


Palm Fungi and Their Key Role in Biodiversity Surveys: A Review

Biosystems and Integrative Sciences Institute (BioISI), Faculdade de Ciências, Universidade de Lisboa, Campo Grande, 1749-016 Lisboa, Portugal
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(11), 1121; https://doi.org/10.3390/jof9111121
Submission received: 15 October 2023
/
Revised: 16 November 2023
/
Accepted: 17 November 2023
/
Published: 19 November 2023
(This article belongs to the Special Issue Taxonomy, Systematics and Evolution of Forestry Fungi)
Abstract
:Over the past three decades, a wealth of studies has shown that palm trees (Arecaceae) are a diverse habitat with intense fungal colonisation, making them an important substratum to explore fungal diversity. Palm trees are perennial, monocotyledonous plants mainly restricted to the tropics that include economically important crops and highly valued ornamental plants worldwide. The extensive research conducted in Southeast Asia and Australasia indicates that palm fungi are undoubtedly a taxonomically diverse assemblage from which a remarkable number of new species is continuously being reported. Despite this wealth of data, no recent comprehensive review on palm fungi exists to date. In this regard, we present here a historical account and discussion of the research on the palm fungi to reflect on their importance as a diverse and understudied assemblage. The taxonomic structure of palm fungi is also outlined, along with comments on the need for further studies to place them within modern DNA sequence-based classifications. Palm trees can be considered model plants for studying fungal biodiversity and, therefore, the key role of palm fungi in biodiversity surveys is discussed. The close association and intrinsic relationship between palm hosts and palm fungi, coupled with a high fungal diversity, suggest that the diversity of palm fungi is still far from being fully understood. The figures suggested in the literature for the diversity of palm fungi have been revisited and updated here. As a result, it is estimated that there are about 76,000 species of palm fungi worldwide, of which more than 2500 are currently known. This review emphasises that research on palm fungi may provide answers to a number of current fungal biodiversity challenges.
1. Introduction
Arecaceae (syn. Palmae), colloquially known as palm trees, is one of the best known and most extensively cultivated plant families, comprising around 2600 species in 181 genera [1]. Palms are important plants in terms of human exploitation in their native range. Some species, such as oil (Elaeis species), coconut (Cocos nucifera), rattan (Calamus species), and date (Phoenix dactylifera) palms, are extremely important in the international trade [2,3,4]. Moreover, palms are highly prized as ornamentals due to their great decorative potential. Thus, although almost exclusively native to tropical or sub-tropical countries, these perennial monocotyledonous trees are currently distributed worldwide due to their use as ornamental plants [4,5]. Therefore, palm trees have become a distinctive component of the urban landscape and an important accessory in interior decoration and in floristry.
Over the last 30 years, a considerable number of studies have shown the association of a diverse range of fungi with palm tissues. The extensive research carried out in Southeast Asia and Australasia showed that palms are a rich source of previously unknown fungal taxa. Thus, many species and genera were formally identified and described as new to science based on palm collections, e.g., refs. [6,7,8]. The comprehensive isolation of fungi from palm tissues has proven that these fungi are an important and taxonomically diverse assemblage that is often referred to as palm fungi or palmicolous fungi, e.g., ref. [9]. While most studies on palm fungi have focused on systematic and descriptive taxonomy, a few studies have explored the biodiversity and ecology of these microfungi, e.g., refs. [10,11,12,13,14,15,16]. Taxonomically, palm fungi are one of the most diverse groups of fungi. The currently established figure is that more than 1500 species of fungi have been described from palm hosts, with representatives from almost all major fungal classes [17]. In earlier studies, most taxa were introduced, described, and arranged in different taxonomic ranks within the Ascomycota based on morphological analyses. This approach was, however, subjective and many taxa were wrongly assigned or assigned to the Ascomycota genera incertae sedis.
Despite the wealth of data on the assemblage of fungi that inhabit palm tissues, there has been no recent comprehensive review of palm fungi to date. In this regard, the present review aims to give a historical overview on the studies that have been performed on palm fungi and reflect on their importance as a diverse and understudied assemblage. Moreover, two main aspects will be presented and discussed: (1) what the taxonomic structure of palm fungi is, along with comments on the need for further studies to place them within modern DNA sequence-based classifications; (2) how palm trees can be regarded as model plants for studying fungal biodiversity, given the key role of palm fungi in biodiversity surveys.
2. Historical Account of Research on Palm Fungi and Reflections on Their Importance
The history of the study of palm fungi can be divided into three distinct periods. The first, between 1880 and 1920, includes the publications of classical mycologists, such as Hennings, Rehm, Penzig, Spegazzini, H. Sydow and P. Sydow, and Saccardo [18,19,20,21,22,23,24,25,26,27]. Although many fungal species were described from palm trees during this period, these publications are barely accessible or legible since most of them were written in Latin. The second, between 1920 and 1990, includes several mycologists who, for 70 years, occasionally reported the occurrence of fungi on palm tissues. However, almost no study on palm fungi in this period was very extensive. The third, which began in the 1990s and continues today, represents the first time that a group of mycologists has dedicated itself to specifically analysing the microfungi that occur on palm trees. This includes the research carried out by Hyde, his students, and colleagues, which represents not only a significant advance in the number of fungal species described on palm substrata, but also a considerable advance in the biodiversity of palm fungi.
2.1. History of Systematic and Descriptive Taxonomy Studies on Palm Fungi
There are numerous reports scattered throughout the literature on fungi collected from different parts of different species of palm trees and from different regions of the world. An overwhelming number of these studies have been dedicated to collecting and describing fungi that inhabit palm tissues, including new genera and new species. Although there are several reports from before the 1990s, the present overview focuses on the extensive studies carried out by Hyde and co-workers. Hyde’s research is the first body of theory to understand palm fungi as a fungal community with taxonomic characteristics, biological processes, and general diversity that is worth exploring in depth. Most of the descriptions prior to Hyde’s research, especially those from the early 20th century, consist of short Latin paragraphs that lack illustrations and/or give limited information about the identity of the fungi.
