becoming hyaline towards the inner region. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 6–13 × 3–5 μm ( = 8 × 4 μm, n = 20), hyaline, cylindrical, discrete, smooth-walled, slightly swollen at the base, forming a single conidium at the tip. Conidia 18–27 × 9–13 μm ( = 22 × 11 μm, n = 50), brown, aseptate, externally smooth, internally verruculose, subcylindrical to ellipsoid, moderately thick-walled, ends rounded, often with a truncate base.
Culture characteristics: Conidia germinating on PDA within 24 h. Colonies are fast growing at 20–25 °C, becoming ash-grey on the surface after 7 days, and finally black after two weeks, velvety and floccose, dense, convex with papillate surface, aerial.
Material examined: China, Sichuan Province, Chengdu City, High Tech West Zone, Xi Yuan Avenue, University of Electronic Science and Technology of China campus, 30°45’9″ N, 103°55’31″ E, on dead twigs of Wisteria sinensis L., 22 November 2020, H.Z. Du, YW297 (GZAAS 23-0583), living culture GZCC 23-0579.
Notes: The sexual morph of Diplodia seriata is linked to Botryosphaeria obtusa Shoemaker [40]. We identified our isolate as Di. seriata based on morphology and phylogeny. This is the first record of Di. seriata found on Wisteria sinensis in China.
Dothiorella ovata N. Wu, A.J. Dissanayake & Jian K. Liu sp. nov. Figure 6.
Index Fungorum number: IF900578; Facesoffungi number: FoF14258.
Etymology: In reference to the ovoid conidia.
Holotype: MFLU 23-0009.
Saprobic on a dead wood. Sexual morph: Not observed. Asexual morph: Conidiomata 119–173 × 160–273 μm ( = 141 × 204 μm, n = 20), semi-immersed or immersed in the substrate, emerging through the epidermis when mature, globose to subglobose, pyriform, dark brown, unilocular, solitary, glabrous, ostiolate. Ostiole 22–26 μm diam., single, circular, papillate, centrally located. Peridium up to 16–38 μm wide, comprising host and fungal tissues, thick-walled, dark brown to hyaline cells of textura angularis. Paraphyses absent. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 4–11 × 3–4 μm ( = 7 × 4 μm, n = 20), hyaline, phialidic, cylindrical, straight or curved, smooth-walled. Conidia 20–26 × 9–11 μm ( = 22 × 10 μm, n = 50), hyaline, aseptate, ovoid, rounded at both ends or sometimes with truncate bases, becoming pigmented brown and one septate at maturation, constricted in the middle, smooth-walled, without longitudinal striations or mucilaginous sheath.
Culture characteristics: Conidia germinating on PDA within 12 h. Colonies are fast growing on PDA, reaching 90 mm diam. after 5–6 days at 20–23 °C. Sparse, aerial, filamentous, white in first few days, after 2 weeks, becoming black.
Material examined: Thailand, Chiang Mai Province, Amphoe Mae Taeng, Tambon Cho Lae, 19°08’01.3″ N, 99°00’29.4″ E, on an unidentified dead wood, 6 August 2019, Na Wu, YW177 (MFLU 23-0009, holotype); ex-type living culture MFLUCC 23-0035; ibid., 7 August 2019, Na Wu, YW231 (MFLU 23-0010, paratype), living culture MFLUCC 23-0036.
Notes: Dothiorella ovata is nested in between Do. albiziae, Do. septata and Do. thailandica but can be recognized as a distinct lineage (Figure 2). Morphologically, Do. ovata is similar to Do. septata and Do. albiziae, which in having oblong to ovoid, hyaline conidia becoming brown and one septate at maturation. However, Do. ovata differs from Do. septata and Do. albiziae by its slightly constricted septum and the larger conidia. In addition, a comparison of tef1-α sequences data of Do. ovata and Do. albiziae showed that there are 14 bp (base pair) differences (of 252 bp including the gaps), while Do. ovata and Do. thailandica showed 18 bp differences (of 302 bp including the gaps). Therefore, we introduce Do. ovata as a new species.
Dothiorella rosacearum N. Wu, A.J. Dissanayake & Jian K. Liu, sp. nov. Figure 7.
Index Fungorum number: IF900579; Facesoffungi number: FoF14259.
Etymology: Referring to the host family Rosaceae on which the type specimen was collected.
Holotype: MFLU 23-0014.
Saprobic on dead twigs of Amygdalus sp. (Rosaceae). Sexual morph: Not observed. Asexual morph: Conidiomata 153–234 × 222–336 μm ( = 187 × 270 μm, n = 20), immersed, scattered, dark brown to black, globose or subglobose, solitary, initially intraepidermal, visible as black dots on the host when mature, thick-walled, glabrous. Ostiole 30–32 μm diam., single, straight, sometimes bent, centrally located. Peridium up to 31–45 μm wide, comprising host and fungal tissues, dark brown to black, 5–10 cell layers of textura angularis, becoming thin-walled and hyaline towards the inner region. Paraphyses absent. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 4–9 × 3–5 μm ( = 7 × 4 μm, n = 20), hyaline, aseptate, contents granular, cylindrical to ellipsoidal, smooth-walled, unbranched or occasionally branched, with swollen bases. Conidia 17–21 × 7–10 μm ( = 18 × 8 μm, n = 50), hyaline, aseptate, contents granular, smooth, thin-walled, oval, straight, rounded at the apex, or sometimes with truncate bases, becoming brown and septate when aged, without longitudinal striations or mucilaginous sheath.
Culture characteristics: Conidia germinating on PDA within 12 h. Colonies are fast growing on PDA, reaching 90 mm diam. after 5–6 days at 20–23 °C. Sparse, aerial, filamentous, becoming dark brown to black after 2 weeks.
Material examined: Thailand, Chiang Mai Province, Amphoe Mae Taeng, Tambon Pa Pae, 19°06’32.6″ N, 98°44’21.1″ E, on dead twigs of Amygdalus sp. (Rosaceae), 8 August 2019, Na Wu, YW255 (MFLU 23-0014, holotype); ex-type living culture MFLUCC 23-0038; ibid., on an unidentified decaying wood, 7 August 2019, Na Wu, YW253 (MFLU 23-0013, paratype), living culture MFLUCC 23-0037.
Notes: The phylogenetic result (Figure 2) showed that two isolates of Dothiorella rosacearum constitute a distinct lineage but claded closer to Do. brevicollis, Do. diospyricola, Do. lampangensis, Do. longicollis, Do. obovata and Do. tectonae. A comparison of ITS sequences data between Do. rosacearum and Do. tectonae showed that there are 19 bp of 539 base pairs differences (including the gaps). In addition, the shortly raised irregular striations can be found on the conidia of Do. tectonae, while no striations were observed in Do. rosacearum. Therefore, Do. rosacearum is a morphologically and phylogenetically distinct species and herein introduced as a new species.
Dothiorella septata N. Wu, A.J. Dissanayake & Jian K. Liu, sp. nov. Figure 8.
Index Fungorum number: IF900580; Facesoffungi number: FoF14260.
Etymology: The epithet “septata” refers to the septum observed in mature conidia.
Holotype: MFLU 23-0007.
Saprobic on an unidentified dead wood. Sexual morph: Not observed. Asexual morph: Conidiomata 114–139 × 150–198 μm ( = 126 × 172 μm, n = 20), pyriform or subglobose, immersing through the host epidermis, unilocular, glabrous, thick-walled, ostiolate. Ostiole 19–25 μm diam., single, straight, centrally located. Peridium up to 16–303 μm wide, with outer 3–5 layers of brown cells of textura angularis and inner 1–2 layers of hyaline cells of textura angularis. Paraphyses absent. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 5–10 × 2–5 μm ( = 7 × 3 μm, n = 20), hyaline, phialidic, subcylindrical, smooth-walled. Conidia 19–21 × 8–10 μm ( = 21 × 9 μm, n = 50), oblong to ovoid with a broadly rounded apex, initially hyaline to yellowish and aseptate, becoming brown to dark brown and one septate at maturation, slightly constricted at the septum, smooth-walled, without a mucilaginous sheath.
Culture characteristics: Conidia germinating on PDA within 12 h. Colonies are fast growing on PDA, reaching 90 mm diam. after 5–6 days at 20–23 °C. Sparse, aerial, filamentous, smooth with a crenate edge, white in first few days, becoming grey after one week, and after 2 weeks, becoming black.
Material examined: Thailand, Chiang Mai Province, Amphoe Mae Taeng, Tambon Sop Poeng, 19°07’52.3″ N, 98°45’35.7″ E, on an unidentified dead wood, 9 August 2019, Na Wu, YW173 (MFLU 23-0007, holotype); ex-type living culture MFLUCC 23-0039; ibid., on a decaying wood in a mountain, 7 August 2019, Na Wu, YW217 (GZAAS 23-0587, paratype), living culture GZCC 23-0583; ibid., YW228 (GZAAS 23-0588, paratype), living culture GZCC 23-0584.
Notes: The phylogenetic results (Figure 2) showed that our isolates clustered with Do. ovata and formed a sister group. A comparison of ITS and tef1-α nucleotides shows that Do. septata is significantly different from its sister species, Do. ovata by 7/569 bp (1.2%) in ITS and 13/303 bp (4.3%) in tef1-α. In the phylogenetic analysis, these two species formed two distinct clades in Dothiorella. Morphologically, there are several differences in conidial morphology between these two species. Considering the morpho-molecular data, we introduced Do. septata as a new species.
Index Fungorum number: IF500235.
Saprobic on an unidentified dead wood. Sexual morph: Not observed. Asexual morph: Conidiomata 151–191 × 178–202 μm ( = 171 × 188 μm, n = 20), semi-immersed or immersed in the substrate, solitary, gregarious or confluent, globose to subglobose, centrally ostiolate. Ostiole 21–33 μm diam., centrally located, papillate. Peridium up to 19–46 μm wide, consisting of black and small-celled textura angularis. Paraphyses 2–3 μm wide, hyaline, cylindrical, aseptate, not branched, rounded at apex. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 7–12 ×4–7 μm ( = 9 × 5 μm, n = 20), hyaline, cylindrical. Conidia 27–33 × 14–17 μm ( = 30 × 16 μm, n = 50), hyaline, aseptate, ellipsoid to ovoid, thick-walled, without longitudinal striations or mucilaginous sheath.
Culture characteristics: Conidia germinating on PDA within 24 h. Colonies are fast growing on PDA at 20–25 °C, becoming ash-grey on the surface after 7 days, the reverse is pale grey to grey, and finally black after two weeks, felt-like, sparse, aerial, surface smooth with a crenate edge, filamentous.
Material examined: Thailand, Chiang Mai, Amphoe Mae Taeng, Tambon Cho Lae, 19°08’01.3″ N, 99°00’29.4″ E, on an unidentified dead wood, 6 August 2019, Na Wu, YW191 (MFLU 23-0011), living culture MFLUCC 23-0060.
