
Taxonomic and Phylogenetic Updates on Apiospora: Introducing Four New Species from Wurfbainia villosa and Grasses in China

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Observation, and Isolation
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
2.4. Pairwise Homoplasy Index (PHI)
3. Results
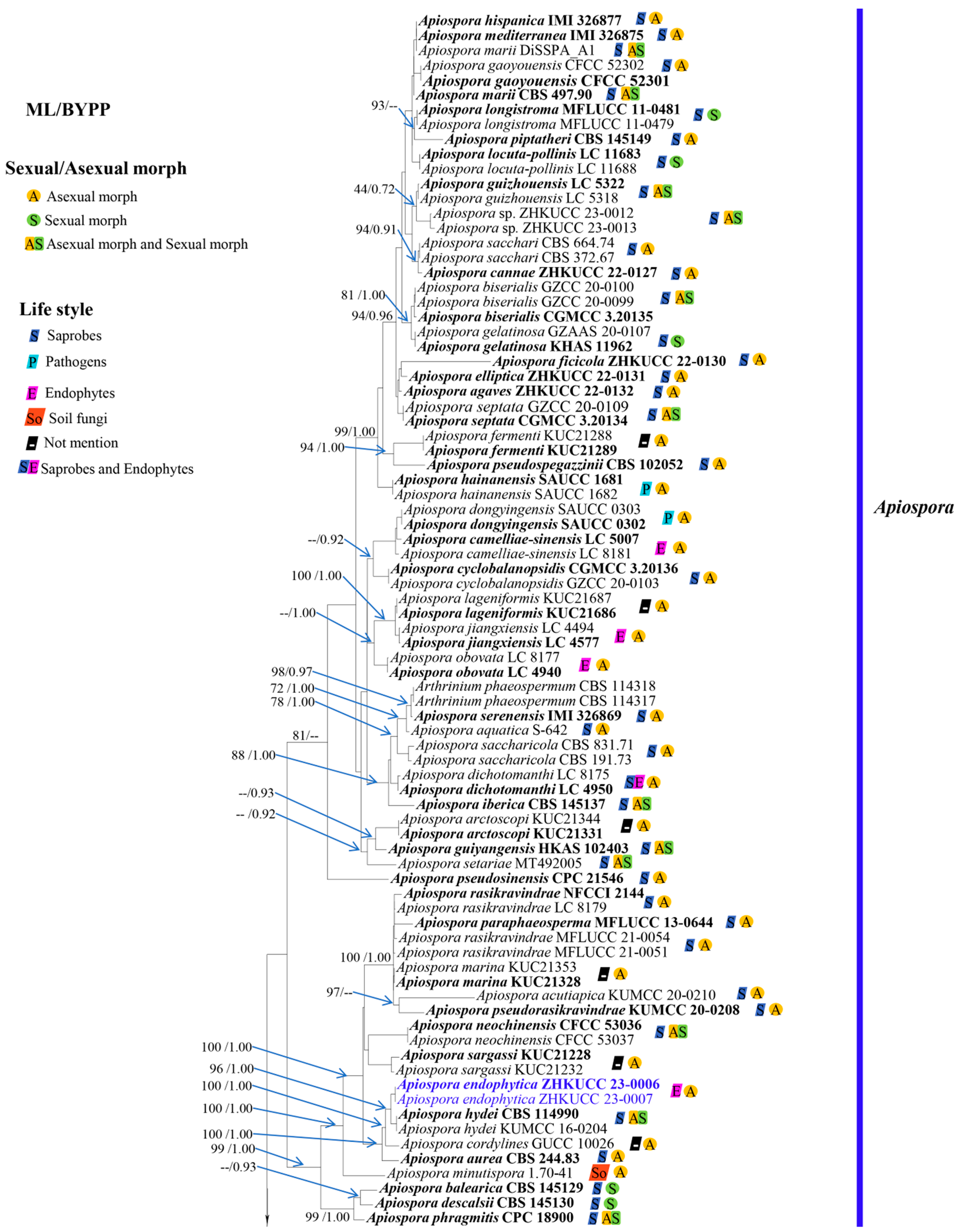
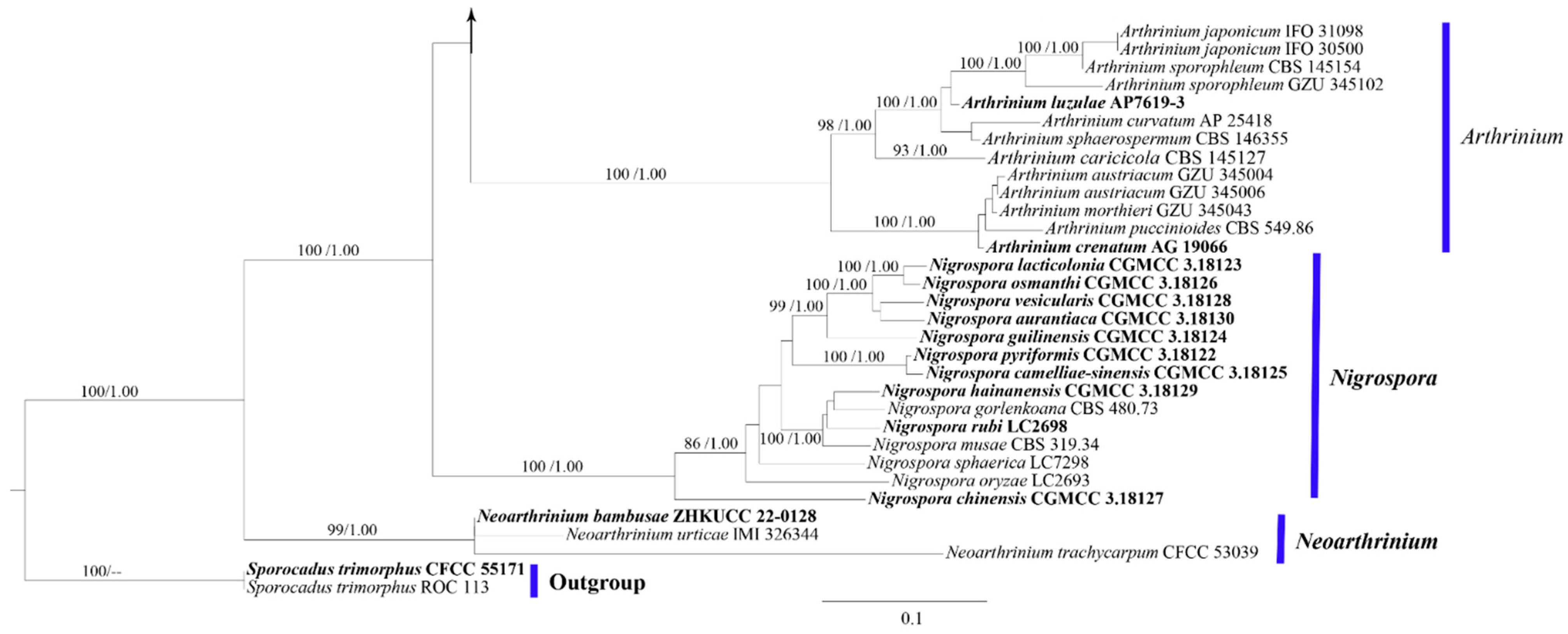
3.1. Phylogeny
3.2. A Pairwise Homoplasy Index
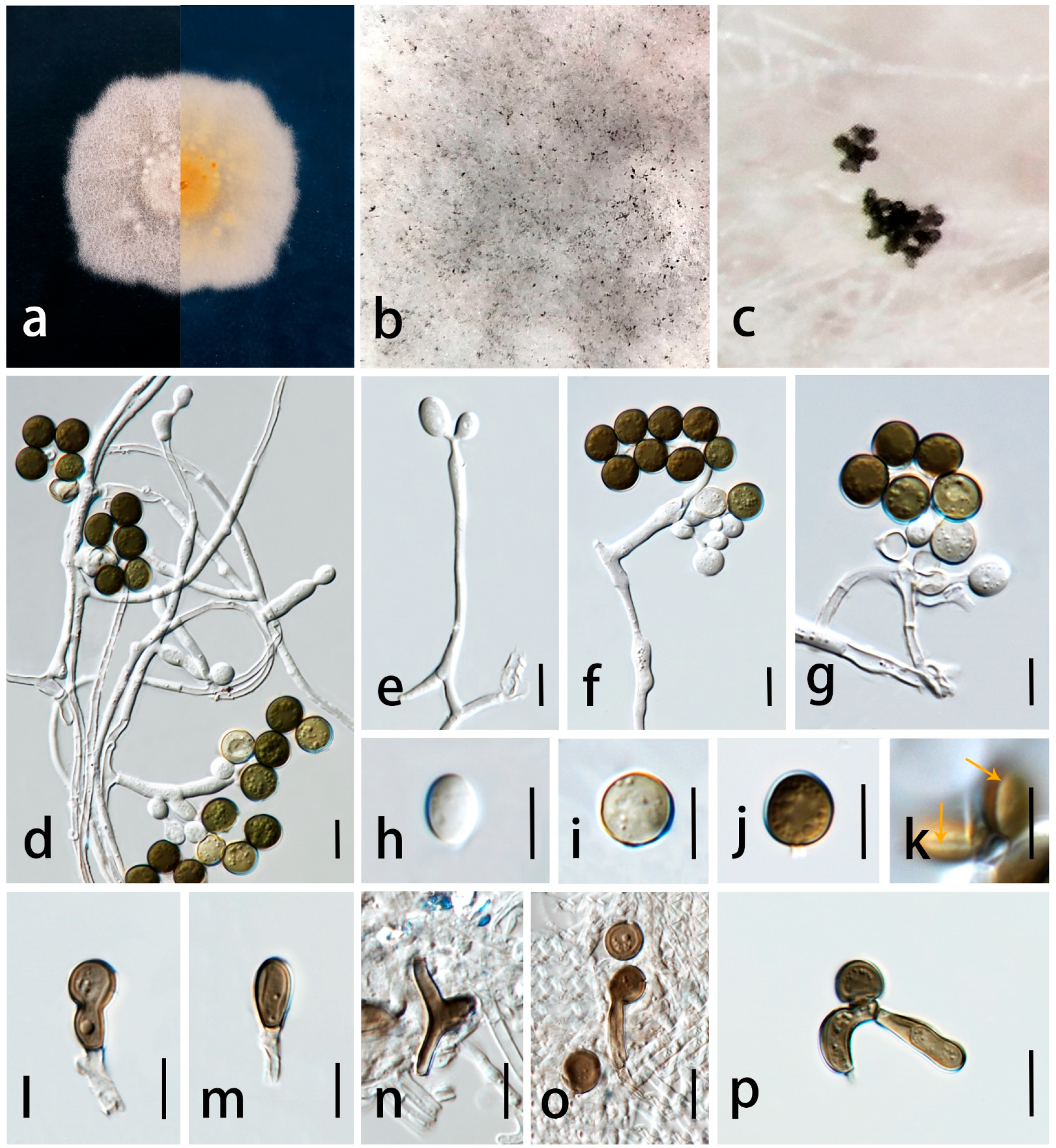
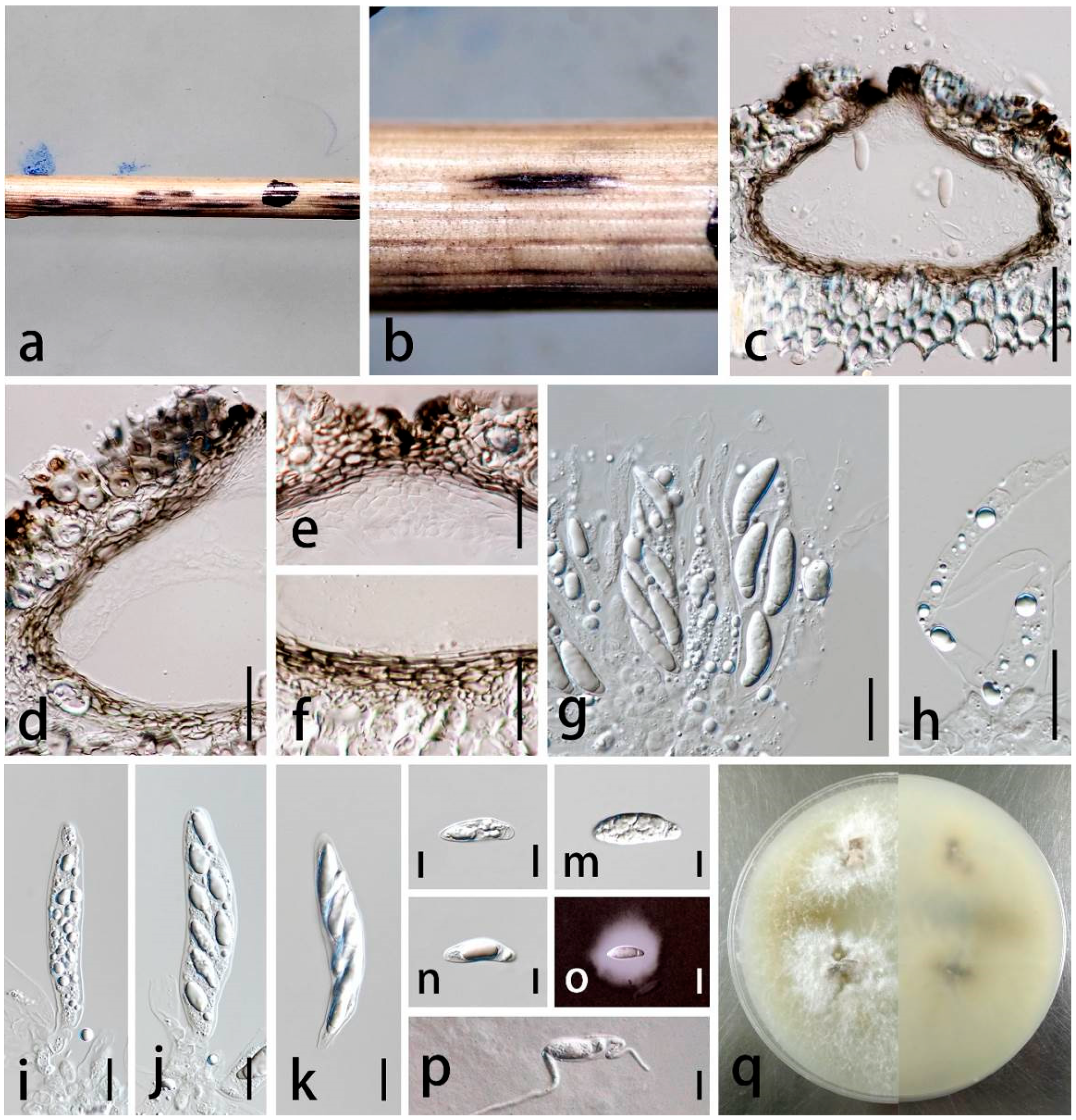
3.3. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phookamsak, R.; Liu, J.K.; McKenzie, E.H.C.; Manamgoda, D.S.; Ariyawansa, H.; Thambugala, K.M.; Dai, D.Q.; Camporesi, E.; Chukeatirote, E.; Wijayawardene, N.N.; et al. Revision of Phaeosphaeriaceae. Fungal Divers. 2014, 68, 159–238. [Google Scholar]
- Hongsanan, S.; Li, Y.M.; Liu, J.K.; Hofmann, T.; Piepenbring, M.; Bhat, D.J.; Boonmee, S.; Doilom, M.; Singtripop, C.; Tian, Q.; et al. Revision of genera in Asterinales. Fungal Divers. 2014, 68, 1–68. [Google Scholar]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [PubMed]
- Pem, D.; Jeewon, R.; Chethana, K.W.T.; Hongsanan, S.; Doilom, M.; Suwannarach, N.; Hyde, K.D. Species concepts of Dothideomycetes: Classification, phylogenetic inconsistencies and taxonomic standardization. Fungal Divers. 2021, 109, 283–319. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa–2021. Mycosphere 2022, 13, 53–453. [Google Scholar]
- Saccardo, P. Conspectus generum pyrenomycetum italicorum additis speciebus fungorum Venetorum novisvel criticis, systemate carpologico dispositorum. Atti della Societa Veneziana-Trentina-Istrianadi. Scienze Naturali 1875, 4, 77–100. [Google Scholar]
- Clements, F.E.; Shear, C.L. The genera of Fungi; H.W. Wilson Company publishing: New York, USA, 1931; pp. 1–496. [Google Scholar]
- Crous, P.W.; Groenewald, J.Z. A phylogenetic reevaluation of Arthrinium. IMA Fungus 2013, 4, 133–154. [Google Scholar]
- Pintos, Á.; Alvarado, P.; Planas, J.; Jarling, R. Six new species of Arthrinium from Europe and notes about A. caricicola and other species found in Carex spp. hosts. MycoKeys 2019, 49, 15–48. [Google Scholar]