2.1.1. From Scattered to the First Systematic Reports on Palm Fungi
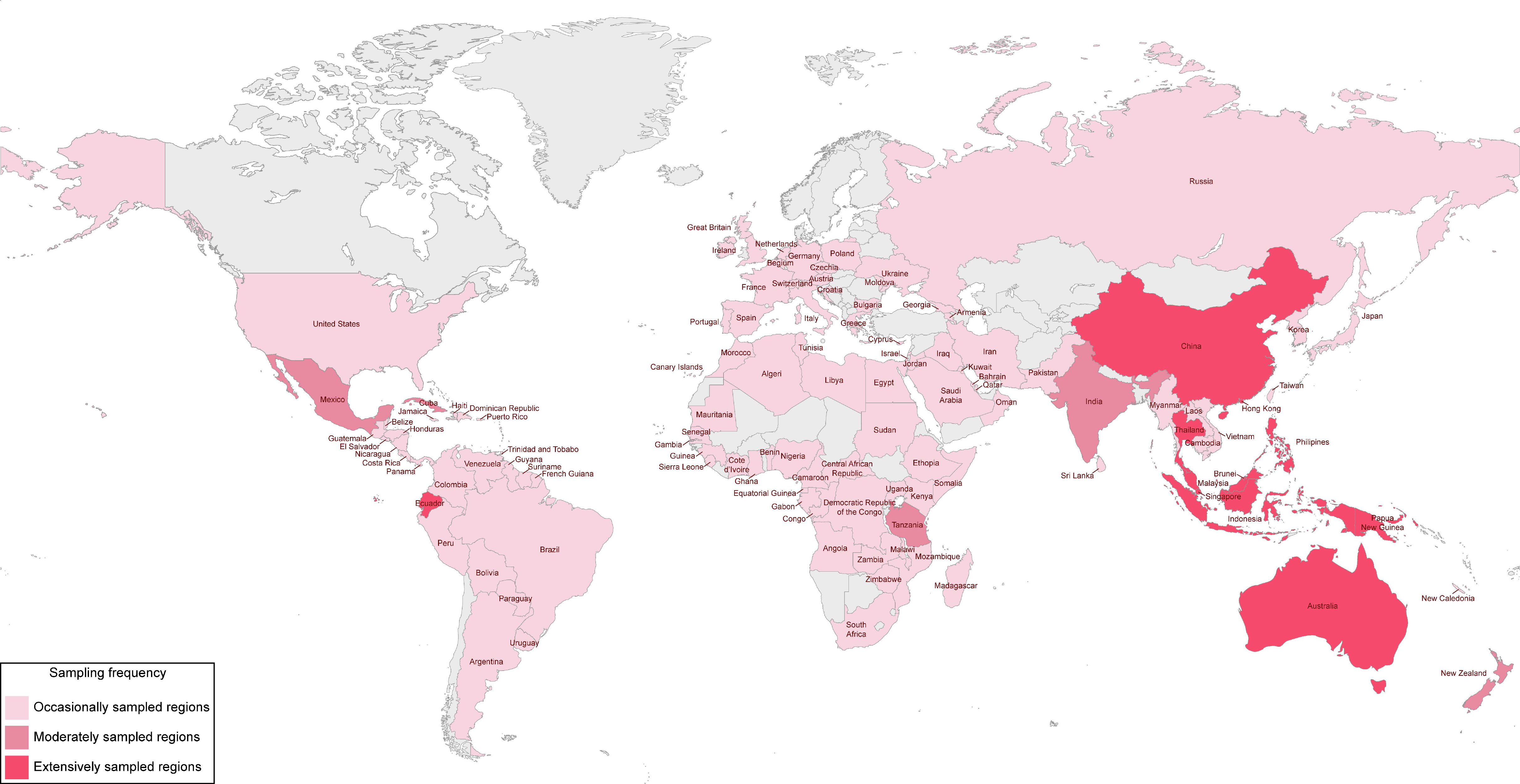
Before the extensive studies carried out by Hyde and co-workers, there were several occasional reports of fungi collected from different palms and tissues, such as leaf litter, green foliage, rachides, trunks, and stems. Although these collections come from different countries around the world, most of them come from tropical and subtropical regions where palm trees are almost exclusively native. A summary of some examples of these studies is given here considering the last decades of the 20th century. The sampling regions around the world where palm fungi have been recorded are mapped in Figure 1, according to the available literature.
Collections of palm species native to tropical regions of Central and South America and West and Southeast Africa (Figure 1) have often yielded taxonomic novelties, including from new genera and species to new records on new hosts and new geographical distributions. For instance, Ellis [28] reported new Lacellina species on dead leaves of Borassus aethiopum from Ghana and Sierra Leone (West Africa). Later, Ellis also reported new species of palm fungi from Central America, West Africa, and Malaysia on his series of papers and books concerning “Dematiaceous Hyphomycetes” [29,30,31,32].
Southeast Asia, Australasia, and India were also frequently surveyed (Figure 1) and the first two would later become major regions for palm fungi investigation. For instance, while studying leaf-spotting hyphomycetes, Deighton [33,34] reported the new species Cercospora raphiae and Pseudocercospora carpentariae on leaves of Raphia farinifera from Zimbabwe (Southeast Africa) and Carpentaria acuminata from Australia. In the same year, several Xylariaceae palmicolous fungi were recorded on the rainforests of North Sulawesi (Indonesia) by Rogers et al. [35].
Several other regions were also only occasionally surveyed (Figure 1), including, for instance, Venezuela (South America), where some palmicolous fungi, including new species, were described in the series of papers “Fungi venezuelani” [36,37,38]; Argentina (South America), where Wright [39] recorded the new genus and species Agaricostilbum palmicola (currently synonymised under Sterigmatomyces pulcherrimus) on weathered spathes of Butia yatay and Phoenix sp.; and Japan (East Asia), where Hino and Katumoto [40,41,42] described some palmicolous fungi, including new species, in the series of papers entitled “Notes on fungi from western Japan”. Less frequently, the USA, European countries, and China, where only a few palm species are native, have also been the stage for collecting palm fungi (Figure 1). For instance, several helicosporous fungi collected from palm tissues were reported from Hawaii (USA) by Goos [43]; the new phialosporus hyphomycetes genus and species Craspedodidymum elatum were reported on rotten petioles of P. canariensis from Czech Republic (Central Europe) by Holubová-Jechová [44]; and the new species Zasmidium caryotae (as Stenella caryotae) have been reported on the leaves of Caryota mitis from Guangdong (China) by Liu and Liao [45].
These first scattered reports of palmicolous fungi have established their importance as a highly diverse and understudied community that can be accessed on palms all over the world (Figure 1). Some reports have expanded the geographical distribution of these fungi, which was important for the establishment of the geographical and ecological range of some important phytopathogens. For example, Samuels and Rossman [46] during their studies on the Amphisphaeriaceae-recorded Leiosphaerella cocoes on fronds and fruits of Cocos nucifera from several different regions, namely Dominican Republic, Guam, Indonesia, Mexico, USA, Tonga, Cook Islands, and Papua New Guinea, and reported two new species of Oxydothis, O. rhopalostylidis and O. selenosporellae, on the leaf midrib of Rhopalostylis sapida from New Zealand. It is worth mentioning that New Zealand was one of the first countries where several reports of palmicolous fungi were published more systematically, most of them from collections of the endemic palm tree R. sapida (nikau palm), which is the only palm native to mainland New Zealand. Thus, several palm fungi, including many new species, were published in the series of papers “New Zealand Fungi” by Hughes [47,48,49,50,51,52,53], as well as in many other occasional publications that were mostly later summarised by McKenzie et al. [54] in their checklist of fungi recorded on nikau palm from New Zealand.
Two of the first most extensive studies on palm fungi include those of Pirozynski and Matsushima. Pirozynski [55] reported forty-six species, including the new genus Bondiella, six new species, and some new combinations of fungi from the oil palm E. guineensis collected from Tanzania. Matsushima [56,57,58,59,60,61,62,63,64,65,66,67] and Matsushima and Matsushima [68,69] reported more than 300 fungi from palm litter, including 8 new genera, viz. Apogaeumannomyces [67], Atrosetaphiale [65], Hyalobelemnospora, Paradactylella [64], Setophiale, Venustocephala [65], Veramyces [64], Verticimonosporium [56], and more than 80 new species, mostly of rare and interesting hyphomycetous fungi. Most of these fungi were recorded in collections of palms from different regions of the world, from the Americas to Australia and Eastern Asia, including Peru, Guam, Taiwan, Cuba, Northern Queensland, and Ecuador (Figure 1), were compiled in the classic book series “Matsushima mycological memoirs”.
Castañeda-Ruiz, Holubová-Jechová, Mena-Portales, and Mercado-Sierra were one of the first groups of mycologists to report several species of palm fungi growing on dead and decaying palm tissues, such as trunks, rachides, and petioles. Although these reports were abundant and consistent, their main purpose was not to report on the palmicolous fungi themselves, i.e., to understand the taxonomic or ecological structure of palm fungal communities. They were part of an investigation of hyphomycetes of Cuba, where many new species were introduced, e.g., refs. [70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85]. Most of these reports were summarised and discussed in series of papers, such as “Hifomicetes demaciáceos de Cuba” [86,87], “Studies on hyphomycetes from Cuba” [88,89,90,91,92,93,94,95], and “Nuevos o raros hifomicetes de Cuba” [96,97,98,99,100,101,102], and books, such as “Hifomicetes demaciáceos de Sierra del Rosario, Cuba” [103]. Although several of these reports were from undetermined palm trees and other particular known species, most of them were from the Cuban royal palm Roystonea regia, where several new species were described, insomuch that Mercado-Sierra [73] has described R. regia as an “ideal substratum for the development of dematiaceous hyphomycetes”.