Notes: The morphology of our collection obtained from decaying woody is similar to the original description of Lasiodiplodia crassispora [41]. In the multi-gene phylogenetic analysis, our new collection clustered with the ex-type strain of L. crassispora (CBS 118741) with strong bootstrap support, and we identified it as L. crassispora.
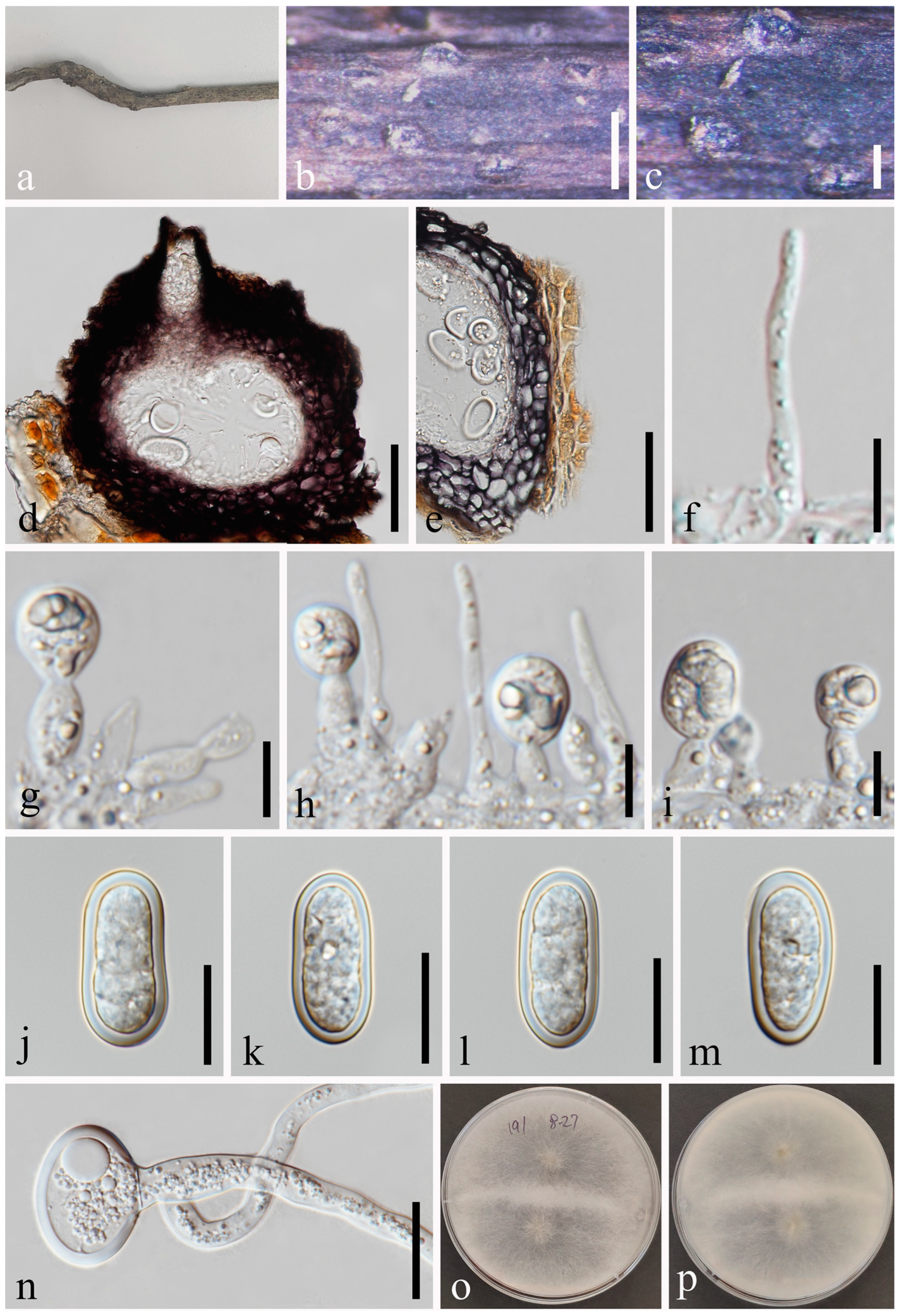
Lasiodiplodia delonicis N. Wu, A.J. Dissanayake & Jian K. Liu, sp. nov. Figure 10.
Index Fungorum number: IF900581; Facesoffungi number: FoF14261.
Etymology: Referring to the host genus on which the fungus was collected, Delonix regia (Fabaceae).
Holotype: MFLU 23-0005.
Saprobic on a fallen pod of Delonix regia L. Sexual morph: Not observed. Asexual morph: Conidiomata 110–180 × 124–171 μm ( = 139 × 151 μm, n = 20), pyriform, immersed to semi-immersed, solitary, black, ostiolate. Ostiole 20–31 μm diam., central, cylindrical to subcylindrical. Peridium up to 18–35 μm wide, with outer 3–4 layers of brown cells of textura angularis and inner 1–2 layers of hyaline cells of textura angularis. Paraphyses 2–3 μm wide, hyaline, cylindrical, aseptate, not branched. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 4–16 × 4–6 μm ( = 8 × 5 μm, n = 20), hyaline, cylindrical, sometimes slightly curved. Conidia 26–38 × 13–29 μm ( = 32 × 17 μm, n = 50), ellipsoid to ovoid, hyaline, aseptate, thick-walled with granular content, occasionally truncate at base, without longitudinal striations or mucilaginous sheath.
Culture characteristics: Conidia germinating on PDA within 24 h. Colonies are fast growing on PDA, reaching 90 mm diam. after 5 days at 20–25 °C, becoming ash-grey on the surface after one week, with the reverse side of the colonies pale grey to grey, and finally black after two weeks, felt-like, sparse, aerial, surface smooth with crenate edge, filamentous.
Material examined: Thailand, Chiang Rai Province, Amphoe Mueang, Tambon Nang Lae, 20°02’22.7″ N, 99°53’38.1″ E, on a fallen pod of Delonix regia, 17 July 2019, Na Wu, YW111 (MFLU 23-0005, holotype); ex-type living culture MFLUCC 23-0058.
Notes: The phylogenetic tree based on ITS, LSU, tef1-α, and tub2 sequence data showed that the new species Lasiodiplodia delonicis (Figure 1) is supported by an absolute bootstrap support (ML/MP/BI = 100/100/1.0). Morphologically, L. delonicis is distinct from other Lasiodiplodia species by its thicker conidial wall and larger conidia. Additionally, conidia of L. delonicis are hyaline throughout the life cycle.
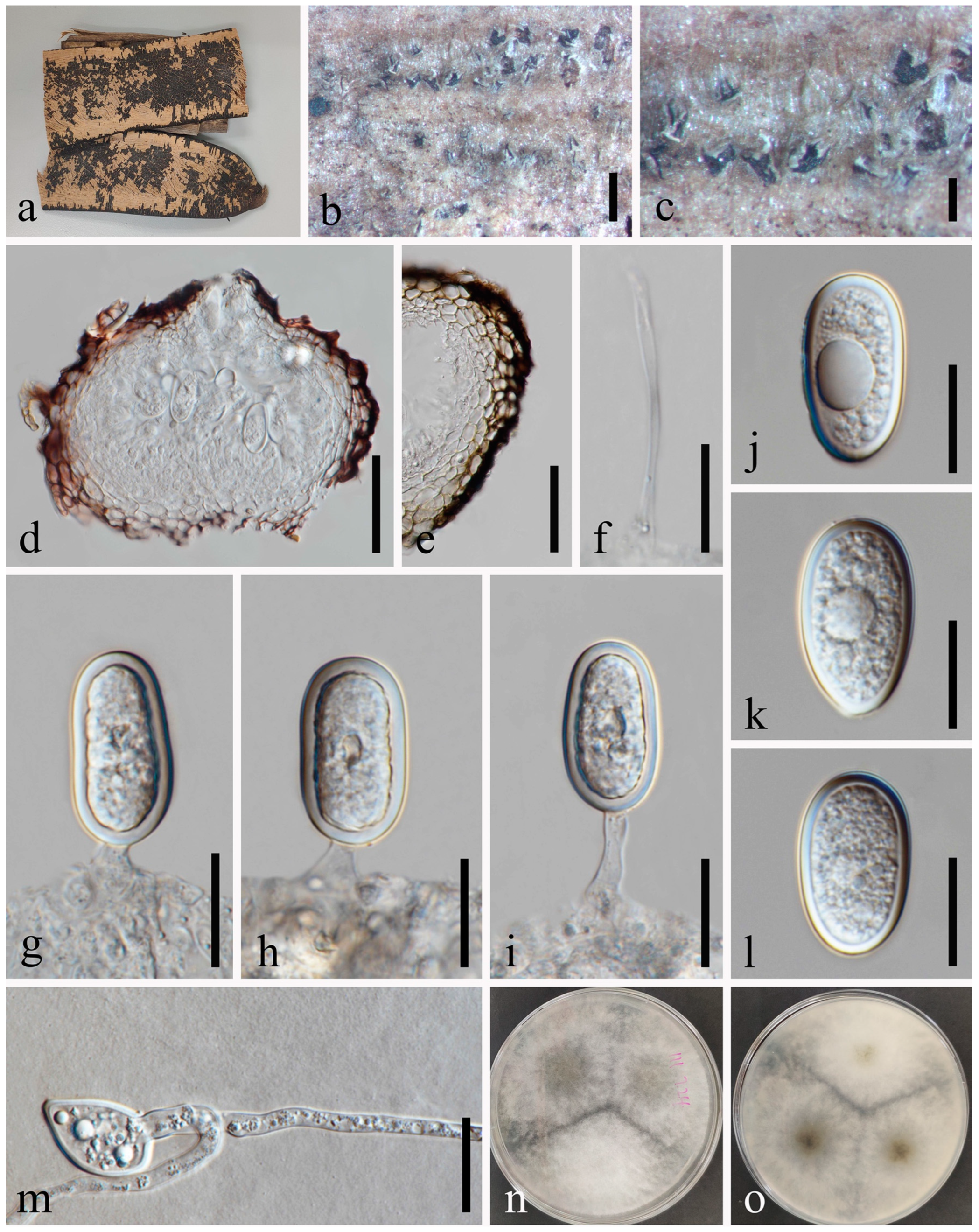
Index Fungorum number: IF514012.
Saprobic on dead seeds of Dipterocarpus retusus L. Sexual morph: Not observed. Asexual morph: Conidiomata 127–164 × 133–192 μm ( = 148 × 160 μm, n = 20), solitary or compound, superficial or immersed, unilocular or multilocular, globose to subglobose, thick-walled, glabrous, ostiolate. Ostiole 26–33 μm diam., single, long, cylindrical to subcylindrical, eccentric. Peridium up to 14–23 μm wide, consisting of brown and small-celled textura angularis. Paraphyses 2–4 μm wide, hyaline, cylindrical, aseptate, not branched, rounded at apex. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 6–13 × 4–5 μm ( = 8 × 5 μm, n = 20), hyaline, cylindrical, proliferating percurrently to form a periclinal thickening. Conidia 24–31 × 14–18 μm ( = 27 × 16 μm, n = 50), initially aseptate, hyaline, ellipsoid to ovoid, thick-walled with granular content, rounded at apex, occasionally truncate at the base, one septate at maturation, without longitudinal striations or mucilaginous sheath.