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Zhang, N. The Amsterdam declaration on fungal nomenclature. IMA Fungus 2011, 2, 105–112. [Google Scholar]
- Réblová, M.; Miller, A.N.; Rossman, A.Y.; Seifert, K.A.; Crous, P.W.; Hawksworth, D.L.; Abdel-Wahab, M.A.; Cannon, P.F.; Daranagama, D.A.; De Beer, Z.W.; et al. Recommendations for competing sexual-asexually typified generic names in Sordariomycetes (except Diaporthales, Hypocreales, and Magnaporthales). IMA Fungus 2016, 7, 131–153. [Google Scholar]
- Wang, M.; Tan, X.M.; Liu, F.; Cai, L. Eight new Arthrinium species from China. MycoKeys 2018, 34, 1–24. [Google Scholar]
- Jiang, N.; Liang, Y.M.; Tian, C.M. A novel bambusicolous fungus from China, Arthrinium chinense (Xylariales). Sydowia 2020, 72, 77–83. [Google Scholar]
- Feng, Y.; Liu, J.K.; Lin, C.G.; Chen, Y.Y.; Xiang, M.M.; Liu, Z.Y. Additions to the genus Arthrinium (Apiosporaceae) from bamboos in China. Front. Microbiol. 2021, 7, 661281. [Google Scholar]
- Pintos, Á.; Alvarado, P. Phylogenetic delimitation of Apiospora and Arthrinium. Fungal Syst. Evol. 2021, 7, 197–221. [Google Scholar] [CrossRef]
- Tian, X.G.; Karunarathna, S.C.; Mapook, A.; Promputtha, I.; Xu, J.; Bao, D.; Tibpromma, S. One new species and two new host records of Apiospora from bamboo and maize in northern Thailand with thirteen new combinations. Life 2021, 11, 1071. [Google Scholar] [CrossRef] [PubMed]
- Index Fungorum. Available online: http://www.indexfungorum.org (accessed on 17 October 2023).
- Kwon, S.L.; Cho, M.; Lee, Y.M.; Kim, C.; Lee, S.M.; Ahn, B.J.; Lee, H.; Kim, J.J. Two unrecorded Apiospora species isolated from marine substrates in Korea with eight new combinations (A. piptatheri and A. rasikravindrae). Mycobiology 2022, 50, 46–54. [Google Scholar] [CrossRef]
- Ji, Z.L.; Zhang, S.W.; Zhu, F.; Wan, B.X.; Liang, R.Z. First report of Arthrinium arundinis causing leaf edge spot of peach in China. Plant Dis. 2020, 104, 3077. [Google Scholar] [CrossRef]
- Chen, K.; Wu, X.Q.; Huang, M.X.; Han, Y.Y. First report of brown culm streak of Phyllostachys praecox caused by Arthrinium arundinis in Nanjing, China. Plant Dis. 2014, 98, 1274. [Google Scholar] [CrossRef]