2.1.2. Hyde and Co-Workers and the Extensive Studies on Palm Fungi from Tropical Regions
The last 30 years has seen an extensive profusion of studies regarding palmicolous fungi by Hyde and co-workers. The extent and depth of their investigation yielded an impressive body of literature that made it possible to begin to understand and characterise the taxonomy of palm fungi, particularly the communities inhabiting tropical and subtropical palms growing on their native regions. These systematic studies culminated in the publication of three books entitled “Genera of ascomycetes from palms” [104], “Palm microfungi” [6], and “Microfungi of tropical and temperate palms” [8], and a series of publications entitled “Fungi from palms”, comprising 49 papers where numerous new fungi to science were described [105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153]. Genera that have been described as new to science and found on palm trees over the last three decades are summarised in Table 1. Genera and respective families in subclasses of Dothideomycetes and Sordariomycetes with common representatives found on palm trees are summarised in Table 2.
Hyde et al. have been studying microfungi from palms since 1988 when new species of the genera Linocarpon, Oxydothis, and Astrosphaeriella (as Trematosphaeria) were described from the fronds of the mangrove palm Nypa fruticans from Brunei [154,155,156], during their investigations concerning tropical marine mangrove fungi on three particular regions, i.e., the Indian Ocean (Seychelles), the Straits of Malacca (North Sumatra, Indonesia), and the South China Sea (Brunei) [157,158,159]. These early studies were perhaps the driving force and what set up the research on palm fungi in the 1990s. After those first three taxonomical novelties, numerous further reports of new taxa were made on fronds of intertidal N. fruticans, predominantly from Brunei, e.g., refs. [105,110,117,131,160,161,162,163,164,165] and Malaysia, e.g., refs. [116,143,166,167], which yielded more than thirty new species and seven new genera (Table 1).
Nipa palm was found to be a “very distinct mangrove habitat” for fungal surveys, where a “largely distinct mycota” was found, including several “unique marine species belonging to genera consistently found on terrestrial palms” [168]. Hyde [110] observed that Astrosphaeriella, Linocarpon, and Oxydothis, which are genera typically associated with terrestrial palm petioles in the tropics, were often associated with decaying intertidal Nipa palms. Not only was this mangrove palm tree important for systematic and descriptive taxonomy studies, but it was also a very important substratum for some of the first studies on the ecology of palm fungi, e.g., ref. [7], as will be discussed later. Although most studies focused on the intertidal fungi occurring on N. fruticans, studies have also been conducted in order to access the fungi inhabiting the aerial parts of this mangrove palm [165].
Perhaps encouraged by those early records, during the last decade of the 20th century, Hyde et al. carried out an extensive survey of palmicolous fungi from a wide range of tropical palm species in different regions of the world (Figure 1). The tropical regions of Southeast Asia and Australasia comprised the countries in which most of the collections were made, such as Brunei [113,115,116,117,133,134,138,154,155,160,161,162,163,164,169,170,171], Indonesia [109,113,115,124,126,131,133,134,172], Malaysia [110,116,117,118,123,131,133,134,165,167,172], and Australia [108,109,112,114,117,133,135,138,172]. Other regions and countries were moderately or occasionally sampled, including Thailand [162], Papua New Guinea [117,123,127,131,133,173,174,175], Philippines [117,125,167], Japan [117,133], Ecuador [128,138,171], Brazil [107], and the USA [122,124]. All the regions surveyed revealed the presence of an enormous diversity of fungi, among which, 22 genera and more than 80 species were described as new to science, several existing genera were re-examined, and some new combinations were proposed. In fact, one of the most remarkable outcomes of these early works by Hyde was the number of new genera introduced (Table 1). Not only did they reveal the great untapped diversity of the fungal communities that inhabit palm trees in the tropics, but they also discovered a very particular unknown and underexplored taxonomic group of ascomycetes which make up what has been termed palm fungi (Table 2). Therefore, this comprehensive research began to unveil and shape the community of palmicolous fungi, insomuch that it allowed to build up and consolidate the knowledge on the common mycota that typically inhabit the tissues of tropical and subtropical palms. According to Hyde [111], fallen palm rachides and leaves in the tropics were found to be invariably colonised by fungi of the genera Astrosphaeriella, Linocarpon, Oxydothis, and Phomatospora. It later became clear that, in addition to Oxydothis, several other genera of Xylariales had common representatives on palm trees, including Anthostomella, Apioclypea, Arecomyces, Astrocystis, Capsulospora, Fasciatispora, Nipicola, and Pemphidium [134,138] (Table 2).
Along with Hyde’s work on clarifying, redescribing, illustrating, and monographing existing genera [105,107,111,113,117,124,126,127,130,133,171], these studies resolved certain taxonomic relationships within various important fungal families and orders. Moreover, they also expanded the knowledge of tropical mycology, especially with regard to the biodiversity of fungi that inhabit tropical hotspots, such as rainforests. Striking examples include the description of numerous new species in the genera Linocarpon [105,154,165,172], Oxydothis [111,112,117,156], and Anthostomella [133,175], three of the most common genera found on palms (Table 2); the clarification of certain unclear generic concepts for some of the genera, and their corresponding species, found on palms, including Pemphidium [107,111,135], Guignardia [126], and Roussoella [171,176]; the description and discussion of some amphisphaeriaceous fungi occurring on palms, such as the genera Fasciatispora [124,161], Myelosperma [113], Seynesia [127], Arecophila [131], and Amphisphaeria [136]; and the investigation of palmicolous fungi inhabiting the palms in tropical Australian, e.g., refs. [112,114], Bruneian, e.g., refs. [170,174], and Ecuadorian, e.g., refs. [128,171] rainforests. These first case studies in the early 1990s verified the existence of a well-represented set of morphological characters to describe these genera, as well as to distinguish them from other related genera, and to clarify the existence of new morphologically similar genera. As a result, it began to become clearer what the taxonomic placement of palm fungi was in higher taxonomic ranks, including different families of the Xylariales and other less representative orders, e.g., ref. [138] (Table 2).
The large number of new genera and species reported on palms by Hyde in the early 1990s quickly prompted a growing interest in the mycobiota of these hosts and several of his students and collaborators, such as Fröhlich, Taylor, Aptroot, and Goh, worked to expand the knowledge of palmicolous fungi. Thus, in addition to the many new taxa recorded, different aspects of these fungi have been studied and have contributed to the fundamental knowledge of fungi and their biodiversity. This intensive research ultimately led to the description of at least three new families to accommodate genera that have been described and found to be common on palms, along with extant related genera, namely Phaeochoraceae for Cocoicola, Phaeochora and Serenomyces [177], Apiosporaceae for Apiospora and Appendicospora [143], and Myelospermaceae for Myelosperma [178].