Culture characteristics: Conidia germinating on PDA within 24 h. Colonies are fast growing on PDA at 20–25 °C, becoming ash-grey on the surface after 7 days, with the reverse side of the colonies pale grey to grey, and finally black after two weeks, felt-like, sparse, aerial, surface smooth with crenate edge, filamentous.
Material examined: Thailand, Chiang Mai Province, Amphoe Mae Taeng, Tambon Cho Lae, 19°08’01.3″ N, 99°00’29.4″ E, on dead seeds of Dipterocarpus retusus L., 10 August 2019, Na Wu, YW151 (MFLU 23-0006), living culture MFLUCC 23-0059.
Notes: In the phylogenetic tree, an isolate obtained in this study (MFLUCC 23-0059) grouped with Lasiodiplodia mahajangana (Figure 1) (ML/MP/BI = 94/95/1.0). Our sample is morphologically similar to L. mahajangana as of the report by Begoude et al. [42], having hyaline, aseptate, ellipsoid to ovoid, thick-walled conidia, which becomes one septate after maturation. We identified our collection as L. mahajangana based on morphology and phylogeny.
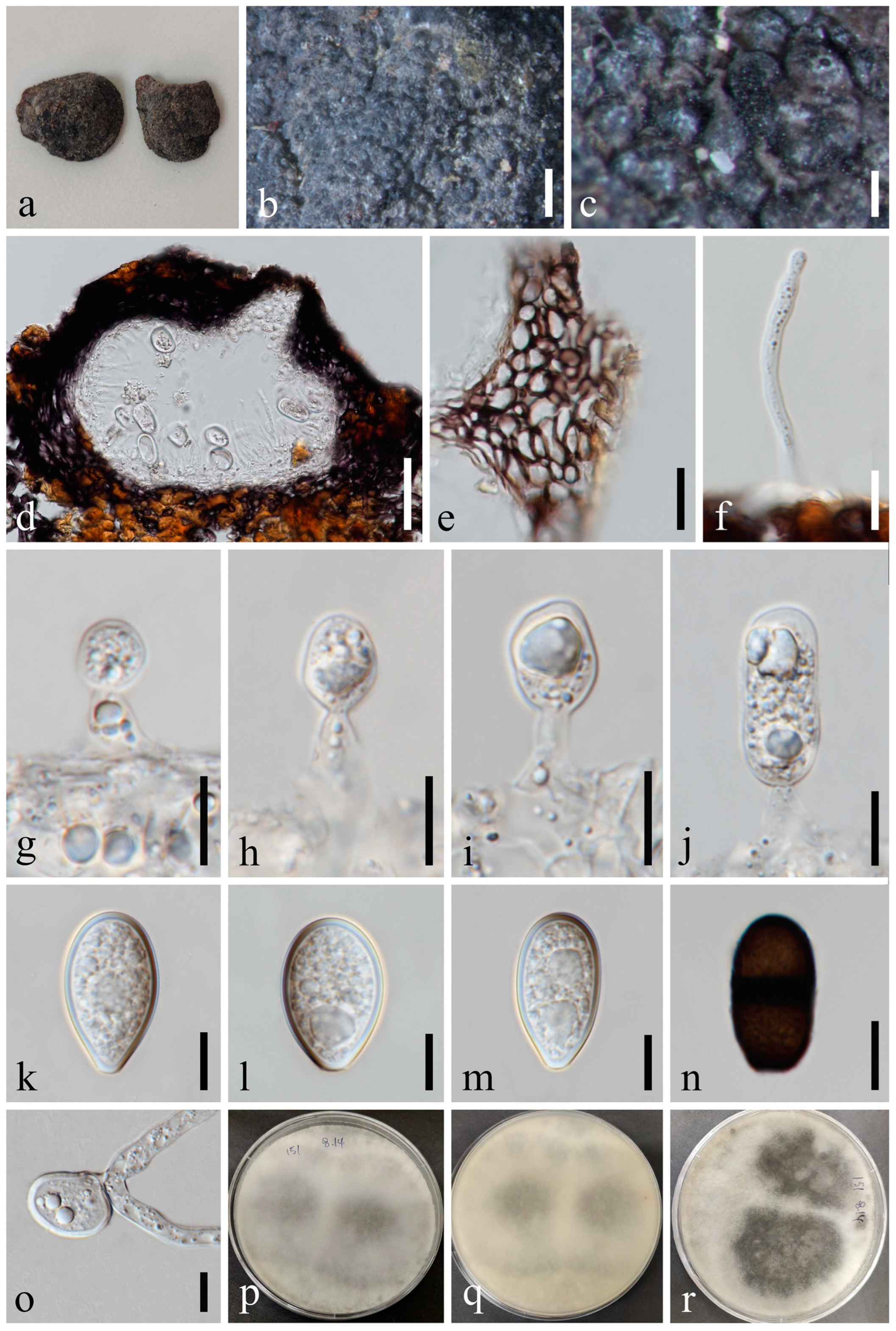
Macrophomina euphorbiicola A.R. Machado, D.J. Soares & O.L. Pereira, Eur. J. Pl. Path. 153: 96 [43] Figure 12.
Index Fungorum number: IF815562.
Saprobic on dead seeds of Plukenetia volubilis L. Sexual morph: Not observed. Asexual morph: Conidiomata 116–172 × 130–161 μm ( = 137 × 149 μm, n = 20), circular, dark brown to black, solitary or gregarious, immersed through the epidermis, visible as black dots or papilla on the host, glabrous. Peridium up to 11–24 μm wide, composed of dark brown to black thick-walled textura angularis, becoming thin-walled and hyaline towards the inner region. Ostiole 12–32 μm diam., cylindrical, short, straight, centrally or laterally located. Paraphyses absent. Conidiophores subcylindrical to ampulliform, reduced to conidiogenous cells. Conidiogenous cells 6–14 × 2–5 μm ( = 9 × 3 μm, n = 20), terminal, hyaline, cylindrical to ellipsoidal, smooth-walled. Conidia 22–26 × 8–11 μm ( = 23 × 9 μm, n = 50), hyaline, oblong to cylindrical, with rounded apex, and narrow, straight, frequently constricted in the middle, aseptate, contents granular, thick- and smooth-walled, bearing octagonal beard-shaped appendages, or widely flared or irregular, undulate, mucoid apical appendage, basal appendages absent.
Culture characteristics: Conidia germinating on PDA within 12 h with germ tubes produced from the middle or each end. Colonies are fast growing on PDA, reaching 90 mm diam. after 5–6 days at 20–23 °C. Sparse, aerial, filamentous, after 2 weeks, becoming dark brown to black.
Material examined: Thailand, Chiang Rai Province, Thoeng, Tambon Nang Lae, Rai Ruen Rom Organic Farm, 19°39’30.2″ N, 100°09’26.4″ E, on dead seeds of Plukenetia volubilis L., 11 June 2019, Na Wu, YW62 (MFLU 23-0004), living culture MFLUCC 23-0057.
Notes: Macrophomina euphorbiicola was introduced by Machado et al. [43]. Due to the previous cultures failing to sporulate, comparison with the type species was not possible. The phylogenetic analysis showed that our isolate was nested within M. euphorbiicola and claded closer to M. pseudophaseolina (Figure 1). We, thus, identify the new collection as M. euphorbiicola.
Index Fungorum number: IF805464; Facesoffungi number: FoF00169.
Saprobic on dead twigs of Tectona grandis. Sexual morph: Ascomata 186–257 × 345–466 μm ( = 233 × 373 μm, n = 20), well-developed, bursting through the bark, generally strongly emerged from the host surface, scattered or gregarious, black, subglobose to obpyriform, ostiolate. Ostiole central, subconical to flattened and the region between the perithecial necks were occupied by black pseudoparenchymatous tissue. Peridium 31–59 μm wide, composed of three strata; an outer stratum is black, thick-walled cells, middle layer and inner layer, black thin-walled cells. Hamathecium comprising up to 2–5 μm wide, dense, cellular pseudoparaphyses, anastomosing above and between the asci. Asci 86–134 × 22–36 μm ( = 113 × 30 μm, n = 20), bitunicate, eight-spored, broadly clavate, apically rounded with a visible ocular chamber. Ascospores 29–34 × 15–19 μm ( = 32 × 17 μm, n = 50), initially rhomboid, hyaline, aseptate, becoming pigmented, brown to dark brown, elliptical to ovoid, thick-walled, smooth at maturity, with an apiculus at either end. Asexual morph: Not observed.
Culture characteristics: Ascospores germinating on PDA within 24 h. Colonies are fast growing on PDA, reaching 60 mm diam. after 5 d at 20–25 °C. Circular, white in first few days, becoming pale grey from the center after one week, and finally black after two weeks, felt-like, sparse, aerial, surface smooth with a crenate edge, filamentous.
Material examined: Thailand, Chiang Mai Province, Amphoe Mae Taeng, Tambon Cho Lae, 19°08’01.3″ N, 99°00’29.4″ E, on dead twigs of Tectona grandis (Lamiaceae), 6 August 2019, Na Wu, YW174 (MFLU 23-0008), living culture MFLUCC 23-0040; ibid., 7 August 2019, Na Wu, YW213 (GZAAS 23-0589), living culture GZCC 23-0589.
Notes: The phylogenetic results (Figure 1) showed that our newly obtained isolate clustered together with Sphaeropsis eucalypticola, and we identified it as S. eucalypticola. This is the first record of S. eucalypticola found on Tectona grandis in Thailand.
4. Discussion
Studies on Botryosphaeriaceae, dealing with the phylogenetic traits and morphology of isolates associated with various hosts, have increased in recent years, enabling the worldwide identification of taxa at the species level [2,5,18,45,46,47,48,49]. In this study, 16 Botryosphaeriaceae isolates were obtained from several decaying woody hosts (dead arial twigs, branches, stems, bark, and seed pods) in southwestern China and northern Thailand, and they were identified as 11 species based on a polyphasic approach of morphological features and molecular phylogeny. These species included Botryosphaeria fujianensis, Diplodia mutila, Di. seriata, Dothiorella ovata, Do. rosacearum, Do. septata, Lasiodiplodia crassispora, L. delonicis, L. mahajangana, Macrophomina euphorbiicola and Sphaeropsis eucalypticola. Of these, Do. ovata, Do. rosacearum, Do. septata and L. delonicis are introduced as novel species, and the remaining seven species were identified as new hosts or new geographical records. All species collected in this study are saprophytic on the host. It should be noted that even though sporulation was induced on sterile toothpicks or pine needles on PDA, the respective asexual morph or sexual morph was not observed. Thus, the fungal identification and classification in this study are based on their morphological characteristics of either asexual or sexual morphs and the phylogenetic analysis results.