- Mavragani, D.C.; Abdellatif, L.; McConkey, B.; Hamel, C.; Vujanovic, V. First report of damping-off of durum wheat caused by Arthrinium sacchari in the semi-arid Saskatchewan fields. Plant Dis. 2007, 91, 469. [Google Scholar] [CrossRef]
- Li, B.J.; Liu, P.Q.; Jiang, Y.; Weng, Q.Y.; Chen, Q.H. First report of culm rot caused by Arthrinium phaeospermum on Phyllostachys viridis in China. Plant Dis. 2016, 100, 1013. [Google Scholar] [CrossRef]
- Dyląg, M.; Hryncewicz-Gwóźdź, A.; Jagielski, T. Onychomycosis due to Arthrinium arundinis: A case report. Acta Derm. Venereol. 2017, 97, 860–861. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Chomnunti, P.; Doilom, M.; Daranagama, D.A.; Kang, J. Multigene phylogeny reveals endophytic Xylariales novelties from Dendrobium species from Southwestern China and Northern Thailand. J. Fungi 2022, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Wang, H.; Umeokoli, B.O.; Eze, P.; Heering, C.; Janiak, C.; Müller, W.E.; Orfali, R.S.; Hartmann, R.; Dai, H.; Lin, W.; et al. Secondary metabolites of the lichen-associated fungus Apiospora montagnei. Tetrahedron Lett. 2017, 58, 1702–1705. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jiang, N.; Tian, C.M. The holomorph of Arthrinium setariae sp. nov. (Apiosporaceae, Xylariales) from China. Phytotaxa 2021, 483, 149–159. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Chen, Y.; Ariyawansa, H.A.; Hyde, K.D.; Haelewaters, D.; Perera, R.H.; Samarakoon, M.C.; Wanasinghe, D.N.; Bustamante, D.E.; Liu, J.K.; et al. Integrative approaches for species delimitation in Ascomycota. Fungal Divers. 2021, 109, 155–179. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Chaiwan, N.; Gomdola, D.; Wang, S.; Monkai, J.; Tibpromma, S.; Doilom, M.; Wanasinghe, D.N.; Mortimer, P.E.; Lumyong, S.; Hyde, K.D. https://gmsmicrofungi.org: An online database providing updated information of microfungi in the Greater Mekong Subregion. Mycosphere 2021, 12, 1513–1526. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 18, p. 7. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- GenBank. Available online: http://www.ncbi.nlm.nih.gov/blast/ (accessed on 17 October 2023).