Hyde, Fröhlich, Taylor, Aptroot, and Goh, studying ascomycetes developing on living, diseased, and dead palm material, surveyed different regions from East, South, and Southeast Asia, including, respectively, China (Hong Kong, Hubei, Hunan) [6,8,129,140,142,143,144,146,147,148,149,151,179,180,181,182,183,184,185,186], India [152], and Brunei, Indonesia, Laos, Malaysia, Philippines, and Singapore [6,8,122,139,140,143,147,148,149,150,153,179,181,185,187,188,189,190,191,192,193]. The Australasian regions were also intensively surveyed, including Australia [6,8,121,140,141,143,144,147,148,181,185,187,189,194,195,196,197,198,199,200,201,202,203,204] and Papua New Guinea [150,198]. Other regions and countries were also frequently or occasionally surveyed, including Ecuador (South America) [6,119,143,144,150,179,181,185,205,206,207,208], USA [120], South Africa [208,209,210], Seychelles [8,178,181], and European countries, such as Switzerland and Great Britain [8] (Figure 1). As expected, the huge diversity of fungi collected from palm trees increased, with 22 new genera and more than 200 new species described. As a result, the importance of palm fungi began to become more evident as studies continuously revealed their broad taxonomic structure (Table 2). Most of these studies and their outcomes have been compiled in the first book published by Hyde and co-workers in the Fungal Diversity Research Series concerning palmicolous fungi, which was entitled “Genera of ascomycetes from palms” and treated 100 selected genera of common fungi inhabiting the tissues of tropical palms [104]. Moreover, another book on the same series, entitled “Palm microfungi”, has been published in the same year, which supplemented the previous information describing the ascomycetes found on palms during their comprehensive collections in Australia, Brunei, and Hong Kong [6]. Remarkably, in addition to the ongoing revision of several genera of ascomycetes with common representatives on tropical palms, Fröhlich and Hyde [6] described 65 taxa as new to science, including 3 new genera and an impressive number of 23 new species of Oxydothis, considered to be the genus most commonly found on palms and invariably one of the earliest colonisers of dead palm leaves and fronds [185,211] (Table 1 and Table 2).
Following Hyde’s early studies of palm fungi in tropical rainforests, e.g., refs. [112,170,171], one intensively studied region worth mentioning is the rainforests of North Queensland (Australia) (Figure 1), where an immense diversity of fungi has been found in collections of different endemic palm species, such as Archontophoenix alexandrae, Laccospadix australasica, Licuala ramsayi, Linospadix microcaryus, L. monostachyos, and Oraniopsis appendiculata, e.g., refs. [121,140,141,143,145,147,148,181,185,187,189,194,195,196,197,198,199,200,202]. While most of the studies conducted by Hyde and co-workers were dedicated to surveying saprobic fungi that develop on palm trees, some of these studies in the North Queensland rainforest were conducted to survey palm phytopathogens, particularly those parasitic on palm leaves and causing leaf spot diseases. Palm phytopathogens were also documented in studies conducted during an expedition of the British Mycological Society to Ecuador in August 1993 to gather fungi developing on palms in the rainforest of Cuyabeno [119,143,144,150,181,185,206,207,208]. Moreover, Hyde and Cannon [212] monographed members of the Catabotrydaceae, Phaeochoraceae, and Phyllachoraceae families that occur in association with tar spots on palms. In addition to treating these families and their placement in the fungal classification, several genera were treated and analysed, some were re-introduced based on palm collections, and four genera and three species were described as new to science (Table 1). Some of the genera discussed by Hyde and Cannon [212] represent important taxa in the phytopathogenic mycobiota of palm trees, such as the genus Serenomyces, a group of mostly biotrophic fungi apparently known only in association with members of Arecaceae [213,214].
Although some palm phytopathogens have been identified, most studies on palmicolous fungi in the last decade of the 20th century focused on the myriad of saprobic fungi that inhabit the palms from tropical rainforests, where palm litter is a major component. Samples were collected from many different palm trees that inhabit the tropics, including from typical Phytelaphas spp. of Central and South America to typical Licuala spp. of Southern China (Figure 1). Palm material, such as senescent and dead petioles, rachides, stems, fronds, and decaying and dead trunks, yielded plentiful novelties (Table 1). One interesting palm tree worth mentioning is A. alexandrae, an endemic palm from Queensland, Australia. Studies on the palmicolous fungi that inhabit the tissues of this palm have perhaps been the driving force for studying other questions about the taxonomy and ecology of palm fungi, as well as extending the research to fungi associated with palms from non-tropical habitats. In fact, its endemic nature and relative geographic isolation make A. alexandrae an ideal substratum for studying host-specificity and fungal biogeography, which have been identified as important factors for the description of many novel palmicolous fungi when hosts are studied in their natural environment [142,194]. Likewise, the description of the new palmicolous genus Cannonia [204] collected from Trachycarpus fortunei in Australia, outside its native temperate range in parts of China and some neighbouring countries (Table 1), also raised questions about the importance of studying the biogeography of palm fungi. As a result, studies on palmicolous fungi that expanded their geographical boundaries and temperate palms also began to be surveyed [215].
The constant description and illustration of new taxa found on palm trees has often led to discussions of their placement in the fungal classification. As a result, several families, and their respective genera, with common representatives on palms, have been extensively treated and the taxonomy of palmicolous fungi, at least in the tropics, has become clearer, e.g., refs. [6,8,104,143,150,171,178,181,185,212,216,217] (Table 2). Noteworthy examples include the description and discussion of common genera that occur on palms with the introduction of several new species, such as Nipicola [137], Nectria, and allied genera [207], Massarina [139,149], Anthostomella [143,209,218,219,220,221], Astrosphaeriella [144,148], and Neolinocarpon [140], whose geographical distribution has widened considerably throughout Australasia, Southeast Asia, South America (Ecuador), and China (including Hong Kong). Other common genera discussed, with the introduction of new species, include Dictyosporium [184] and Lasiosphaeria and similar genera, such as Chaetosphaeria, Iodosphaeria, and the newly described genus Arecacicola [146,181,185] (Table 1). Likewise, some doubtful species described for some genera that occur on palms have been clarified and revised, namely in Mycosphaerella and Sphaerella, which are common phytopathogens [145], Didymosphaeria and similar taxa, which include truly terrestrial ascomycetes with ascospores with appendages described from palms [147,222,223], and genera of unitunicate ascomycetes with apiospores, which are frequently recorded on palms, such as Anthostomella, Apioclypea, Apiospora, Appendicospora, and the new described genera Brunneiapiospora and Palmaria (as Palmomyces) [143] (Table 1). Several xylariaceous genera recorded on palms have also been treated based on herbarium specimens and fresh material collected during the investigation on palmicolous fungi, which led to the description of several new species, for example, of the genus Astrocystis [150].
Following the incredible contribution to the knowledge of fungal biodiversity and their taxonomy, these studies have also made it possible, on several occasions, to discuss and even uncover some links between sexual and asexual morphs [150,167,181,203,208,224]. The importance of studying “anamorph-teleomorph” connections was strongly emphasised in the last compilation book published by Taylor and Hyde [8] in the Fungal Diversity Research Series on palm fungi, entitled “Microfungi of tropical and temperate palms”. As well as continuing the previous studies on palm fungi carried out on tropical rainforest palms, Taylor and Hyde [8] also studied fungi associated with palms in non-rainforest habitats in the tropics and palms in temperate habitats. Interestingly, this is the first book to deal not only with the identification of palmicolous fungi but also with the extent of their diversity and the factors that affect it, which reflects the complexity of the investigation carried out by Hyde and co-workers and the diversity of the approaches that were used. A total of thirty-four new species, including four new species of Anthostomella, and one new genus, Tribulatia, were described (Table 1), which is a surprising number considering the extent of the sampling and the number of taxa already described from palms by Hyde and co-workers.
The studies on palmicolous fungi in tropical regions continued, with other regions beginning to be surveyed more systematically, including many reports from additional Hyde co-workers, namely McKenzie, Pinnoi, Pinruan, and Yanna. Although the description of new taxa remained the main objective of these studies, which yielded more than thirty species and seven genera described as new to science (Table 1), they have also begun to provide ecological data on the communities of tropical palmicolous fungi that inhabit some tropical palm tree species, such as Livistona chinensis and Phoenix hanceana, e.g., refs. [13,14,15]. Yanna et al. [224,225,226,227,228,229] described several new species from different palm tree species in Hong Kong, comprising the typical Ascomycota assemblage commonly found on palms, particularly species of Appendicospora [225], to atypical hyphomycetes and coelomycetes genera, such as Koorchaloma [226], Staurophoma [224], Endomelanconium [227], and Everhartia [229].