Macrophomina and Sphaeropsis are two of the least common genera in the family Botryosphaeriaceae. Five species are validly known in Macrophomina, among which M. phaseolina and M. euphorbiicola were introduced as pathogens [43,50,51,52]. In this study, the asexual morph of M. euphorbiicola was collected from Plukenetia volubilis in northern Thailand. Due to the previous cultures failing to sporulate, the morphology of M. euphorbiicola has not been described [43]. Hence, we provide the first detailed description and illustration of M. euphorbiicola for the first time and also report it as a new record from Plukenetia volubilis in Thailand. Sphaeropsis was typified with S. visci by Saccardo [53] with 632 records in Index Fungorum (Accessed July 2023), and only eight species are recognized with accessible cultures so far [16,44]. In this study, one previously known species, S. eucalypticola, was collected from Tectona grandis in Thailand and reported as a new host record. Sphaeropsis eucalypticola has also been reported on Bauhinia purpurea and Eucalyptus sp. in Thailand [4,54]. Mapook et al. [55] identified S. chromolaenicola from Chromolaena odorata in Thailand. However, the remaining members of the genus have not been found on any host in Thailand.
This study revealed two previously known Diplodia species, Di. mutila and Di. seriata from Sichuan province. It is worth noting that similarly to our collection of Di. mutila (from Prunus persica) and Di. seriata (from Wisteria sinensis), Li et al. [49] also found these two species from dead branches of Camellia oleifera, and another two Diplodia species (Di. acerigena and Di. pistaciicola) in Sichuan province. Diplodia species mainly occur on woody hosts, causing rots, cankers, shoot and tip blight [11,56,57,58,59]. Thus, the discovery and in-depth research of this genus are conducive to the protection of woody plants and the maintenance of greater economic benefits.
Dothiorella was the most frequently isolated genus in this study, as seven Dothiorella isolates were obtained from decaying woody hosts in Chiang Mai Province, Thailand. Dothiorella was introduced by Saccardo [53] with Do. pyrenophora as the type species, and presently, only 38 species are accepted in this genus based on phylogenetic analyses [16,18,49]. Zhang et al. [48] made a systematic revision of Dothiorella by synonymizing 15 known species, which reduced the number of Dothiorella members and established a more stable systematic relationship. Most of the members of the genus Dothiorella were rarely collected in Thailand in the past; however, there have been many reports of Dothiorella species being collected in Thailand in recent years [60,61,62,63]. We speculate that this may be due to random sampling. In this study, three new species Do. ovata, Do. rosacearum and Do. septata are introduced based on morphological features (asexual morphs) and phylogenetic evidence.
Lasiodiplodia was formally established by Clendenin [64] with L. tubericola Ellis and Everhart (=L. theobromae) [4] as the type species. So far, 37 ex-type/isotype/neotype species entries have been accepted and uploaded to the Botryosphaeriales website [16,65,66]. It is worth noting that most of the species were introduced as asexual morphs of Lasiodiplodia, and only a few species of sexual morph have been found in nature, such as L. gonubiensis, L. lignicola and L. theobromae [44,67,68]. The three Lasiodiplodia species collected in this study were all asexual morphs and collected from woody plants. Among them, L. crassispora and L. mahajangana were previously known species, and L. delonicis was introduced as a new species. Lasiodiplodia crassispora was first introduced by Burgess et al. [41] based on distinctive morphological characters and phylogenetic analyses. Zhang et al. [48] synonymized L. pyri under L. crassispora. In this study, L. crassispora was collected from decaying wood. Though the species has been found in several countries, such as Australia, Brazil, Namibia, Senegal, and Venezuela [41,48], this is the first time L. crassispora has been reported in Thailand. We collected L. mahajangana from Dipterocarpus retusus in this study. Zhang et al. [48] synonymized L. caatinguensis, L. curvata, L. exigua, L. irregularis, L. macroconidia, and L. pandanicola under L. mahajangana, thus expanding the host range and geographical distribution of this species. Interestingly, the conidia of L. mahajangana are straight or curved, and its conidia morphology is more special compared with other species of Lasiodiplodia [42,48]. At the same time, this study also collected a new species, L. delonicis, from a fallen pod of Delonix regia. In addition, mature conidia with longitudinal striations of Lasiodiplodia is one of its distinguishing features from Diplodia [44]. However, it has been observed that if the asexual stage is produced on culture, the conidia often have obvious longitudinal striations, while the asexual stage produced in nature has less distinct or absent longitudinal striations [61,68,69,70]. This inference can be found in previous reports and this study. The reason for this phenomenon might be due to the variations in the environment in which the fungi grow. Thus, it is important to collect more fresh specimens to verify this observation.
Botryosphaeria fujianensis was introduced as a pathogen-causing stem canker of blueberry in Fujian province, China [37], whereas our species was isolated from dead twigs of Tectona grandis (Lamiaceae) in Chiang Mai Province, Thailand. As this is the first record of B. fujianensis isolated from Thailand, we suspect that it might be found on more hosts in the future.
With the increased number of studies of Botryosphaeriaceae based on morphology, ecology, and DNA-based phylogeny, more new species and records are constantly being discovered [7,71,72,73,74]. However, there are still many aspects needed to clarify this fungal family, such as specifying species from environmental samples, resolving the opportunistic pathogenic nature, and defining species boundaries. The results of this study indicate that there is still much potential for Botryosphaeriaceae members to be discovered in China and Thailand. As members of the Botryosphaeriaceae family represent a growing threat to agricultural crops and urban and natural forest ecosystems [75,76,77,78], this finding raises questions about the origin, introduction, and pathway of these fungi as well as underlining the need to develop suitable actions to limit their further spread.
Author Contributions
Conceptualization, J.-K.L., N.W. and A.J.D.; methodology, N.W. and H.-Z.D.; formal analysis, N.W. and A.J.D.; resources, N.W.; data curation, N.W.; writing—original draft preparation, N.W., A.J.D. and H.-Z.D.; writing—review and editing, N.W., A.J.D., H.-Z.D. and J.-K.L.; supervision, J.-K.L.; project administration, J.-K.L.; funding acquisition, J.-K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the second Tibetan Plateau Scientific Expedition and Research (STEP) Program (2019QZKK0503) and the National Natural Science Foundation of China (NSFC 31600032).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated for this study can be found in the NCBI database.
Acknowledgments
Na Wu is grateful to Shaun Pennycook for correcting the Latin names of the novel taxa; Ningguo Liu and Chuangen Lin are thanked for their valuable suggestions. Na Wu also would like to thank the Mae Fah Luang University for its financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J.K. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Wulandari, N.F.; To-anun, C.; Hyde, K.D.; Duong, L.M.; de Gruyter, J.; Meffert, J.P.; Groenewald, J.Z.; Crous, P.W. Phyllosticta citriasiana sp. nov., the cause of Citrus tan spot of Citrus maxima in Asia. Fungal Divers. 2009, 34, 23–39. [Google Scholar]
- Liu, J.K.; Phookamsak, R.; Doilom, M.; Wikee, S.; Li, Y.M.; Ariyawansha, H.; Boonmee, S.; Chomnunti, P.; Dai, D.Q.; Bhat, J.D.; et al. Towards a natural classification of Botryosphaeriales. Fungal Divers. 2012, 57, 149–210. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Phillips, A.J.L.; Hyde, K.D.; Li, X.H. Botryosphaeriaceae: Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, S.; Zhao, L.; Sun, X.; He, W.; Zhang, Y.; Dai, Y.C. Lasiodiplodia spp. associated with Aquilaria crassna in Laos. Mycol. Prog. 2019, 18, 683–701. [Google Scholar] [CrossRef]
- Senwanna, C.; Mapook, A.; Samarakoon, M.C.; Karunarathna, A.; Wang, Y.; Tang, A.M.C.; Haituk, S.; Suwannarach, N.; Hyde, K.D.; Cheewangkoon, R. Ascomycetes on Para rubber (Hevea brasiliensis). Mycosphere 2021, 12, 1334–1512. [Google Scholar] [CrossRef]
- De Silva, N.I.; Hyde, K.D.; Lumyong, S.; Phillips, A.J.L.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Tennakoon, D.S.; Suwannarach, N.; Karunarathna, S.C. Morphology, phylogeny, host association and geography of fungi associated with plants of Annonaceae, Apocynaceae and Magnoliaceae. Mycosphere 2022, 13, 955–1076. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Li, X.; Zhang, W.; Hyde, K.D.; Yan, J. Trail of decryption of molecular research on Botryosphaeriaceae in woody plants. Phytopathol. Mediterr. 2016, 55, 147–171. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Hyde, K.D.; Chethana, K.W.T.; Zhang, W.; Zhao, W.S.; Yan, J.Y.; Li, X.H. Mycosphere Essays 14: Assessing the aggressiveness of plant pathogenic Botryosphaeriaceae. Mycosphere 2016, 7, 883–892. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Mehl, J.W.; Slippers, B.; Roux, J.; Wingfield, M.J. Overlap of latent pathogens in the Botryosphaeriaceae on a native and agricultural host. Fungal Biol. 2017, 121, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Theissen, F.; Sydow, H. Vorentwürfe zu den Pseudosphaeriales. Ann. Mycol. 1918, 16, 1–34. [Google Scholar]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CABI Publishing: Wallingford, UK, 2008; ISBN 978-0-85199-826-8. [Google Scholar]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Luecking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Wu, N.; Dissanayake, A.J.; Manawasinghe, I.S.; Rathnayaka, A.R.; Liu, J.K.; Phillips, A.J.L.; Promputtha, I.; Hyde, K.D. https://botryosphaeriales.org/, an up-to-date classification and account of taxa of Botryosphaeriales. Database 2021, 2021, baab061. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoðdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Lin, L.; Bai, Y.K.; Pan, M.; Tian, C.M.; Fan, X.L. Morphology and molecular analyses reveal three new species of Botryosphaeriales isolated from diseased plant branches in China. MycoKeys 2023, 97, 1–19. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E.; Jones, E.B.G.; Moslem, M.; Hyde, K.D. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute & British Mycological Society: Kew, UK, 1970. [Google Scholar]