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series, London, UK, 8–12 October 1999; pp. 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA, USA, 21 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4: Tree Figure Drawing Tool. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 17 October 2023).
- Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Z.; Zhang, Y.; Ming, X.B.; Zhang, Q.; Long, H.; Hyde, K.D.; Li, Y.; Wang, Y. Morphological and phylogenetic resolution of Arthrinium from medicinal plants in Yunnan, including A. cordylines and A. pseudomarii spp. nov. Mycotaxon 2021, 136, 183–199. [Google Scholar] [CrossRef]
- Calvo, M.A.; Guarro, J. Arthrinium aureum sp. nov. from Spain. Trans. Br. Mycol. Soc. 1980, 75, 156–157. [Google Scholar] [CrossRef]
- Yang, C.L.; Xu, X.L.; Dong, W.; Wanasinghe, D.N.; Liu, Y.G.; Hyde, K.D. Introducing Arthrinium phyllostachium sp. nov. (Apiosporaceae, Xylariales) on Phyllostachys heteroclada from Sichuan province, China. Phytotaxa 2019, 406, 91–110. [Google Scholar] [CrossRef]
- Monkai, J.; Phookamsak, R.; Tennakoon, D.S.; Bhat, D.J.; Xu, S.; Li, Q.; Xu, J.; Mortimer, P.E.; Kumla, J.; Lumyong, S. Insight into the taxonomic resolution of Apiospora: Introducing novel species and records from bamboo in China and Thailand. Diversity 2022, 14, 918. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.; Sun, Y.R.; Wijesinghe, S.N.; Raza, M.; Bao, D.F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 1–60. [Google Scholar] [CrossRef]
- Larrondo, J.V.; Calvo, M.A. New contributions to the study of the genus Arthrinium. Mycologia 1992, 84, 475–478. [Google Scholar] [CrossRef]
- Zeng, Y.; Ali, M.K.; He, W.; Deng, L.; Yang, X.; Li, X.; Pu, X.; Yang, J. Chemical constituents of functional food Amomum villosum to combat human diseases. Curr. Chin. Sci. 2022, 2, 57–67. [Google Scholar] [CrossRef]
- Bon, M.C.; Goolsby, J.A.; Mercadier, G.; Guermache, F.; Kashefi, J.; Cristofaro, M.; Vacek, A.T.; Kirk, A. Detection of a diverse endophyte assemblage within fungal communities associated with the Arundo Leaf Miner, Lasioptera donacis (Diptera: Cecidomyiidae). Diversity 2023, 15, 571. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–75. [Google Scholar] [CrossRef]
- Shrestha, P.; Ibáñez, A.B.; Bauer, S.; Glassman, S.I.; Szaro, T.M.; Bruns, T.D.; Taylor, J.W. Fungi isolated from Miscanthus and sugarcane: Biomass conversion, fungal enzymes, and hydrolysis of plant cell wall polymers. Biotechnol. Biofuels 2015, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.H.; Jang, S.; Heo, Y.M.; Min, M.; Lee, H.; Lee, Y.M.; Lee, H.; Kim, J.J. Investigation of marine-derived fungal diversity and their exploitable biological activities. Mar. Drugs 2015, 13, 4137–4155. [Google Scholar] [CrossRef]
- Astuti, P.; Pratoko, D.K.; Rollando, R.; Nugroho, G.W.; Wahyuono, S.; Hertiani, T.; Nurrochmad, A. Bioactivities of A Major Compound from Arthrinium rasikravindrae an endophytic fungus of Coleus amboinicus. Lour. Fabad J. Pharm. Sci. 2021, 46, 23–30. [Google Scholar]
- van Eijk, G.W. Bostrycin, a tetrahydroanthraquinone pigment and some other metabolites from the fungus Arthrinium phaeospermum. Experientia 1975, 31, 783–784. [Google Scholar] [CrossRef]
- Li, S.; Tang, Y.; Fang, X.; Qiao, T.; Han, S.; Zhu, T. Whole-genome sequence of Arthrinium phaeospermum, a globally distributed pathogenic fungus. Genomics 2020, 112, 919–929. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Strain Numbers | Substrates | Known Lifestyles | Countries | GenBank Accession Numbers | |||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | tub2 | tef1-α | |||||
| Apiospora acutiapica | KUMCC 20-0210 | Bambusa bambos | Saprobe | China | MT946343 | MT946339 | MT947366 | MT947360 |
| Ap. agari | KUC21333T | Agarum cribrosum | Not mentioned | Republic of Korea | MH498520 | - | MH498478 | MH544663 |
| Ap. agari | KUC21361 | Agarum cribrosum | Not mentioned | Republic of Korea | MH498519 | - | MH498477 | MN868914 |
| Ap. aquatica | S-642 | Submerged wood | Saprobe | China | MK828608 | MK835806 | - | - |
| Ap. arctoscopi | KUC21331T | Egg of Arctoscopus japonicus | Not mentioned | Republic of Korea | MH498529 | - | MH498487 | MN868918 |
| Ap. arctoscopi | KUC21344 | Egg of Arctoscopus japonicus | Not mentioned | Republic of Korea | MH498528 | - | MH498486 | MN868919 |
| Ap. arundinis | CBS 133509 | Aspergillus flavus sclerotium | Saprobe/endophyte | USA | KF144886 | KF144930 | KF144976 | KF145018 |
| Ap. arundinis | CBS 449.92 | Aspergillus flavus sclerotium | Saprobe/endophyte | USA | KF144887 | KF144931 | KF144977 | KF145019 |
| Ap. aurea | CBS 244.83T | - | Saprobe | Japan | AB220251 | KF144935 | KF144981 | KF145023 |
| Ap. balearica | CBS 145129T | Undetermined Poaceae | Saprobe | Spain | MK014869 | MK014836 | MK017975 | MK017946 |
| Ap. bambusicola | MFLUCC 20-0144T | Schizostachyum brachycladum | Saprobe | Thailand | MW173030 | MW173087 | - | MW183262 |
| Ap. biserialis | CGMCC 3.20135T | Bamboo | Saprobe | China | MW481708 | MW478885 | MW522955 | MW522938 |
| Ap. biserialis | GZCC 20-0099 | Bamboo | Saprobe | China | MW481709 | MW478886 | MW522956 | MW522939 |
| Ap. biserialis | GZCC 20-0100 | Bamboo | Saprobe | China | MW481710 | MW478887 | MW522957 | MW522940 |
| Ap. camelliae-sinensis | LC 5007T | Camellia sinensis | Endophyte | China | KY494704 | KY494780 | KY705173 | KY705103 |
| Ap. camelliae-sinensis | LC 8181 | Camellia sinensis | Endophyte | China | KY494761 | KY494837 | KY705229 | KY705157 |