Although the initial studies on palms focused mainly on the evaluation of the Ascomycota coverage through the presence of its sexual morphs on the host, the evident potential of these hosts for biodiversity surveys has led to a diversification of approaches and, consequently, discoveries. As a result, a considerable number of reports began to describe several new species and genera of palmicolous hyphomycetes, particularly dematiaceous hyphomycetes, in addition to the usual sexual morphs of ascomycetes from well-studied regions, such as Australia (North Queensland) [15,189,201,202,230], China (Hong Kong and Hainan) [182,183,184,231,232,233,234,235], Brunei, and Thailand [166,188,189,190,228,236,237,238] (Figure 1). In addition to hyphomycetes, species of the discomycetes genus Lachnum, new species of which are often found on palm trees [239,240], have also been reported in tropical China, e.g., ref. [231].
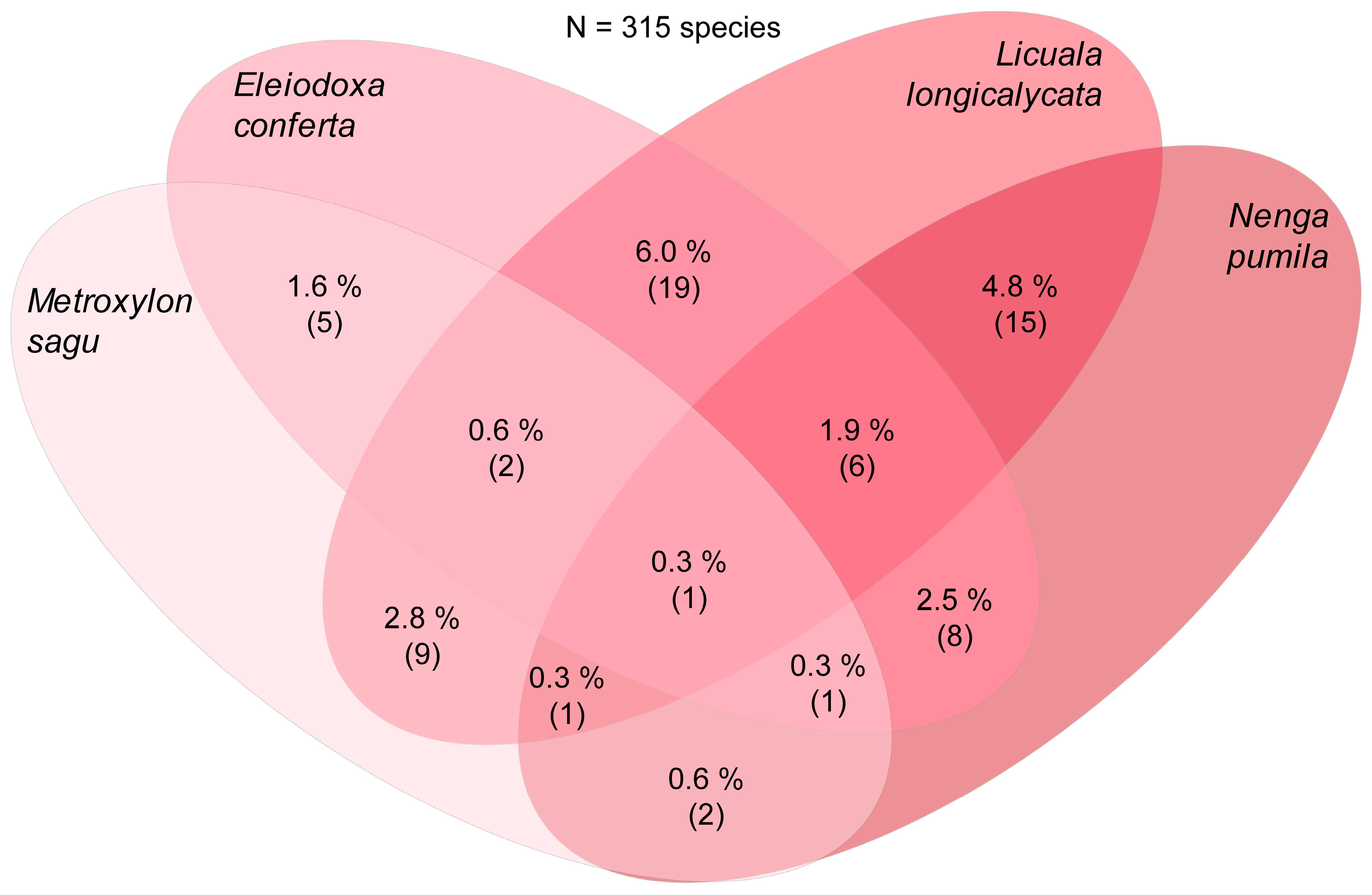
Just before the input of molecular data began to broaden and strengthen the research of Hyde and co-workers on palm fungi, McKenzie, Pinnoi, and Pinruan et al., studying the fungal diversity on palms from the Sirindhom Peat Swamp Forest at Narathiwat, Southern Thailand, found and described several new palmicolous taxa [236,241,242,243,244,245,246,247]. Thus, in addition to new species of the typical Ascomycota assemblage reported from palm tissues in the tropics, including new species of Submersisphaeria [247], Jahnula [242], and the new genus Unisetosphaeria [245] (Table 1), some new palmicolous “anamorphs” were introduced, including new species of Chalara [241], Dactylaria [245], Custingophora, Vanakripa [246], Craspedodidymum [243], and Stachybotrys [244]. Moreover, following the studies carried out by Yanna et al., ecological data on tropical palm fungal communities have also been documented in the peat swamp palms Eleiodoxa conferta [248] and Licuala longicalycata [249], as well as in Calamus spp. [16].
2.1.3. The Palmicolous Hyphomycetes from Central American Countries
A perusal of the available literature on palm fungi reveals that, to date, no intensive studies have been carried on palmicolous “anamorphs”, i.e., hyphomycetes and especially coelomycetes. However, it is worth mentioning a few scattered studies that reported new species and genera of palmicolous hyphomycetes. These studies have shown that the predominant group of “anamorphs” in palm litter are the dematiaceous hyphomycetes. In fact, one of the first consistent reports of palmicolous hyphomycetes is that of Ellis in the 1960s and 1970s, who reported several new species and some new combinations based on collections of palms from West Africa, Southeast Asia, and Central America in his series of papers and books about “Dematiaceous hyphomycetes” [29,30,31,32,250,251,252,253,254,255,256,257,258].

{kind=link}
{kind=link}
Table 1.
Genera described as new to science and found on Arecaceae hosts in the last three decades.