- Chethana, K.W.T.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them? Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Pem, D.; Jeewon, R.; Chethana, K.W.T.; Hongsanan, S.; Doilom, M.; Suwannarach, N.; Hyde, K.D. Species concepts of Dothideomycetes: Classifcation, phylogenetic inconsistencies and taxonomic standardization. Fungal Divers. 2021, 109, 283–319. [Google Scholar] [CrossRef]
- Chaiwan, N.; Gomdola, D.; Wang, S.; Monkai, J.; Tibpromma, S.; Doilom, M.; Wanasinghe, D.N.; Mortimer, P.E.; Lumyong, S.; Hyde, K.D. https://gmsmicrofungi.org: An online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere 2021, 12, 1513–1526. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2653–2677. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest (Version 2.2); Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0 b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Chu, R.T.; Dou, Z.P.; He, W.; Zhang, Y. Two novel species of Botryosphaeria causing stem canker of blueberries from China. Mycosystema 2021, 40, 473–486. [Google Scholar] [CrossRef]
- Montagne, J.F.C. Notice sur les plantes cryptogames récemment découvertes en France contenant aussi l’indication précis des localités de quelques espèces les plus rares de la flore française. Ann. Des Sci. Nat. Bot. 1834, 2, 295–307. [Google Scholar]
- De Notaris, G. Micromycetes Italici novi vel minus cogniti, Decas 4. Mem. Reale Accad. Sci. Torino 1845, 2, 17–30. [Google Scholar]
- Shoemaker, R.A. Conidial states of some Botryosphaeria species on Vitis and Quercus. Can. J. Biochem. 1964, 42, 1297–1303. [Google Scholar] [CrossRef]
- Burgess, T.I.; Barber, P.A.; Mohali, S.; Pegg, G.; De Beer, W.; Wingfield, M.J. Three new Lasiodiplodia spp. from the tropics, recognized based on DNA sequence comparisons and morphology. Mycologia 2006, 98, 423–435. [Google Scholar] [CrossRef]
- Begoude, B.A.D.; Slippers, B.; Wingfield, M.J.; Roux, J. Botryosphaeriaceae associated with Terminalia catappa in Cameroon, South Africa and Madagascar. Mycol. Prog. 2010, 9, 101–123. [Google Scholar] [CrossRef]
- Machado, A.R.; Pinho, D.B.; Soares, D.J.; Gomes, A.A.M.; Pereira, O.L. Bayesian analyses of five gene regions reveal a new phylogenetic species of Macrophomina associated with charcoal rot on oilseed crops in Brazil. Eur. J. Plant Pathol. 2018, 153, 89–100. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Chen, Y.Y.; Cheewangkoon, R.; Liu, J.K. Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China. J. Fungi 2021, 7, 893. [Google Scholar] [CrossRef]
- Garcia, J.F.; Lawrence, D.P.; Morales-Cruz, A.; Travadon, R.; Minio, A.; Hernandez-Martinez, R.; Rolshausen, P.E.; Baumgartner, k.; Cantu, D. Phylogenomics of Plant-Associated Botryosphaeriaceae species. Front. Microbiol. 2021, 12, 652802. [Google Scholar] [CrossRef]
- Xiao, X.E.; Wang, W.; Crous, P.W.; Wang, H.K.; Jiao, C.; Huang, F.; Pu, Z.X.; Zhu, Z.R.; Li, H.Y. Species of Botryosphaeriaceae associated with citrus branch diseases in China. Persoonia 2021, 47, 106–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef] [PubMed]
- Li, W.L.; Liang, R.R.; Dissanayake, A.J.; Liu, J.K. Botryosphaerialean fungi associated with woody oil plants cultivated in Sichuan Province, China. MycoKeys 2023, 97, 71–116. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Suh, S.O.; Schneider, R.W.; Russin, J.S. Host specialization in the charcoal rot fungus, Macrophomina phaseolina. Phytopathology 2001, 91, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Babu, B.K.; Saxena, A.K.; Srivastava, A.K.; Arora, D.K. Identification and detection of Macrophomina phaseolina by using species-specific oligonucleotide primers and probe. Mycologia 2007, 99, 797–803. [Google Scholar] [CrossRef]
- Sarr, M.P.; Ndiaye, M.B.; Groenewald, J.Z.; Crous, P.W. Genetic diversity in Macrophomina phaseolina, the causal agent of charcoal rot. Phytopathol. Mediterr. 2014, 53, 250–268. [Google Scholar] [CrossRef]
- Saccardo, P.A. Conspectus genera fungorum Italiae inferiorum. Michelia 1880, 2, 1–38. [Google Scholar]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Lopes, J.; Abdollahzadeh, J.; Bobev, S.; Alves, A. Resolving the Diplodia complex on apple and other Rosaceae hosts. Persoonia—Mol. Phylogeny Evol. Fungi 2012, 29, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in holm oak decline on Caprera island (Italy). For. Pathol. 2014, 44, 191–200. [Google Scholar] [CrossRef]
- Giambra, S.; Piazza, G.; Alves, A.; Mondello, V.; Berbegal, M.; Armengol Fortí, J.; Burruano, S. Botryosphaeriaceae species associated with diseased loquat trees in Italy and description of Diplodia rosacearum sp. nov. Mycosphere 2016, 7, 978–989. [Google Scholar] [CrossRef]
- Doilom, M.; Shuttleworth, L.A.; Roux, J.; Chukeatirote, E.; Hyde, K.D. Botryosphaeriaceae associated with Tectona grandis (teak) in Northern Thailand. Phytotaxa 2015, 233, 1–26. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Rathnayaka, A.R.; Chethana, K.W.T.; Phillips, A.J.L.; Jones, E.B.G. Two new species of Botryosphaeriaceae (Botryosphaeriales) and new host/geographical records. Phytotaxa 2022, 564, 8–38. [Google Scholar] [CrossRef]
- Rathnayaka, A.R.; Chethana, K.W.T.; Pasouvang, P.; Phillips, A.J.L. Morphology and muti-gene phylogenetic analysis reveals Dothiorella chiangmaiensis sp. nov. (Botryosphaeriaceae, Botryosphaeriales) from Thailand. Curr. Res. Environ. Appl. Mycol. (J. Fungal Biol.) 2022, 12, 322–332. [Google Scholar] [CrossRef]
- Clendenin, I. Lasiodiplodia Ellis. and Everh. n. gen. Bot. Gaz. 1896, 21, 92–93. [Google Scholar] [CrossRef]
- El-Ganainy, S.M.; Ismail, A.M.; Iqbal, Z.; Elshewy, E.S.; Alhudaib, K.A.; Almaghasla, M.I.; Magistà, D. Diversity among Lasiodiplodia Species Causing Dieback, Root Rot and Leaf Spot on Fruit Trees in Egypt, and a Description of Lasiodiplodia newvalleyensis sp. nov. J. Fungi 2022, 8, 1203. [Google Scholar] [CrossRef]
- Jami, F.; Marincowitz, S.; Durán, A.; Slippers, B.; Abad, J.I.; Chen, S.; Wingfield, M.J. Botryosphaeriaceae diversity on Eucalyptus clones in different climate zones of Indonesia. For. Pathol. 2022, 52, e12737. [Google Scholar] [CrossRef]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-Anun, C.; Crous, P.W. Caulicolous Botryosphaeriales from Thailand. Persoonia–Mol. Phylogeny Evol. Fungi 2015, 34, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.P.; He, W.; Zhang, Y. Lasiodiplodia chinensis, a new holomorphic species from China. Mycosphere 2017, 8, 521–532. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Xia, G.Y.; Manawasinghe, I.S.; Phillips, A.J.L.; You, C.P.; Jayawardena, R.S.; Luo, M.; Hyde, K.D. Lasiodiplodia fici sp. nov., Causing Leaf Spot on Ficus altissima in China. Pathogens 2022, 11, 840. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; de Silva, N.I.; Jeewon, R.; Bhat, D.J.; Phookamsak, R.; Doilom, M.; Boonmee, S.; Jayawardena, R.S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Jayawardena, R.S.; Li, H.L.; Zhou, Y.Y.; Zhang, W.; Phillips, A.J.L.; Wanasinghe, D.N.; Dissanayake, A.J.; Li, X.H.; Li, Y.H.; et al. Microfungi associated with Camellia sinensis: A case study of leaf and shoot necrosis on Tea in Fujian, China. Mycosphere 2021, 12, 430–518. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Calabon, M.S.; Jones, E.B.G.; Zhang, Y.X.; Liao, C.F.; Xiong, Y.; Chaiwan, N.; Kularathnage, N.D.; Liu, N.G.; Tang, S.M.; et al. Mycosphere notes 345–386. Mycosphere 2022, 13, 454–557. [Google Scholar] [CrossRef]
- Sun, J.E.; Meng, C.R.; Phillips, A.J.L.; Wang, Y. Two new Botryosphaeria (Botryosphaeriales, Botryosphaeriaceae) species in China. MycoKeys 2022, 94, 1–15. [Google Scholar] [CrossRef]
- Karani, S.; Njuguna, J.; Runo, S.; Muchugi, A.; Machua, J.; Mwaniki, P. Molecular and morphological identification of fungi causing canker and dieback diseases on Vangueria infausta (Burch) subsp. rotundata (Robyns) and Berchemia discolor (Klotzsch) Hemsl in lower Eastern Kenya. Afr. J. Biotechnol. 2022, 21, 6–15. [Google Scholar] [CrossRef]
- Wijesinghe, S.N.; Zucconi, L.; Camporesi, E.; Wanasinghe, D.N.; Boonmee, S.; Samarakoon, M.C.; Chethana, K.W.T.; Puwakpitiya Gedara, C.; Maharachchikumbura, S.S.N.; Yong, W.; et al. An updated account of Fagales-inhabiting Italian Ascomycota and mycogeography, with additions to Pezizomycotina. Asian J. Mycol. 2022, 5, 79–186. [Google Scholar]
- Si, Y.Z.; Sun, J.W.; Wan, Y.; Chen, Y.N.; He, J.; Li, W.Z.; Li, D.W.; Zhu, L.H. Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China. J. Fungi 2023, 9, 404. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Pornsuriya, C.; Sunpapao, A. A New Species of Neoscytalidium hylocereum sp. nov. Causing Canker on Red-Fleshed Dragon Fruit (Hylocereus polyrhizus) in Southern Thailand. J. Fungi 2023, 9, 197. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic tree generated from maximum likelihood (ML) analysis based on combined ITS, LSU, tef1-α, and tub2 sequence data for selected closely related genera within the family Botryosphaeriaceae. Bootstrap values for maximum likelihood (ML) and maximum parsimony (MP) equal to or greater than 75% are given near the nodes. Bayesian posterior probabilities (BYPP) equal to or greater than 0.95 are denoted in thickened branches. The strain numbers are given after the species names, and ex-type strains are indicated in bold. The newly generated isolates of this study are in red. The tree is rooted with two isolates of Pseudofusicoccum adansoniae (CBS 122055, CBS 122056).
Figure 1.
Phylogenetic tree generated from maximum likelihood (ML) analysis based on combined ITS, LSU, tef1-α, and tub2 sequence data for selected closely related genera within the family Botryosphaeriaceae. Bootstrap values for maximum likelihood (ML) and maximum parsimony (MP) equal to or greater than 75% are given near the nodes. Bayesian posterior probabilities (BYPP) equal to or greater than 0.95 are denoted in thickened branches. The strain numbers are given after the species names, and ex-type strains are indicated in bold. The newly generated isolates of this study are in red. The tree is rooted with two isolates of Pseudofusicoccum adansoniae (CBS 122055, CBS 122056).