| Ap. chiangraiense | MFLUCC 21-0053T | Dead culms of bamboo | Saprobe | Thailand | MZ542520 | MZ542524 | MZ546409 | - |
| Ap. chromolaenae | MFLUCC 17-1505T | Chromolaena odorata | Saprobe | Thailand | MT214342 | MT214436 | - | MT235802 |
| Ap. cordylines | GUCC 10026 | Cordyline fruticosa | Not mentioned | China | MT040105 | - | MT040147 | MT040126 |
| Ap. cyclobalanopsidis | CGMCC 3.20136T | Cyclobalanopsidis glauca | Saprobe | China | MW481713 | MW478892 | MW522962 | MW522945 |
| Ap. cyclobalanopsidis | GZCC 20-0103 | Cyclobalanopsidis glauca | Saprobe | China | MW481714 | MW478893 | MW522963 | MW522946 |
| Ap. descalsii | CBS 145130T | Ampelodesmos mauritanicus | Saprobe | Spain | MK014870 | MK014837 | MK017976 | MK017947 |
| Ap. dichotomanthi | LC 4950T | Dichotomanthes tristaniicarpa | Saprobe/endophyte | China | KY494697 | KY494773 | KY705167 | KY705096 |
| Ap. dichotomanthi | LC 8175 | Dichotomanthes tristaniicarpa | Saprobe/endophyte | China | KY494755 | KY494831 | KY705223 | KY705151 |
| Ap. dongyingensis | SAUCC 0302T | Leaf of bamboo | Pathogen | China | OP563375 | OP572424 | OP573270 | OP573264 |
| Ap. dongyingensis | SAUCC 0303 | Leaf of bamboo | Pathogen | China | OP563374 | OP572423 | OP573263 | OP573269 |
| Ap. endophytica | ZHKUCC 23-0006T | Wurfbainia villosa | Endophyte | China | OQ587996 | OQ587984 | OQ586062 | OQ586075 |
| Ap. endophytica | ZHKUCC 23-0007 | Wurfbainia villosa | Endophyte | China | OQ587997 | OQ587985 | OQ586063 | OQ586076 |
| Ap. esporlensis | CBS 145136T | Phyllostachys aurea | Saprobe | Spain | MK014878 | MK014845 | MK017983 | MK017954 |
| Ap. euphorbiae | IMI 285638b | Bambusa sp. | Saprobe | Bangladesh | AB220241 | AB220335 | AB220288 | - |
| Ap. fermenti | KUC21289T | Seaweed | Not mentioned | Republic of Korea | MF615226 | - | MF615231 | MH544667 |
| Ap. fermenti | KUC21288 | Seaweed | Not mentioned | Republic of Korea | MF615230 | - | MF615235 | MH544668 |
| Ap. gaoyouensis | CFCC 52301T | Phragmites australis | Saprobe | China | MH197124 | - | MH236789 | MH236793 |
| Ap. gaoyouensis | CFCC 52302 | Phragmites australis | Saprobe | China | MH197125 | - | MH236790 | MH236794 |
| Ap. garethjonesii | KUMCC 16-0202T | Dead culms of bamboo | Saprobe | China | KY356086 | KY356091 | - | - |
| Ap. gelatinosa | KHAS 11962T | Bamboo | Saprobe | China | MW481706 | MW478888 | MW522958 | MW522941 |
| Ap. gelatinosa | GZAAS 20-0107 | Bamboo | Saprobe | China | MW481707 | MW478889 | MW522959 | MW522942 |
| Ap. guangdongensis | ZHKUCC 23-0004T | Wurfbainia villosa | Endophyte | China | OQ587994 | OQ587982 | OQ586060 | OQ586073 |
| Ap. guangdongensis | ZHKUCC 23-0005 | Wurfbainia villosa | Endophyte | China | OQ587995 | OQ587983 | OQ586061 | OQ586074 |
| Ap. guiyangensis | HKAS 102403T | Unidentified grass | Saprobe | China | MW240647 | MW240577 | MW775604 | MW759535 |
| Ap. guizhouensis | LC 5318 | Air in karst cave, bamboo | Airborne/endophyte | China | KY494708 | KY494784 | KY705177 | KY705107 |
| Ap. guizhouensis | LC 5322T | Air in karst cave, bamboo | Airborne/endophyte | China | KY494709 | KY494785 | KY705178 | KY705108 |
| Ap. hainanensis | SAUCC 1681T | Leaf of bamboo | Pathogen | China | OP563373 | OP572422 | OP573268 | OP573262 |
| Ap. hainanensis | SAUCC 1682 | Leaf of bamboo | Pathogen | China | OP563372 | OP572421 | OP573267 | OP573261 |
| Ap. hispanica | IMI 326877T | Beach sand | Saprobe | Spain | AB220242 | AB220336 | AB220289 | - |
| Ap. hydei | CBS 114990T | Culms of Bambusa tuldoides | Saprobe | Hong Kong, China | KF144890 | KF144936 | KF144982 | KF145024 |
| Ap. hydei | KUMCC 16-0204 | Bambusa tuldoides | Saprobe | China | KY356087 | KY356092 | - | - |
| Ap. hyphopodii | MFLUCC 15-0003T | Bambusa tuldoides | Saprobe | China | KR069110 | - | - | - |
| Ap. hyphopodii | KUMCC 16-0201 | Bambusa tuldoides | Saprobe | China | KY356088 | KY356093 | - | - |
| Ap. hysterina | ICPM 6889T | Bamboo | Saprobe | New Zealand | MK014874 | MK014841 | MK017980 | MK017951 |
| Ap. hysterina | CBS 145133 | Bamboo | Saprobe | New Zealand | MK014875 | MK014842 | MK017981 | MK017952 |
| Ap. iberica | CBS 145137T | Arundo donax | Saprobe | Portugal | MK014879 | MK014846 | MK017984 | MK017955 |
| Ap. intestini | CBS 135835T | Gut of a grasshopper | Saprobe | India | KR011352 | MH877577 | KR011350 | KR011351 |
| Ap. intestini | MFLUCC 21-0052 | Gut of a grasshopper | Saprobe | India | MZ542521 | MZ542525 | MZ546410 | MZ546406 |
| Ap. italica | CBS 145138T | Arundo donax | Saprobe | Italy | MK014880 | MK014847 | MK017985 | MK017956 |
| Ap. italica | CBS 145139 | Arundo donax | Saprobe | Italy | MK014881 | MK014848 | MK017986 | - |
| Ap. jatrophae | AMH-9557T | Jatropha podagrica | Saprobe | India | JQ246355 | - | - | - |
| Ap. jatrophae | AMH-9556 | Jatropha podagrica | Saprobe | India | HE981191 | - | - | - |
| Ap. jiangxiensis | LC 4494 | Maesa sp. | Endophyte | China | KY494690 | KY494766 | KY705160 | KY705089 |
| Ap. jiangxiensis | LC 4577T | Maesa sp. | Endophyte | China | KY494693 | KY494769 | KY705163 | KY705092 |
| Ap. kogelbergensis | CBS 113332 | Dead culms of Restionaceae | Saprobe | South Africa | KF144891 | KF144937 | KF144983 | KF145025 |
| Ap. kogelbergensis | CBS 113333T | Dead culms of Restionaceae | Saprobe | South Africa | KF144892 | KF144938 | KF144984 | KF145026 |
| Ap. koreana | KUC21332T | Egg of Arctoscopus japonicus | Not mentioned | Republic of Korea | MH498524 | - | MH498482 | MH544664 |