| Genus | Type Species | Host | Country/Region | Sequence Data 1 | Reference |
|---|---|---|---|---|---|
| Acarocybellina | A. arengae | On a dead leaf of Arenga engleri | Japan | N/A | [259] |
| Acarocybiopsis | A. cubitaensis | On a dead trunk of Roystonea regia | Cuba | N/A | [260] |
| Acuminatispora | A. palmarum | On decaying petioles and rachides of an unidentified palm in mangrove | Thailand | A | [261] |
| Agrabeeja | A. kavakapriya | On synnemata of Melanographium citri on a rachis of Korthalsia grandis | Singapore | N/A | [262] |
| Allodiatrype | A. arengae | On a dead petiole of Arenga pinnata | Thailand | A | [263] |
| Anabahusakala | A. amapensis | On decaying leaves of Syagrus sp. | Brazil (Amapá) | N/A | [264] |
| Anisospadicoides | A. macrocontinua (as Spadicoides macrocontinua) | On a rotten petiole of an unidentified palm | Peru | N/A | [64,265] |
| Apioclypea | A. livistonae | On a rachis of Livistona sp. | Papua New Guinea | N/A | [175] |
| Apogaeumannomyces | A. perplexus | On a decaying frond of an unidentified palm | Peru | N/A | [67] |
| Appendicospora | A. coryphae | On dead rachides of Corypha elata | Philippines | N/A | [125] |
| Appendispora | A. frondicola | On a dead rachis of Oncosperma horridum on forest floor | Brunei | N/A | [115] |
| Arecacicola | A. calami | On a trunk of Calamus sp. | Indonesia (Java) | N/A | [185] |
| Arecomyces | A. frondicola | On a rachis of Arenga undulatifolia | Brunei | N/A | [138] |
| Arecophila | A. gulubiicola | On a dead trunk of Gulubia costata | Papua New Guinea | N/A | [131] |
| Ashtaangam | A. Sundaram | On a rachis of an unidentified palm | Malaysia | N/A | [266] |
| Astrosphaeriellopsis | A. bakeriana | On a petiole of Borassus sp. | Thailand | A | [267] |
| Asymmetricospora | A. calamicola | On a dead stem of Calamus caryotoides | Australia (Queensland) | N/A | [141] |
| Atrosetaphiale | A. flagelliformis | On a decayed petiole of an unidentified palm | Peru | N/A | [65] |
| Aunstrupia | A. nodipes | On rotten and dead leaves and rotten petiole and branches of unidentified palms | China (Guangdong) | A | [268] |
| Bacusphaeria | B. nypae | On a petiole base of Nypa fruticans | Malaysia | A | [269] |
| Baipadisphaeria | B. spathulospora | On a trunk of Licuala longicalycata submerged in peat bog | Thailand | A | [270] |
| Basauxia | B. pulchra | On a rachis of an unidentified palm | Malaysia | N/A | [266] |
| Bhadradriella | B. hyalina | On fallen pods of Roystonea regia | India (Andhra Pradesh) | N/A | [271] |
| Brachysporiopsis | B. chinensis | On a decaying rachis of Livistona chinensis | China (Hong Kong) | N/A | [228] |
| Brobdingnagia | B. nigeriensis | On tissues of Calamus sp. | Nigeria | N/A | [212] |
| Brunneiapiospora | B. javensis | On a rachis of Calamus sp. | Indonesia (Java) | N/A | [143] |
| Bulbocatenospora | B. complanata | On fallen leaves of Bactris setulosa | Venezuela | N/A | [272] |
| Cannonia | C. australlis | On rotten branches of Butia yatay | Argentina | N/A | [204] |
| Capsulospora | C. frondicola | On a rachis of Daemonorops sp. | Brunei | N/A | [134] |
| Carinispora | C. nypae | On decaying intertidal fronds of Nypa fruticans | Brunei | N/A | [162] |
| Castanedospora | C. pachyanthicola | On the petiole of a dead leaf of Sabal palmetto | USA (Florida) | A | [273] |
| Caudatispora | C. palmicola | On a dead rachis of Phytelaphas | Ecuador | N/A | [119] |
| Cenangiumella | C. rattanicola | On a dead rattan sheath of Calamus conirostris | Brunei | N/A | [6] |
| Chitinasiproducens | C. palmae | ||||
| Circinoconiopsis | C. amazonica | On decaying leaves of Oenocarpus sp. | Brazil (Pará) | N/A | [274] |
| Cocoicola | C. cylindrospora | On petioles of Cocos nucifera | Papua New Guinea | N/A | [123] |
| Corynesporasca * | C. caryotae | On rotting leaves of Caryota urens | Sri Lanka | N/A | [275] |
| Curvatispora | C. singaporensis | On a fallen decaying frond of Livistona spinosa | Singapore | N/A | [153] |
| Cyanopulvis | C. australiensis | On a dead rattan of Calamus australis | Australia (Queensland) | N/A | [6] |
| Cylindrotorula | C. indica | On a decaying spathe of Cocos nucifera | India (Maharashtra) | A | [276] |
| Diabolocovidia | D. claustri | On leaves of Serenoa repens | USA (Florida) | A | [277] |
| Dictyopalmispora | D. palmae | On decaying leaves of Licuala longicalycata | Thailand | A | [278] |
| Discopycnothyrium | D. palmae | On the branches of an unidentified palm | Thailand | A | [279] |
| Durispora | D. elaeidicola | On dead rachides of Elaeis guineensis | Malaysia | N/A | [118] |
| Dwibahubeeja | D. indica | On leaves of C. tenuis | India (Uttar Pradesh) | N/A | [280] |
| Endosporoideus | E. pedicellatus (as E. pedicellata) | On a dead petiole of Phoenix hanceana | China (Hong Kong) | N/A | [235] |
| Fasciatispora | F. nypae | On a rotten frond of intertidal Nypa fruticans | Brunei | A | [161] |
| Fissuroma | F. maculans | On dead leaves of Arenga westerhoutii | Thailand | A | [281] |
| Flammispora | F. bioteca | On dead leaves of Licuala longicalycata submerged in peat swamp | Thailand | A | [282] |
| Fluviatispora | F. tunicata | On submerged rachides of Livistona sp. | Papua New Guinea | N/A | [174] |
| Frondicola | F. tunitricuspis | On decaying fronds of Nypa fruticans | Brunei | N/A | [162] |
| Frondisphaeria | F. palmicola | On a rachis of Eugeissona minor | Brunei | N/A | [170] |
| Frondispora | F. bicalcarata | On dead petioles of Chamaerops humilis | Italy | N/A | [111] |
| Gossypinidium | G. sporodochiale | On a dead rachis of Praestoea montana | Puerto Rico | A | [283] |
| Guestia | G. gonetropospora | On a dead rachis of Mauritia flexuosa | Ecuador | N/A | [150] |
| Haploanthostomella | H. elaeidis | On dead leaves and rachis of Elaeis guineensis | Thailand | A | [284] |
| Haplohelminthosporium | H. calami | On living leaves and petioles of Calamus sp. | Thailand | A | [285] |
| Helensiella (as Digitella) | H. rigidophora (as D. rigidophora) | On a rachis of an unidentified palm | Mexico (Veracruz) | N/A | [286,287] |
| Helminthosporiella | H. stilbacea | On a dead petiole of Cocos nucifera | Thailand | A | [285,288] |
| Hemisynnema # | H. malayasianum | On a rachis of an unidentified palm | Malaysia | N/A | [289] |
| Hyalobelemnospora | H. amazonica | On a rotten petiole of an unidentified palm | Peru | N/A | [64] |
| Kalamarospora | K. multiflagellata | On rachides of dead leaves of Sabal palmetto | USA (Florida) | N/A | [290] |
| Letendraeopsis | L. palmarum | On leaves of Euterpe oleracea | Brazil (Pará) | N/A | [291] |
| Lockerbia | L. palmicola | On dead rachides of an unidentified palm | Australia (Queensland) | N/A | [114] |
| Longicorpus | L. striatisporus (as L. striataspora) | On a decayed rachis of Nypa fruticans | Thailand | A | [9] |
| Mackenziella (as Mackenziea) | M. livistonae | On decaying rachides of Oraniopsis appendiculata | Australia (Queensland) | N/A | [15] |
| Maculatifrondes (as Maculatifrondis) | M. aequatoriensis | On leaves of an unidentified palm in rainforest | Ecuador | N/A | [208] |
| Maculatipalma | M. frondicola | On a leaf of Linospadix microcarya | Australia (Queensland) | N/A | [197] |
| Malthomyces | M. calamigena (as M. calamigenus) | On tissues of Calamus rudentum | Sri Lanka | N/A | [212] |