Figure 2.
Phylogenetic tree generated from the maximum likelihood (ML) analysis based on combined ITS, LSU, tef1-α, and tub2 sequence data of Dothiorella. Bootstrap values for maximum likelihood (ML) and maximum parsimony (MP) equal to or greater than 75% are given near the nodes. Branches with Bayesian posterior probabilities (BYPP) equal to or greater than 0.95 are thickened. The new isolates obtained in this study are indicated in red, and ex-type strains are in bold. The tree is rooted to Neofusicoccum parvum (CBS 110301, CMW 9081).
Figure 2.
Phylogenetic tree generated from the maximum likelihood (ML) analysis based on combined ITS, LSU, tef1-α, and tub2 sequence data of Dothiorella. Bootstrap values for maximum likelihood (ML) and maximum parsimony (MP) equal to or greater than 75% are given near the nodes. Branches with Bayesian posterior probabilities (BYPP) equal to or greater than 0.95 are thickened. The new isolates obtained in this study are indicated in red, and ex-type strains are in bold. The tree is rooted to Neofusicoccum parvum (CBS 110301, CMW 9081).

Figure 3.
Botryosphaeria fujianensis (MFLU 23-0012). (a–c) Ascomata on host substrate. (d) Vertical section of ascomata. (e) Structure of the peridium. (f) Pseudoparaphyses. (g,h) Asci. (i,j) Ascospores. (k) Germinated ascospore. (l–n) Colonies on PDA, above (l,n) and below (m). Scale bars: (d) = 50 μm, (e) = 5 μm, (f–j) = 10 μm, (k) = 50 μm.
Figure 3.
Botryosphaeria fujianensis (MFLU 23-0012). (a–c) Ascomata on host substrate. (d) Vertical section of ascomata. (e) Structure of the peridium. (f) Pseudoparaphyses. (g,h) Asci. (i,j) Ascospores. (k) Germinated ascospore. (l–n) Colonies on PDA, above (l,n) and below (m). Scale bars: (d) = 50 μm, (e) = 5 μm, (f–j) = 10 μm, (k) = 50 μm.

Figure 4.
Diplodia mutila (GZAAS 23-0582). (a–c) Conidiomata on host substrate. (d,e) Vertical section of conidiomata. (f) Section of peridium. (g–i) Conidiogenous cells and developing conidia. (j–m) Conidia. (n,o) Colonies on PDA, above (n) and below (o). Scale bars: (d–f) = 20 μm, (g–m) = 10 μm.
Figure 4.
Diplodia mutila (GZAAS 23-0582). (a–c) Conidiomata on host substrate. (d,e) Vertical section of conidiomata. (f) Section of peridium. (g–i) Conidiogenous cells and developing conidia. (j–m) Conidia. (n,o) Colonies on PDA, above (n) and below (o). Scale bars: (d–f) = 20 μm, (g–m) = 10 μm.

Figure 5.
Diplodia seriata (GZAAS 23-0583). (a–c) Conidiomata on host substrate. (d,e) Vertical section of conidiomata. (f) Section of peridium. (g–i) Conidiogenous cells and developing conidia. (j–m) Conidia. Scale bars: (d–f) = 20 μm, (g–m) = 10 μm.
Figure 5.
Diplodia seriata (GZAAS 23-0583). (a–c) Conidiomata on host substrate. (d,e) Vertical section of conidiomata. (f) Section of peridium. (g–i) Conidiogenous cells and developing conidia. (j–m) Conidia. Scale bars: (d–f) = 20 μm, (g–m) = 10 μm.

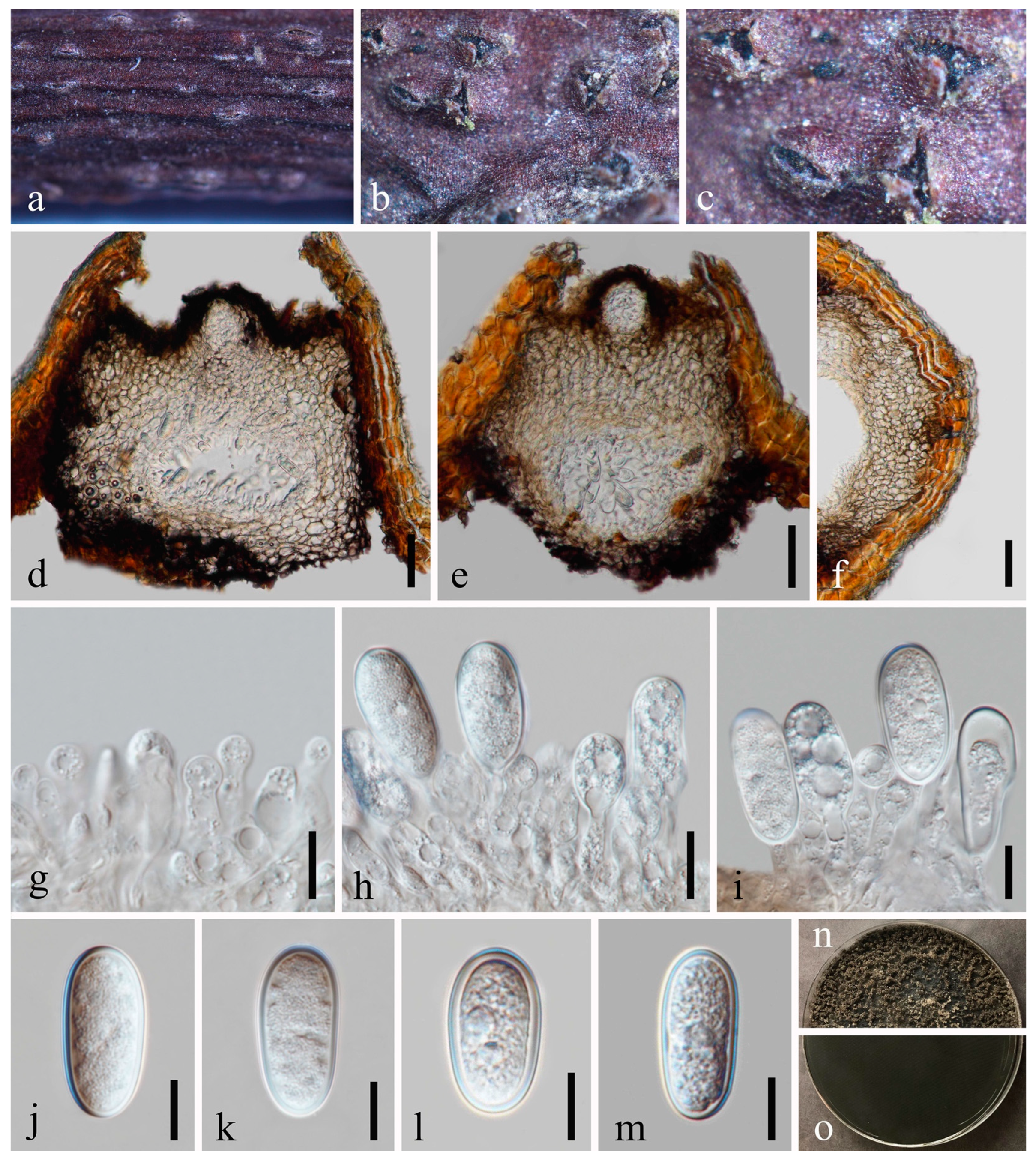
Figure 6.
Dothiorella ovata (MFLU 23-0009, holotype). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Ostiolar region with periphyses. (g–j) Conidiogenous cells and developing conidia. (k–o) Conidia. (p) Germinated conidium. (q,r) Colonies on PDA, above (q) and below (r). Scale bars: (b,c) = 50 μm, (d,e) = 20 μm, (f) = 10 μm, (g,h) = 5 μm, (i–p) = 10 μm.
Figure 6.
Dothiorella ovata (MFLU 23-0009, holotype). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Ostiolar region with periphyses. (g–j) Conidiogenous cells and developing conidia. (k–o) Conidia. (p) Germinated conidium. (q,r) Colonies on PDA, above (q) and below (r). Scale bars: (b,c) = 50 μm, (d,e) = 20 μm, (f) = 10 μm, (g,h) = 5 μm, (i–p) = 10 μm.

Figure 7.
Dothiorella rosacearum (MFLU 23-0014, holotype). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f–i) Conidiogenous cells and developing conidia. (j–m) Conidia. (n) Germinated conidium. (o,p) Colonies on PDA, above (o) and below (p). Scale bars: (d,e) = 50 μm, (f–n) = 10 μm.
Figure 7.
Dothiorella rosacearum (MFLU 23-0014, holotype). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f–i) Conidiogenous cells and developing conidia. (j–m) Conidia. (n) Germinated conidium. (o,p) Colonies on PDA, above (o) and below (p). Scale bars: (d,e) = 50 μm, (f–n) = 10 μm.

Figure 8.
Dothiorella septata (MFLU 23-0007, holotype). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Ostiolar region with periphyses. (g–k) Conidiogenous cells and developing conidia. (l–o) Conidia. (p) Germinated conidium. (q–s) Colonies on PDA, above (q,r) and below (s). Scale bars: (b) = 50 μm, (c,d) = 20 μm, (e–p) = 10 μm.
Figure 8.
Dothiorella septata (MFLU 23-0007, holotype). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Ostiolar region with periphyses. (g–k) Conidiogenous cells and developing conidia. (l–o) Conidia. (p) Germinated conidium. (q–s) Colonies on PDA, above (q,r) and below (s). Scale bars: (b) = 50 μm, (c,d) = 20 μm, (e–p) = 10 μm.

Figure 9.
Lasiodiplodia crassispora (MFLU 23-0011). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Paraphyses. (g–i) Conidiogenous cells and developing conidia. (j–m) Conidia. (n) Germinated conidium. (o,p) Colonies on PDA, above (o) and below (p). Scale bars: (b) = 500 μm, (c) = 200 μm, (d,e) = 50 μm, (f–i) = 10 μm, (j–n) = 20 μm.
Figure 9.
Lasiodiplodia crassispora (MFLU 23-0011). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Paraphyses. (g–i) Conidiogenous cells and developing conidia. (j–m) Conidia. (n) Germinated conidium. (o,p) Colonies on PDA, above (o) and below (p). Scale bars: (b) = 500 μm, (c) = 200 μm, (d,e) = 50 μm, (f–i) = 10 μm, (j–n) = 20 μm.

Figure 10.
Lasiodiplodia delonicis (MFLU 23-0005, holotype). (a) Specimen. (b,c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Paraphyses. (g–i) Conidiogenous cells and developing conidia. (j–l) Conidia. (m) Germinated conidium. (n,o) Colonies on PDA, above (n) and below (o). Scale bars: (b) = 50 μm, (c) = 20 μm, (d,e) = 50 μm, (f–m) = 20 μm.