| Ap. koreana | KUC21348 | Egg of Arctoscopus japonicus | Not mentioned | Republic of Korea | MH498523 | - | MH498481 | MN868927 |
| Ap. lageniformis | KUC21686T | Branch of Phyllostachys pubescens | Not mentioned | Republic of Korea | ON764022 | ON787761 | ON806636 | ON806626 |
| Ap. lageniformis | KUC21687 | Branch of Phyllostachys pubescens | Not mentioned | Republic of Korea | ON764023 | ON787762 | ON806637 | ON806627 |
| Ap. locuta-pollinis | LC 11688 | Brassica campestris | Saprobe | China | MF939596 | - | MF939623 | MF939618 |
| Ap. locuta-pollinis | LC 11683T | Brassica campestris | Saprobe | China | MF939595 | - | MF939622 | MF939616 |
| Ap. longistroma | MFLUCC 11-0479 | Dead culms of bamboo | Saprobe | Thailand | KU940142 | KU863130 | - | - |
| Ap. longistroma | MFLUCC 11-0481T | Dead culms of bamboo | Saprobe | Thailand | KU940141 | KU863129 | - | - |
| Ap. magnispora | ZHKUCC 22-0001 | Bamboo | Saprobe | China | OM728647 | OM486971 | OM0543544 | OM543543 |
| Ap. malaysiana | CBS 102053T | Macaranga hullettii | Saprobe | Malaysia | KF144896 | KF144942 | KF144988 | KF145030 |
| Ap. marianiae | CBS 148710T | Phleum pratense | Saprobe | Spain | NR_183001 | NG_149092 | - | |
| Ap. marianiae | AP301119 | Phleum pratense | Saprobe | Spain | ON692407 | ON692423 | ON677187 | ON677181 |
| Ap. marii | CBS 497.90T | Beach sands | Saprobe | Spain | AB220252 | KF144947 | KF144993 | KF145035 |
| Ap. marii | DiSSPA_A1 | Beach sands | Saprobe | Spain | MK602320 | - | MK614695 | MK645472 |
| Ap. marina | KUC21328T | Seaweed | Not mentioned | Republic of Korea | MH498538 | - | MH498496 | MH544669 |
| Ap. marina | KUC21353 | Seaweed | Not mentioned | Republic of Korea | MH498537 | - | MH498495 | MN868923 |
| Ap. mediterranea | IMI 326875T | Air | Saprobe | Spain | AB220243 | AB220337 | AB220290 | - |
| Ap. minutispora | 1.70-41 | Mountain soil | Soil | Republic of Korea | LC517882 | - | LC518888 | LC518889 |
| Ap. mori | MFLUCC 20-0181T | Morus australis | Saprobe | Taiwan | MW114313 | MW114393 | - | - |
| Ap. mori | NCYUCC 19-034 | Morus australis | Saprobe | Taiwan | MW114314 | MW114394 | - | - |
| Ap. mukdahanensis | MFLUCC 22-0056T | dead bamboo leave | Saprobe | Thailand | OP377735 | OP377742 | - | OP381089 |
| Ap. multiloculata | MFLUCC 21-0023T | Dead bamboo | Saprobe | Thailand | OL873137 | OL873138 | - | - |
| Ap. mytilomorpha | DAOM 214595T | Andropogon sp. | Saprobe | India | KY494685 | - | - | - |
| Ap. neobambusae | LC 7106T | Leaves of bamboo | Saprobe/endophyte | China | KY494718 | KY494794 | KY705186 | KY806204 |
| Ap. neobambusae | LC 7124 | Leaves of bamboo | Saprobe/endophyte | China | KY494727 | KY494803 | KY705195 | KY806206 |
| Ap. neochinensis | CFCC 53036T | Fargesia qinlingensis | Saprobe | China | MK819291 | - | MK818547 | MK818545 |
| Ap. neochinensis | CFCC 53037 | Fargesia qinlingensis | Saprobe | China | MK819292 | - | MK818548 | MK818546 |
| Ap. neogarethjonesii | KUMCC 18-0192 | Bamboo | Saprobe | China | MK070897 | MK070898 | - | - |
| Ap. neosubglobosa | JHB 006 | Bamboo | Saprobe | China | KY356089 | KY356094 | - | - |
| Ap. neosubglobosa | KUMCC 16-0203T | Bamboo | Saprobe | China | KY356090 | KY356095 | - | - |
| Ap. obovata | LC 4940T | Lithocarpus sp. | Endophyte | China | KY494696 | KY494772 | KY705166 | KY705095 |
| Ap. obovata | LC 8177 | Lithocarpus sp. | Endophyte | China | KY494757 | KY494833 | KY705225 | KY705153 |
| Ap. ovata | CBS 115042T | Arundinaria hindsii | Saprobe | China | KF144903 | KF144950 | KF144995 | KF145037 |
| Ap. paraphaeosperma | MFLUCC 13-0644T | Dead culms of bamboo | Saprobe | Thailand | KX822128 | KX822124 | - | - |
| Ap. phragmitis | CPC 18900T | Phragmites australis | Saprobe | Italy | KF144909 | KF144956 | KF145001 | KF145043 |
| Ap. phyllostachydis | MFLUCC 18-1101T | Phyllostachys heteroclada | Saprobe | China | MK351842 | MH368077 | MK291949 | MK340918 |
| Ap. piptatheri | CBS 145149T | Piptatherum miliaceum | Saprobe | Spain | MK014893 | MK014860 | - | MK017969 |
| Ap. pseudohyphopodii | KUC21680T | Culm of Phyllostachys pubescens | Not mentioned | Republic of Korea | ON764026 | ON787765 | ON806640 | ON806630 |
| Ap. pseudohyphopodii | KUC21684 | Culm of Phyllostachys pubescens | Not mentioned | Republic of Korea | ON764027 | ON787766 | ON806641 | ON806631 |
| Ap. pseudoparenchymatica | LC 7234T | Leaves of bamboo | Endophyte | China | KY494743 | KY494819 | KY705211 | KY705139 |
| Ap. pseudoparenchymatica | LC 8173 | Leaves of bamboo | Endophyte | China | KY494753 | KY494829 | KY705221 | KY705149 |
| Ap. pseudorasikravindrae | KUMCC 20-0208T | Bambusa dolichoclada | Saprobe | China | MT946344 | - | MT947367 | MT947361 |
| Ap. pseudosinensis | CPC 21546T | Leaves of bamboo | Saprobe | Netherlands | KF144910 | KF144957 | - | KF145044 |
| Ap. pseudospegazzinii | CBS 102052T | Macaranga hullettii | Saprobe | Malaysia | KF144911 | KF144958 | KF145002 | KF145045 |
| Ap. pterosperma | CBS 123185 | Lepidosperma gladiatum | Saprobe | Australia | KF144912 | KF144959 | KF145003 | - |
| Ap. pterosperma | CPC 20193T | Lepidosperma gladiatum | Saprobe | Australia | KF144913 | KF144960 | KF145004 | KF145046 |
| Ap. pusillisperma | KUC21321T | Seaweed | Not mentioned | Republic of Korea | MH498533 | - | MH498491 | MN868930 |
| Ap. pusillisperma | KUC21357 | Seaweed | Not mentioned | Republic of Korea | MH498532 | - | MH498490 | MN868931 |
| Ap. qinlingensis | CFCC 52303T | Fargesia qinlingensis | Saprobe | China | MH197120 | - | MH236791 | MH236795 |