| Manokwaria | M. notabilis | On dead rachides of an unidentified palm in freshwater swamp | Indonesia | N/A | [109] |
| Monosporoschisma | M. elegans | On a dead material of an unidentifed palm | Chian (Hainan) | A | [268] |
| Neoastrosphaeriella | N. krabiensis | On a petiole of Metroxylon sagu | Thailand | A | [281] |
| Neobarrmaelia | N. hyphaenes | On leaves of Hyphaene sp. | South Africa | A | [292] |
| Neolinocarpon | N. globosicarpum | On decaying intertidal fronds of Nypa fruticans | Brunei | N/A | [162] |
| Neoxylaria | N. arengae | On a dead petiole of Arenga pinnata | Thailand | A | [293] |
| Nigromammilla (as Nigramammilla) | N. calami | On a sheath of dead rattan of Daemonorops margaritae | China (Hong Kong) | N/A | [179] |
| Nipicola | N. carbospora | On immersed fronds of Nypa fruticans | Brunei | N/A | [163] |
| Nusia | N. scheeleae | On a rachis of Scheelea insignis | Singapore | N/A | [294] |
| Nypaella | N. frondicola | On intertidal fronds of Nypa fruticans | Brunei | N/A | [164] |
| Ornatispora # | O. palmicola | On a dead rachis of an unidentified palm | Ecuador | N/A | [181] |
| Oxodeora | O. petrakii | On living fronds of Oreodoxa regia | Dominican Republic | N/A | [212] |
| Palmaria (as Palmomyces) | P. montanea (as P. montaneus) | On a leaf of Oraniopsis appendiculata | Australia (Queensland) | N/A | [143] |
| Palmeiromyces | P. chamaeropicola | On leaf spots of Chamaerops humilis | Portugal | A | [295] |
| Palmicola | P. archontophoenicis | On a fallen rachis of Archontophoenix alexandrae | Australia (Queensland) | N/A | [108] |
| Paracapsulospora | P. metroxyli | On a dead Metroxylon sagu | Thailand | A | [296] |
| Paradactylella | P. peruviana | On a rotten petiole of an unidentified palm | Peru | N/A | [64] |
| Paraproliferophorum | P. hyphaenes | On living leaves of Hyphaene sp. | South Africa | A | [297] |
| Pararamichloridium | P. livistonae | On leaves of Livistona australis | Australia (New South Wales) | A | [298] |
| Parateichospora | P. phoenicicola | On leaves of Phoenix reclinata | South Africa | A | [299] |
| Phaeochoropsis | P. neowashingtoniae | On leaves of Neowashingtonia filamentosa | USA (California) | N/A | [212] |
| Phaeomonilia | P. pleiomorpha | On a decaying petiole of an unidentified palm submerged in stream | Mexico (Veracruz) | N/A | [300] |
| Phruensis | P. brunneispora | On a dead trunk of Licuala longicalycata | Thailand | A | [301] |
| Polybulbophiale | P. palmicola | On the decaying petiole of Licuala sp. | Brunei | N/A | [190] |
| Porodiplodia | P. livistonae | On leaves of Livistona australis | Australia (New South Wales) | A | [302] |
| Pseudopalawania | P. siamensis | On a dead rachis of Caryota sp. | Thailand | A | [303] |
| Pulmosphaeria | P. archontophoenicis | On a dead petiole of Archontophoenix alexandrae | Australia (Queensland) | N/A | [194] |
| Quasiphoma | Q. hyphaenes | On leaves of Hyphaene sp. | South Africa | A | [292] |
| Rachidicola | R. palmae | On a rachis of Calamus sp. | China (Hong Kong) | N/A | [129] |
| Rattania | R. setulifera | On leaves of Calamus thwaitesii | India (Goa) | N/A | [304] |
| Rogergoosiella | R. roystoneicola | On a dead petiole of Roystonea regia | Cuba | N/A | [305] |
| Sabalicola | S. sabalensioides | On petioles of Sabal serrulata | USA (Florida) | N/A | [122] |
| Sawantomyces | S. indicus (as S. indica) | On a spathe of Cocos nucifera | India (Maharashtra) | N/A | [306] |
| Setophiale | S. unisetulata | On a decayed petiole of an unidentified palm | Peru | N/A | [65] |
| Sorokinella | S. appendicospora | On a dead petiole of Livistona chinensis | China (Hong Kong) | N/A | [6] |
| Stratiphoromyces | S. brunneisporus | On decaying petioles of Licuala sp. | Brunei | N/A | [189] |
| Striatiguttula | S. nypae | On a decayed rachis of Nypa fruticans | Thailand | A | [9] |
| Thailandiomyces | T. bisetulosus | On senescent trunks of Licuala longicalycata | Thailand | A | [307] |
| Tirisporella | T. beccariana | On decaying leaf bases of Nypa fruticans | Malaysia | N/A | [167] |
| Tretendophragmia | T. palmivora | On a rachis of Korthalsia sp. | Singapore | N/A | [308] |
| Tretocephala | T. decidua | On a leaf sheath and rachis of Oncosperma horridum | Singapore | N/A | [309] |
| Tribulatia | T. appendicospora | On a dead petiole of Archontophoenix alexandrae | Australia (Queensland) | N/A | [8] |
| Triseptatospora | T. calami | On dead petioles of Calamus sp. | Thailand | A | [310] |
| Unisetosphaeria | U. penguinoides | On a petiole of Eleiodoxa conferta submerged in peat swamp | Thailand | N/A | [245] |
| Uwemyces | U. elaeidis | On leaves of Elaeis oleifera | Colombia | A | [288] |
| Venustocephala | V. aequatorialis | On a decayed petiole of an unidentified palm | Ecuador | N/A | [65] |
| Venustisporium (as Venustusporium) | V. chelyoforme (as V. chelysforme) | On fallen rotten leaves of Bactris setulosa | Venezuela | N/A | [311] |
| Veramycella | V. bispora | On rachides of dead leaves of Sabal palmetto | USA (Florida) | N/A | [312] |
| Veramyces | V. manuensis | On a rotten petiole of an unidentified palm | Peru | N/A | [64] |
| Waihonghopes | W. australiensis | On a decaying rachis of Oraniopsis appendiculata | Australia (Queensland) | N/A | [15] |
1 Availability of DNA sequence data for the type species of each genus, N/A: DNA sequence data not available; A: DNA sequence data available. * The monotypic genus Corynesporasca was introduced to accommodate C. caryotae and linked with an unnamed Corynespora asexual morph in culture by Sivanesan [275]. Corynesporasca has been treated as a synonym of Corynespora in several studies, e.g., ref. [313]. However, the present review follows Hyde et al. [314], who did not synonymize Corynesporasca under Corynespora. Corynespora was shown to be polyphyletic and Corynespora-like asexual morphs have been associated with many genera, e.g., ref. [315]. Therefore, the type species of both genera may be unrelated. # Genera currently synonymised under other genera. Hemisynnema was synonymised under Morrisiella by Wu and Zhuang [316]. Ornatispora was synonymised under Stachybotrys by Wang et al. [317].
There are several scattered reports of palmicolous hyphomycetes in different regions of the world. For instance, species of helicosporous fungi from various genera, such as Drepanospora, Helicoma, Helicomyces, Helicosporium, and Xenosporium, have been recorded in collections of palm tissues. These were reviewed by Goos [318,319,320,321,322,323,324,325] and Goos et al. [326] during their studies on anamorphic genera of helicosporous fungi. In the 1990s, along with the examples previously cited by Hyde and co-workers, several palmicolous hyphomycetes were reported in studies carried out by Subramanian in India and in Southeast Asian countries, such as Malaysia and Singapore (Figure 1). In these studies, Subramanian [259,262,266,289,294,308,309,327,328] introduced many new species and genera of dematiaceous hyphomycetes from different palm trees species, including Tretendophragmia [308], Tretocephala [309], Ashtaangam, Basauxia [266], Acarocybellina [259], Agrabeeja [262], Nusia [294], and Hemisynnema [289] (Table 1). These records further emphasised the importance of palm trees for the description of taxonomic novelties and for the study of fungal biodiversity, and the same trend would be found in the palm trees of Central American countries.