Figure 10.
Lasiodiplodia delonicis (MFLU 23-0005, holotype). (a) Specimen. (b,c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Paraphyses. (g–i) Conidiogenous cells and developing conidia. (j–l) Conidia. (m) Germinated conidium. (n,o) Colonies on PDA, above (n) and below (o). Scale bars: (b) = 50 μm, (c) = 20 μm, (d,e) = 50 μm, (f–m) = 20 μm.

Figure 11.
Lasiodiplodia mahajangana (MFLU 23-0006). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Paraphyses. (g–j) Conidiogenous cells and developing conidia. (k–n) Conidia. (o) Germinated conidium. (p–r) Colonies on PDA, above (p,r) and below (q). Scale bars: (b) = 100 μm, (c) = 20 μm, (d) = 50 μm, (e) = 20 μm, (f–o) = 10 μm.
Figure 11.
Lasiodiplodia mahajangana (MFLU 23-0006). (a–c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Paraphyses. (g–j) Conidiogenous cells and developing conidia. (k–n) Conidia. (o) Germinated conidium. (p–r) Colonies on PDA, above (p,r) and below (q). Scale bars: (b) = 100 μm, (c) = 20 μm, (d) = 50 μm, (e) = 20 μm, (f–o) = 10 μm.

Figure 12.
Macrophomina euphorbiicola (MFLU 23-0004). (a) Specimen. (b,c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Ostiole. (g,h) Conidiogenous cells and developing conidia. (i–l) Conidia bearing apical appendages (arrows). (m) Germinated conidium. (n,o) Colonies on PDA, above (n) and below (o). Scale bars: (b) = 50 μm, (c–e) = 20 μm, (f–m) = 10 μm.
Figure 12.
Macrophomina euphorbiicola (MFLU 23-0004). (a) Specimen. (b,c) Conidiomata on host substrate. (d) Vertical section of conidiomata. (e) Section of peridium. (f) Ostiole. (g,h) Conidiogenous cells and developing conidia. (i–l) Conidia bearing apical appendages (arrows). (m) Germinated conidium. (n,o) Colonies on PDA, above (n) and below (o). Scale bars: (b) = 50 μm, (c–e) = 20 μm, (f–m) = 10 μm.

Figure 13.
Sphaeropsis eucalypticola (MFLU 23-0008). (a–c) Ascomata on host substrate. (d) Vertical section of ascoma. (e) Structure of peridium. (f) Ostiole. (g) Pseudoparaphyses. (h–j) Asci. (k–o) Ascospores. (p) Germinated ascospore. (q,r) Colonies on PDA, above (q) and below (r). Scale bars: (b) = 50 μm, (c) = 20 μm, (d–f) = 50 μm, (g) = 10 μm, (h–j) = 20 μm, (k–p) = 10 μm.
Figure 13.
Sphaeropsis eucalypticola (MFLU 23-0008). (a–c) Ascomata on host substrate. (d) Vertical section of ascoma. (e) Structure of peridium. (f) Ostiole. (g) Pseudoparaphyses. (h–j) Asci. (k–o) Ascospores. (p) Germinated ascospore. (q,r) Colonies on PDA, above (q) and below (r). Scale bars: (b) = 50 μm, (c) = 20 μm, (d–f) = 50 μm, (g) = 10 μm, (h–j) = 20 μm, (k–p) = 10 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers and PCR protocols used in this study.
| Locus | Primers | Optimized PCR Protocols | Reference |
|---|---|---|---|
| ITS | ITS5 | 94 °C 3 min; 35 cycles of 94 °C 30 s, 55 °C 50 s, 72 °C 1 min; 72 °C 10 min; 4 °C on hold | [25] |
| ITS4 | |||
| LSU | LR0R | 94 °C 3 min; 35 cycles of 94 °C 30 s, 55 °C 50 s, 72 °C 1 min; 72 °C 10 min; 4 °C on hold | [26] |
| LR5 | |||
| tef1-α | EF1-728F | 94 °C 3 min; 35 cycles of 94 °C 30 s, 55 °C 50 s, 72 °C 1 min; 72 °C 10 min; 4 °C on hold | [27] |
| EF1-986R | |||
| tub2 | Bt2a | 94 °C 3 min; 35 cycles of 94 °C 30 s, 55 °C 50 s, 72 °C 1 min; 72 °C 10 min; 4 °C on hold | [28] |
| Bt2b |
Table 2.
Taxa Names, Strain or Specimen numbers, and corresponding GenBank accession numbers of the taxa used for the phylogenetic studies. The newly generated sequences are indicated in red, and ex-type strains are indicated in bold.
Table 2.
Taxa Names, Strain or Specimen numbers, and corresponding GenBank accession numbers of the taxa used for the phylogenetic studies. The newly generated sequences are indicated in red, and ex-type strains are indicated in bold.
| Taxa Names | Strain/Specimen Numbers | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| ITS | LSU | tef1-α | tub2 | ||
| Alanphillipsia aloeicola | CBS 138896 | KP004444 | KP004472 | MT592027 | – |
| Alanphillipsia aloetica | CBS 136409 | KF777139 | KF777195 | MT592028 | – |
| Barriopsis archontophoenicis | MFLUCC 14-1164 | KX235306 | KX235307 | KX235305 | – |
| Barriopsis stevensiana | CBS 174.26 | EU673330 | DQ377857 | EU673296 | – |
| Botryobambusa fusicoccum | MFLUCC 11-0143 | JX646792 | JX646809 | JX646857 | – |
| Botryobambusa guizhouensis | CGMCC 3.20348 | MZ781425 | MZ781492 | MZ852498 | – |
| Botryosphaeria dothidea | CBS 115476 | AY236949 | AY928047 | AY236898 | AY236927 |
| Botryosphaeria dolichospermatii | CGMCC 3.19096 | MH491970 | MH562323 | MH491974 | MH562327 |
| Botryosphaeria fabicerciana | CBS 127193 | HQ332197 | MF410028 | HQ332213 | KF779068 |
| Botryosphaeria fabicerciana | CBS 127194 | HQ332198 | MF410029 | HQ332214 | KF779069 |
| Botryosphaeria fujianensis | CGMCC 3.19099 | MH491973 | MH562326 | MH491977 | MH562330 |
| Botryosphaeria fujianensis | BJFUCC 180226-3 | MW251380 | MW251381 | MW251388 | MW251379 |
| Botryosphaeria fujianensis | MFLUCC 23-0041 | OR052056 | OR052040 | OR030453 | OR030471 |
| Botryosphaeria qingyuanensis | CGMCC 3.18742 | KX278000 | MF410042 | KX278105 | KX278209 |
| Botryosphaeria tenuispora | MUCC 237 | LC585278 | – | LC585150 | LC585174 |
| Cophinforma eucalypti | MFLUCC 11-0425 | JX646800 | JX646817 | JX646865 | JX646848 |
| Cophinforma eucalypti | MFLUCC 11-0655 | JX646801 | JX646818 | JX646866 | JX646849 |
| Cophinforma mamane | CBS 117444 | KF531822 | DQ377855 | KF531801 | KF531802 |
| Diplodia mutila | CBS 112553 | AY259093 | AY928049 | AY573219 | DQ458850 |
| Diplodia mutila | CBS 112875 | AY343484 | – | AY343370 | MT592509 |
| Diplodia mutila | GZCC 23-0578 | OR052057 | OR020607 | OR030454 | OR030472 |
| Diplodia neojuniperi | CBS 138652 | KM006431 | – | KM006462 | MT592516 |
| Diplodia sapinea | CBS 393.84 | DQ458895 | DQ377893 | DQ458880 | DQ458863 |
| Diplodia scrobiculata | CBS 118110 | AY253292 | KF766326 | AY624253 | AY624258 |
| Diplodia seriata | CBS 112555 | AY259094 | AY928050 | AY573220 | DQ458856 |
| Diplodia seriata | CBS 112661 | MT587378 | – | MT592084 | MT592541 |
| Diplodia seriata | GZCC 23-0579 | OR052058 | OR052041 | OR030455 | OR030473 |
| Diplodia subglobosa | CBS 124133 | GQ923856 | – | GQ923824 | MT592576 |
| Dothiorella acacicola | CBS 141295 | KX228269 | KX228320 | KX228376 | – |
| Dothiorella acericola | KUMCC 18-0137 | MK359449 | – | MK361182 | – |
| Dothiorella albiziae | MFLUCC 22-0057 | ON751762 | ON751764 | ON799588 | ON799590 |
| Dothiorella alpina | CGMCC 3.18001 | KX499645 | – | KX499651 | – |
| Dothiorella baihuashanensis | CFCC 58549 | – | – | OQ692933 | OQ692927 |
| Dothiorella baihuashanensis | CFCC 58788 | – | – | OQ692934 | OQ692928 |
| Dothiorella brevicollis | CBS 130411 | JQ239403 | JQ239416 | JQ239390 | JQ239371 |
| Dothiorella camelliae | CGMCC 3.24158 | OQ190531 | – | OQ241464 | OQ275064 |
| Dothiorella capri-amissi | CBS 121763 | EU101323 | KX464301 | EU101368 | KX464850 |
| Dothiorella casuarinae | CBS 120688 | DQ846773 | MH874647 | DQ875331 | DQ875340 |
| Dothiorella citricola | CBS 124728 | EU673322 | – | EU673289 | KX464852 |
| Dothiorella citrimurcotticola | CGMCC 3.20394 | MW880661 | – | MW884164 | MW884193 |
| Dothiorella citrimurcotticola | CGMCC 3.20395 | MW880662 | – | MW884165 | MW884194 |
| Dothiorella diospyricola | CBS 145972 | MT587398 | – | MT592110 | MT592581 |
| Dothiorella dulcispinae | CBS 130413 | JQ239400 | JQ239413 | JQ239387 | JQ239373 |
| Dothiorella dulcispinae | CMW 36462 | JQ239402 | JQ239415 | JQ239389 | JQ239375 |
| Dothiorella eriobotryae | CBS 140852 | KT240287 | – | KT240262 | MT592582 |
| Dothiorella heterophyllae | CMW46458 | MN103794 | – | MH548348 | MH548324 |
| Dothiorella iranica | CBS 124722 | KC898231 | – | KC898214 | KX464856 |
| Dothiorella koae | CMW 48017 | MH447652 | – | MH548338 | MH548327 |
| Dothiorella lampangensis | MFLUCC 18-0232 | MK347758 | – | MK340869 | MK412874 |
| Dothiorella longicollis | CBS 122068 | EU144054 | MH874718 | EU144069 | KF766130 |
| Dothiorella magnoliae | CFCC 51563 | KY111247 | – | KY213686 | – |