| Ap. qinlingensis | CFCC 52304 | Fargesia qinlingensis | Saprobe | China | MH197121 | - | MH236792 | MH236796 |
| Ap. rasikravindrae | LC 8179 | Brassica rapa | Saprobe | China | KY494759 | KY494835 | KY705227 | KY705155 |
| Ap. rasikravindrae | NFCCI 2144T | Soil | Saprobe | Norway | JF326454 | - | - | - |
| Ap. rasikravindrae | MFLUCC 21-0051 | Dead culms of bamboo | Saprobe | Thailand | MZ542523 | MZ542527 | MZ546412 | MZ546408 |
| Ap. rasikravindrae | MFLUCC 21-0054 | Dead culms of Maize | Saprobe | Thailand | MZ542522 | MZ542526 | MZ546411 | MZ546407 |
| Ap. sacchari | CBS 372.67 | Air | Endophyte | - | KF144918 | KF144964 | KF145007 | KF145049 |
| Ap. sacchari | CBS 664.74 | Soil under Calluna vulgaris | Endophyte | Netherlands | KF144919 | KF144965 | KF145008 | KF145050 |
| Ap. saccharicola | CBS 191.73 | Air | Endophyte | Netherlands | KF144920 | KF144966 | KF145009 | KF145051 |
| Ap. saccharicola | CBS 831.71 | - | Endophyte | Netherlands | KF144922 | KF144969 | KF145012 | KF145054 |
| Ap. sargassi | KUC21228T | Sargassum fulvellum | Not mentioned | Republic of Korea | KT207746 | - | KT207644 | MH544677 |
| Ap. sargassi | KUC21232 | Sargassum fulvellum | Not mentioned | Republic of Korea | KT207750 | - | KT207648 | MH544676 |
| Ap. sasae | CBS 146808T | dead culms | Saprobe | Netherlands | MW883402 | MW883797 | MW890120 | MW890104 |
| Ap. septata | CGMCC 3.20134T | bamboo | Saprobe | China | MW481711 | MW478890 | MW522960 | MW522943 |
| Ap. septata | GZCC 20-0109 | bamboo | Saprobe | China | MW481712 | MW478891 | MW522961 | MW522944 |
| Ap. serenensis | IMI 326869T | excipients, atmosphere andhome dust | Saprobe | Spain | AB220250 | AB220344 | AB220297 | - |
| Ap. setariae | MT492005 | Setaria viridis | Saprobe | China | MT492005 | - | MT497467 | MW118457 |
| Ap. setostroma | KUMCC 19-0217T | Dead branches of bamboo | Saprobe | China | MN528012 | MN528011 | - | MN527357 |
| Ap. sichuanensis | HKAS 107008T | dead culm of grass | Saprobe | China | MW240648 | MW240578 | MW775605 | MW759536 |
| Ap. sorghi | URM 93000T | Sorghum bicolor | Endophyte | Brazil | MK371706 | - | MK348526 | - |
| Ap. sp. | ZHKUCC 23-0010 | Wurfbainia villosa | Endophyte | China | OQ588000 | OQ587988 | OQ586066 | OQ586079 |
| Ap. sp. | ZHKUCC 23-0011 | Wurfbainia villosa | Endophyte | China | OQ588001 | OQ587989 | OQ586067 | OQ586080 |
| Ap. sp. | ZHKUCC 23-0012 | Wurfbainia villosa | Endophyte | China | OQ588002 | OQ587990 | OQ586068 | OQ586081 |
| Ap. sp. | ZHKUCC 23-0013 | Wurfbainia villosa | Endophyte | China | OQ588003 | OQ587991 | OQ586069 | OQ586082 |
| Ap. stipae | CBS 146804T | dead culm of Stipa gigantea | Saprobe | Spain | MW883403 | MW883798 | MW890121 | MW890082 |
| Ap. subglobosa | MFLUCC 11-0397T | Dead culms of bamboo | Saprobe | Thailand | KR069112 | KR069113 | - | - |
| Ap. subrosea | LC 7291 | Leaves of bamboo | Endophyte | China | KY494751 | KY494827 | KY705219 | KY705147 |
| Ap. subrosea | LC 7292T | Leaves of bamboo | Endophyte | China | KY494752 | KY494828 | KY705220 | KY705148 |
| Ap. taeanensis | KUC21322T | Seaweed | Not mentioned | Republic of Korea | MH498515 | - | MH498473 | MH544662 |
| Ap. taeanensis | KUC21359 | Seaweed | Not mentioned | Republic of Korea | MH498513 | - | MH498471 | MN868935 |
| Ap. thailandica | MFLUCC 15-0199 | Dead culms of bamboo | Saprobe | Thailand | KU940146 | KU863134 | - | - |
| Ap. thailandica | MFLUCC 15-0202T | Dead culms of bamboo | Saprobe | Thailand | KU940145 | KU863133 | - | - |
| Ap. tropica | MFLUCC 21-0056T | Dead culms of bamboo | Saprobe | Thailand | OK491657 | OK491653 | OK560922 | - |
| Ap. vietnamensis | IMI 99670T | Citrus sinensis | Saprobe | Vietnam | KX986096 | KX986111 | KY019466 | - |
| Ap. wurfbainiae | ZHKUCC 23-0008T | Wurfbainia villosa | Endophyte | China | OQ587998 | OQ587986 | OQ586064 | OQ586077 |
| Ap. wurfbainiae | ZHKUCC 23-0009 | Wurfbainia villosa | Endophyte | China | OQ587999 | OQ587987 | OQ586065 | OQ586078 |
| Ap. xenocordella | CBS 478.86T | Soil from roadway | Soil | Zimbabwe | KF144925 | KF144970 | KF145013 | KF145055 |
| Ap. xenocordella | CBS 595.66 | On dead branches | Saprobe | Misiones | KF144926 | KF144971 | - | - |
| Ap. yunnana | DDQ 00281 | Phyllostachys nigra | Saprobe | China | KU940148 | KU863136 | - | - |
| Ap. yunnana | MFLUCC 15-1002T | Phyllostachys nigra | Saprobe | China | KU940147 | KU863135 | - | - |
| Ap. yunnanensis | ZHKUCC 23-0014T | Grass | Saprobe | China | OQ588004 | OQ587992 | OQ586070 | OQ586083 |
| Ap. yunnanensis | ZHKUCC 23-0015 | Grass | Saprobe | China | OQ588005 | OQ587993 | OQ586071 | OQ586084 |
| Arthrinium austriacum | GZU 345004 | Carex pendula | Saprobe | Austria | MW208928 | - | - | - |
| Ar. austriacum | GZU 345006 | Carex pendula | Saprobe | Austria | MW208929 | MW208860 | - | - |
| Ar. sporophleum | GZU 345102 | Carex firma | Saprobe | Austria | MW208944 | MW208866 | - | - |
| Ar. caricicola | CBS 145127 | Carex ericetorum | Saprobe | China | MK014871 | MK014838 | MK017977 | MK017948 |
| Ar. crenatum | AG19066T | Dead leaves of grass (probably Festuca burgundiana) | Saprobe | France | MW208931 | MW208861 | - | - |
| Ar. curvatum | AP 25418 | Leaves of Carex sp. | Saprobe | China | MK014872 | MK014839 | MK017978 | MK017949 |
| Ar. japonicum | IFO 30500 | - | Saprobe | Japan | AB220262 | AB220356 | AB220309 | - |
| Ar. japonicum | IFO 31098 | Leaves of Carex despalata | Saprobe | Japan | AB220264 | AB220358 | AB220311 | - |
| Ar. luzulae | AP7619-3T | Luzula sylvatica | Saprobe | Spain | MW208937 | MW208863 | - | - |