| Dothiorella mangifericola | IRAN 1584C | MT587407 | – | MT592119 | – |
| Dothiorella moneti | MUCC 505 | EF591920 | EF591937 | EF591971 | EF591954 |
| Dothiorella obovata | MFLUCC 22-0058 | ON751763 | ON751765 | ON799589 | ON799591 |
| Dothiorella ovata | MFLUCC 23-0035 | OR052059 | OR020691 | OR030456 | OR030474 |
| Dothiorella ovata | MFLUCC 23-0036 | OR052060 | OR052042 | OR030457 | OR030475 |
| Dothiorella plurivora | CBS 124724 | KC898225 | – | KC898208 | KX464874 |
| Dothiorella pretoriensis | CBS 130404 | JQ239405 | JQ239418 | JQ239392 | JQ239376 |
| Dothiorella prunicola | CBS 124723 | EU673313 | EU673232 | EU673280 | EU673100 |
| Dothiorella rosacearum | MFLUCC 23-0038 | OR052061 | OR052043 | OR030458 | OR030476 |
| Dothiorella rosacearum | MFLUCC 23-0037 | OR052062 | OR052044 | OR030459 | OR030477 |
| Dothiorella santali | WAC 13155 | EF591924 | EF591941 | EF591975 | EF591958 |
| Dothiorella sarmentorum | CBS 115038 | AY573206 | DQ377860 | AY573223 | EU673101 |
| Dothiorella sarmentorum | IMI 63581b | AY573212 | AY928052 | AY573235 | – |
| Dothiorella septata | MFLUCC 23-0039 | OR020942 | OR020695 | OR030462 | OR030480 |
| Dothiorella septata | GZCC 23-0583 | OR019776 | OR052047 | OR030463 | OR030481 |
| Dothiorella septata | GZCC 23-0584 | OR019803 | OR052048 | OR030464 | OR030482 |
| Dothiorella striata | CBS 124731 | EU673321 | – | EU673288 | EU673143 |
| Dothiorella striata | CBS 124730 | EU673320 | EU673240 | EU673287 | EU673142 |
| Dothiorella tectonae | MFLUCC 18-0382 | KM396899 | – | KM409637 | KM510357 |
| Dothiorella thailandica | MFLUCC 11-0438 | JX646796 | JX646813 | JX646861 | JX646844 |
| Dothiorella thripsita | CBS 125445 | FJ824738 | – | KJ573639 | KJ577550 |
| Dothiorella ulmacea | CBS 138855 | KR611881 | KR611899 | KR611910 | KR611909 |
| Dothiorella ulmacea | CBS 140005 | KR611882 | – | KR857697 | MT592607 |
| Dothiorella uruguayensis | CBS 124908 | EU080923 | MH874932 | EU863180 | KX464886 |
| Dothiorella vinea-gemmae | DAR 81012 | KJ573644 | – | KJ573641 | KJ577552 |
| Dothiorella viticola | CBS 117009 | AY905554 | MH874565 | AY905559 | EU673104 |
| Dothiorella yunnana | CGMCC 3.18000 | KX499644 | – | KX499650 | – |
| Dothiorella zanthoxyli | CGMCC 3.24159 | OQ190536 | – | OQ241468 | OQ275069 |
| Endomelanconiopsis endophytica | CBS 120397 | EU683656 | EU683629 | EU683637 | KF766131 |
| Endomelanconiopsis freycinetiae | MFLUCC 17-0547 | MG646955 | MG646948 | MG646983 | MG646924 |
| Eutiarosporella africana | CMW 38423 | KC769956 | KC769990 | KC769852 | – |
| Eutiarosporella dactylidis | MFLUCC 13-0276 | KM978944 | KM978949 | KP031694 | – |
| Eutiarosporella urbis-rosarum | CMW 36477 | JQ239407 | JQ239420 | JQ239394 | JQ239381 |
| Lasiodiplodia crassispora | CBS 118741 | DQ103550 | DQ377901 | DQ103557 | KU887506 |
| Lasiodiplodia crassispora | CMW 13488 | DQ103552 | – | DQ103559 | KU887507 |
| Lasiodiplodia crassispora | MFLUCC 23-0060 | OR052065 | OR020699 | OR030465 | OR030483 |
| Lasiodiplodia delonicis | MFLUCC 23-0058 | OR052066 | OR052049 | OR030466 | OR030484 |
| Lasiodiplodia mahajangana | CBS 124925 | FJ900595 | – | FJ900641 | FJ900630 |
| Lasiodiplodia mahajangana | CBS 124926 | FJ900596 | – | FJ900642 | FJ900631 |
| Lasiodiplodia mahajangana | MFLUCC 23-0059 | OR052067 | OR052050 | OR030467 | OR030485 |
| Lasiodiplodia rubropurpurea | WAC 12535 | DQ103553 | DQ377903 | DQ103571 | EU673136 |
| Lasiodiplodia rubropurpurea | WAC 12536 | DQ103554 | – | DQ103572 | KU887530 |
| Lasiodiplodia thailandica | CBS 138760 | KJ193637 | – | KJ193681 | – |
| Lasiodiplodia thailandica | CBS 138653 | KM006433 | – | KM006464 | – |
| Lasiodiplodia theobromae | CBS 164.96 | AY640255 | EU673253 | AY640258 | KU887532 |
| Lasiodiplodia theobromae | CBS 111530 | EF622074 | – | EF622054 | KU887531 |
| Lasiodiplodia venezuelensis | WAC 12539 | DQ103547 | DQ377904 | DQ103568 | KU887533 |
| Lasiodiplodia venezuelensis | WAC 12540 | DQ103548 | – | DQ103569 | KU887534 |
| Lasiodiplodia viticola | CBS 128313 | HQ288227 | KX098286 | HQ288269 | HQ288306 |
| Lasiodiplodia viticola | CBS 128314 | HQ288228 | – | HQ288270 | HQ288307 |
| Macrophomina euphorbiicola | CMM 4134 | KU058936 | – | KU058906 | MF457658 |
| Macrophomina euphorbiicola | CMM 4045 | KU058928 | – | KU058898 | MF457657 |
| Macrophomina euphorbiicola | MFLUCC 23-0057 | OR052068 | OR052051 | OR030468 | OR030486 |
| Macrophomina phaseolina | CBS 205.47 | KF951622 | – | KF951997 | MW592323 |
| Macrophomina pseudophaseolina | CBS 137165 | KF951791 | – | KF952153 | KF952233 |
| Macrophomina tecta | BRIP 70781 | MW591684 | – | MW592271 | MW592300 |
| Marasasiomyces karoo | CBS 118718 | KF531828 | DQ377939 | KF531807 | KF531808 |
| Mucoharknessia cortaderiae | CPC 19974 | KM108374 | KM108401 | – | – |
| Mucoharknessia anthoxanthii | MFLUCC 15-0904 | KU246377 | KU246379 | – | – |
| Neodeightonia subglobosa | CBS 448.91 | EU673337 | DQ377866 | EU673306 | EU673137 |
| Neodeightonia phoenicum | CBS 122528 | EU673340 | EU673261 | EU673309 | EU673116 |
| Neodeightonia phoenicum | CBS 123168 | EU673339 | EU673260 | EU673308 | EU673115 |
| Neoscytalidium dimidiatum | CBS 145.78 | KF531816 | DQ377922 | KF531795 | KF531796 |
| Neoscytalidium dimidiatum | CBS 251.49 | KF531819 | DQ377923 | KF531797 | KF531799 |
| Oblongocollomyces variabilis | CBS 121774 | EU101312 | KX464536 | EU101357 | – |
| Oblongocollomyces variabilis | CBS 121775 | EU101314 | MT587319 | EU101359 | – |
| Phaeobotryon mamane | CBS 122980 | EU673332 | EU673248 | EU673298 | – |
| Phaeobotryon cupressi | CBS 124700 | FJ919672 | KX464538 | FJ919661 | – |
| Sakireeta madreeya | CBS 532.76 | KC769960 | DQ377940 | KM108427 | KX465084 |
| Sardiniella celtidis | MFLUCC 17-981 | MF443249 | – | MF443248 | – |
| Sardiniella urbana | CBS 141580 | KX379674 | KX379676 | KX379675 | – |
| Sardiniella urbana | BL180 | KX379677 | KX379679 | KX379678 | – |
| Sphaeropsis citrigena | ICMP 16812 | EU673328 | EU673246 | EU673294 | EU673140 |
| Sphaeropsis eucalypticola | MFLUCC 11-0579 | JX646802 | JX646819 | JX646867 | JX646850 |
| Sphaeropsis eucalypticola | MFLUCC 11-0654 | JX646803 | JX646820 | JX646868 | JX646851 |
| Sphaeropsis eucalypticola | MFLUCC 23-0040 | OR052069 | OR052052 | OR030469 | OR030488 |
| Sphaeropsis eucalypticola | GZCC 23-0589 | OR052070 | OR052053 | OR030470 | OR030487 |
| Sphaeropsis porosa | CBS 110496 | AY343379 | DQ377894 | AY343340 | EU673130 |
| Sphaeropsis visci | CBS 100163 | EU673324 | EU754215 | EU673292 | EU673127 |
| Sphaeropsis guizhouensis | CGMCC 3.20352 | MZ781433 | MZ781500 | MZ852506 | – |
| Tiarosporella palludosa | CPC 22701 | KM108378 | KM108404 | – | KM108471 |
| Tiarosporella palludosa | CPC 22702 | KM108379 | KM108405 | – | KM108472 |
| Neofusicoccum mangiferae | CBS 118531 | AY615185 | DQ377921 | DQ093221 | AY615172 |
| Neofusicoccum parvum | CMW 9081 | AY236943 | AY928045 | AY236888 | AY236917 |
| Neofusicoccum parvum | CBS 110301 | AY259098 | AY928046 | AY573221 | EU673095 |
| Pseudofusicoccum adansoniae | CBS 122055 | EF585523 | – | EF585571 | MT592771 |
| Pseudofusicoccum adansoniae | CBS 122056 | EF585524 | – | MT592279 | MT592772 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, N.; Dissanayake, A.J.; Du, H.-Z.; Liu, J.-K. Morpho-Phylogenetic Evidence Reveals Novel Species and New Records of Botryosphaeriaceae in China and Thailand. J. Fungi 2023, 9, 1051. https://doi.org/10.3390/jof9111051
AMA Style
Wu N, Dissanayake AJ, Du H-Z, Liu J-K. Morpho-Phylogenetic Evidence Reveals Novel Species and New Records of Botryosphaeriaceae in China and Thailand. Journal of Fungi. 2023; 9(11):1051. https://doi.org/10.3390/jof9111051
Chicago/Turabian StyleWu, Na, Asha J. Dissanayake, Hong-Zhi Du, and Jian-Kui Liu. 2023. "Morpho-Phylogenetic Evidence Reveals Novel Species and New Records of Botryosphaeriaceae in China and Thailand" Journal of Fungi 9, no. 11: 1051. https://doi.org/10.3390/jof9111051
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.