| Ar. morthieri | GZU 345043 | Carex pilosa | Saprobe | Austria | MW208938 | MW208864 | - | - |
| Ar. phaeospermum | CBS 114317 | Leaves of Hordeum vulgare | Saprobe | Iran | KF144906 | KF144953 | KF144998 | KF145040 |
| Ar. phaeospermum | CBS 114318 | Leaves of Hordeum vulgare | Saprobe | Iran | KF144907 | KF144954 | KF144999 | KF145041 |
| Ar. puccinioides | CBS 549.86 | Lepidosperma gladiatum | Saprobe | Germany | AB220253 | AB220347 | AB220300 | - |
| Ar. sphaerospermum | CBS 146355 | Probably on Poaceae | Saprobe | Norway | MW208943 | MW208865 | - | - |
| Ar. sporophleum | CBS 145154 | Dead leaves of Juncus sp. | Saprobe | Spain | MK014898 | MK014865 | MK018001 | MK017973 |
| Ar. trachycarpum | CFCC 53039 | Trachycarpus fortune | Pathogen | China | MK301099 | - | MK303395 | MK303397 |
| Ar. urticae | IMI 326344 | - | Saprobe | - | AB220245 | AB220339 | AB220292 | - |
| Nigrospora aurantiaca | CGMCC 3.18130T | Nelumbo sp. | Saprobe | China | KX986064 | KX986098 | KY019465 | KY019295 |
| N. camelliae-sinensis | CGMCC 3.18125T | Camellia sinensis | Endophyte/pathogen | China | KX985986 | KX986103 | KY019460 | KY019293 |
| N. chinensis | CGMCC 3.18127T | Machilus breviflora | Endophyte/pathogen | China | KX986023 | KX986107 | KY019462 | KY019422 |
| N. gorlenkoana | CBS 480.73 | Vitis vinifera | Endophyte/pathogen | Kazakhstan | KX986048 | KX986109 | KY019456 | KY019420 |
| N. guilinensis | CGMCC 3.18124T | Camellia sinensis | Endophyte/pathogen | China | KX985983 | KX986113 | KY019459 | KY019292 |
| N. hainanensis | CGMCC 3.18129T | Musa paradisiaca | Endophyte/pathogen | China | KX986091 | KX986112 | KY019464 | KY019415 |
| N. lacticolonia | CGMCC 3.18123T | Camellia sinensis | Endophyte/pathogen | China | KX985978 | KX986105 | KY019458 | KY019291 |
| N. musae | CBS 319.34 | Musa sp. | Endophyte/pathogen | Australia | MH855545 | KX986110 | KY019455 | KY019419 |
| N. oryzae | LC2693 | Neolitsea sp. | Saprobe | China | KX985944 | KX986101 | KY019471 | KY019299 |
| N. osmanthi | CGMCC 3.18126T | Hedera nepalensis | Endophyte/pathogen | China | KX986010 | KX986106 | KY019461 | KY019421 |
| N. pyriformis | CGMCC 3.18122T | Citrus sinensis | Endophyte/pathogen | China | KX985940 | KX986100 | KY019457 | KY019290 |
| N. rubi | LC2698T | Rubus sp. | Endophyte/pathogen | China | KX985948 | KX986102 | KY019475 | KY019302 |
| N. sphaerica | LC7298 | Nelumbo sp. | Saprobe | China | KX985937 | KX986097 | KY019606 | KY019401 |
| N. vesicularis | CGMCC 3.18128T | Musa paradisiaca | Endophyte | China | KX986088 | KX986099 | KY019463 | KY019294 |
| Sporocadus trimorphus | CFCC 55171T | Rose | Not mentioned | China | OK655798 | OK560389 | OM401677 | OL814555 |
| S. trimorphus | ROC 113 | Rose | Not mentioned | China | OK655799 | OK560390 | OM401678 | OL814556 |
| Characters | Apiospora Species | |||
|---|---|---|---|---|
| Ap. endophytica | Ap. hydei | Ap. cordylines | Ap. aurea | |
| Host/substrate | Asymptomatic leaf of Wurfbainia villosa | Culms of Bambusa tuldoides | Leaves of Cordyline fruticosa | Air |
| Conidiophores | Reduced to conidiogenous cells | Pale brown, smooth, subcylindrical, transversely septate, branched, 20–40 × 3–5 μm | NA | NA |
| Conidiogenous cells | Aggregated in clusters or solitary, hyaline to golden brown, smoothly, erect, unbranched, cylindrical or clavate, ampulliform or obtriangular, 4–14 × 2–7 μm ( = 7.5 × 5 μm) | Aggregated in clusters, brown, smooth, subcylindrical to doliiform to lageniform, 5–8 × 4–5 μm | Erect, aggregated into clusters, hyaline to pale brown, smooth, doliiform to ampulliform or lageniform, (3–)5–10(–15) × 2.6–5.3 µm ( = 7.0 × 4.5 µm) | Integrated, polyblastic, denticulate |
| Conidia | Initially hyaline, becoming pale brown to dark brown, globose to subglobose, obovoid to ellipsoidal in the face view, lenticular with a thick equatorial slit in the side view, smooth-walled, 14–19 × 12–18 μm ( = 17 × 15 μm, n = 30) in the face view, 11–19 × 9–16 μm ( = 15 × 12 μm, n = 20) | Brown, roughened, globose in face view, lenticular in the side view, with pale equatorial slit, (15–)17– 19(–22) μm diam. in face view, (10–)11–12(–14) μm diam. in the side view, with a central scar, 1.5–2 μm diam. | Olivaceous to brown, smooth to finely roughened, subglobose to ellipsoidal, 15–19 × 12.5–18.5 µm ( = 17.5 × 15.7 µm) | Solitary, terminal, and sometimes also lateral with a hyaline rim, brown or dark brown, smooth, aseptate, 10–30 × 10–15 μm |
| Reference | This study | [8] | [49] | [50] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.; Senanayake, I.C.; Dong, W.; Thilini Chethana, K.W.; Tangtrakulwanich, K.; Zhang, Y.; Doilom, M. Taxonomic and Phylogenetic Updates on Apiospora: Introducing Four New Species from Wurfbainia villosa and Grasses in China. J. Fungi 2023, 9, 1087. https://doi.org/10.3390/jof9111087
Liao C, Senanayake IC, Dong W, Thilini Chethana KW, Tangtrakulwanich K, Zhang Y, Doilom M. Taxonomic and Phylogenetic Updates on Apiospora: Introducing Four New Species from Wurfbainia villosa and Grasses in China. Journal of Fungi. 2023; 9(11):1087. https://doi.org/10.3390/jof9111087
Chicago/Turabian StyleLiao, Chunfang, Indunil Chinthani Senanayake, Wei Dong, Kandawatte Wedaralalage Thilini Chethana, Khanobporn Tangtrakulwanich, Yunxia Zhang, and Mingkwan Doilom. 2023. "Taxonomic and Phylogenetic Updates on Apiospora: Introducing Four New Species from Wurfbainia villosa and Grasses in China" Journal of Fungi 9, no. 11: 1087. https://doi.org/10.3390/jof9111087