Palmicolous hyphomycetes have been widely collected from palms in Central American countries, including Cuba and Mexico (Figure 1), by Castañeda-Ruiz, Holubová-Jechová, Mena-Portales, Mercado-Sierra, and many other co-workers, following their previously mentioned investigation of hyphomycetes from Cuba in the 1980s. Although most of these studies were not conducted to explicitly evaluate palm fungi, the extent of these reports has made it possible to reveal the composition of the assemblage of palm hyphomycetes that inhabit palm tissues in the tropics. Furthermore, it has become evident, particularly in studies from Cuba, that some parts of palm trees, mainly their decaying rachides and large petioles, are exceptional substrata for the growth and development of microfungi, mainly hyphomycetes, and several new taxa have been identified [260,305,329,330,331,332,333,334,335,336,337,338,339,340,341,342,343,344].
Similar to what Hyde and co-workers discovered in their investigation in East and Southeast Asia, Australasia, and Ecuador, many of the fungi growing on palm trees in Cuba were described as new to science and found to form an autochthonous mycobiota, where host-specificity, sometimes at the host genus or species level, is often observed. The fungus–host plant relationship and the factors that affect it were expressly discussed and reviewed by Mercado-Sierra et al. [329], particularly for genera of palm trees that grow abundantly in Cuba, namely Roystonea, Cocos, and Coccothrinax. The Cuban royal palm Roystonea regia appears to be a particular case where an enormous diversity of fungi was found, including 265 species from different taxonomic groups, a number much higher than that reported for other plant species endemic to Cuba [345]. Moreover, its relevance and importance in studies of hyphomycetes were pointed out by Mercado-Sierra [73]. Many of the fungi identified on palm trees from Cuba were new reports for Cuban mycobiota and several taxa were described as new to science, including more than thirty species and six genera, viz. Consetiella [75], Holubovaea [73], Phragmospathulella [96], Cheiromyceopsis [99], Rogergoosiella [305], and Acarocybiopsis [260] (Table 1).
Several Cuban provinces and localities, such as Pinar Del Río, Camagüey, Matazanas, and Sancti Spíritus, have been extensively sampled during surveys of hyphomycetes in protected natural areas in Cuba, and on several occasions these hyphomycetes have been collected from palm trees, e.g., refs. [338,346,347,348,349,350,351]. Although the extension of these studies made it possible to uncover the assemblage of palm hyphomycetes in those locations, their objective was mainly to study ecological stations, biosphere reserves, and protected areas in order to promote the conservation of important hotspots of fungal diversity. These studies continue to be carried out today and report an exceptional diversity of palm fungi. Recently, Mena-Portales et al. [352], following Mercado-Sierra et al. [329], reviewed and analysed the relationship between fungal diversity and palms trees in Cuba by compiling information on some interesting species of hyphomycetes found in different Arecaceae hosts.
Similar studies have also been carried out in other Central American countries (Figure 1). Mercado-Sierra et al. [353,354] reported some palmicolous hyphomycetes from Costa Rica. Recently, a checklist of asexual fungi from Costa Rica, which compiled information obtained during 1927 to 2018 based on scientific papers, was presented by Granados-Montero et al. [355], including several reports of hyphomycetes and coelomycetes on palm trees. Very few palm fungi, including hyphomycetes, have been reported from Puerto Rico and most reports have been summarised in an annotated bibliography entitled “The Fungi of Puerto Rico and the American Virgin Islands” by Stevenson [356]. However, some new species were later introduced based on occasional palm collections, e.g., refs. [240,357,358]. Palmicolous hyphomycetes from Panama and Nicaragua have been listed in checklists of Panamanian and Nicaraguan fungi by Piepenbring [359] and Delgado-Rodríguez [360], respectively. Likewise, several palmicolous hyphomycetes were reported in Mexico during studies carried out in order to increase the knowledge about Mexican hyphomycetes [286,300,361,362,363,364,365,366,367,368,369,370,371], where two new genera, Phaeomonilia [300] and Digitella [286], and some new species were introduced (Table 1). Similar to the studies in Cuba, the investigation in Mexico, which continues today, was not pursued to study palm fungi, but to inventory and gain knowledge about conidial fungi from plant litter in tropical forests, particularly in the states of Campeche, Tabasco, and Veracruz, due to the accelerated deforestation of tropical habitats, e.g., refs. [367,371,372,373,374,375,376,377,378,379,380]. As a result, since palm trees are an important component of the flora of these forests, several palmicolous hyphomycetes have been reported.
In addition to the studies in Central American countries, South American countries have also been occasionally surveyed (Figure 1). Therefore, palmicolous hyphomycetes have been reported in collections from Argentina [381], Colombia [382], Peru [265], and Venezuela [272,311,383,384,385,386,387], where some new species and genera have been introduced (Table 1).
2.1.4. Palm Fungi from Understudied Tropical Hotspots, Argentina, India, and Brazil
Some scattered studies have surveyed palmicolous fungi in Argentina. However, with the exception of Mercado-Sierra et al. [381], these were not systematic studies of descriptive taxonomy, but studies carried out to better understand the diversity of ascomycetes in woody parts of palms in Argentina, especially in areas or parks that had been proposed as natural reserves for protection and where some native palm species are an important element of the local flora, including Butia yatay, Euterpe edulis, and Syagrus romanzoffiana [388,389,390]. Capdet and Romero [389] summarised previous information on palm fungi and their occurrence in Argentina, reflecting on the lack of knowledge about palm fungi in the country.
Similarly, considering the available literature on palm fungi, except for a few stray collections, no comprehensive investigation on the fungal diversity that occurs on palms in India has been carried out (Figure 1). However, some remarkable reports by Subramanian and his students from Chennai, P. R. Rao, D. Rao, and V. Rao, and colleagues from Hyderabad and Bhat and colleagues from Goa, revealed a considerable diversity of palmicolous fungi, especially hyphomycetes, occurring on leaf litter. An overview of these studies is given here.
Subramanian [391,392,393,394,395,396,397], in his series of seven papers “Fungi imperfecti from Madras”, reported several new palmicolous hyphomycetes from dead palm leaves in Chennai. Moreover, in addition to his previously mentioned reports on dematiaceous hyphomycetes [259,262,266,289,294,308,309,327,328], Subramanian extensively studied hyphomycetes in tropical regions, especially India. These studies resulted in a major monograph of the Indian species [398], many of which were collected from palm material and introduced as new to science [399,400,401,402,403,404,405,406]. A number of these new palmicolous hyphomycetes were introduced in his series of six papers entitled “Hyphomycetes”, where new genera and species were described on the leaves of several different palm species, such as Cocos nucifera, Phoenix canariensis, Rhopalostylis sapida, and Borassus flabellifer from Tamil Nadu [407,408,409,410,411,412].
Rao and Rao [413,414,415,416,417,418,419,420], Chaudhury and Rao [421], and Rao and Chaudhury [422], also reported several palmicolous fungi, including new taxa, from different palm species, such as B. flabellifer, Caryota urens, C. nucifera, and Livistona chinensis, mainly from Hyderabad. Moreover, new species of palmicolous fungi were reported by Rao [423,424,425] from Maharashtra. Later, Varghese and Rao [426,427] recorded several palm fungi during their mycological survey of the forests of Kerala, near where Pande and Rao [428] collected the new species Rosellinia lakshadweepensis on the pericarp of C. nucifera from the island of Kavaratti (Lakshadweep).
Bhat and co-workers isolated some new taxa of palmicolous litter-inhabiting hyphomycetes in palm collections from India in their studies on fungi from the forests of the Western Ghats hills in Goa, the rainforests of the Andaman-Nicobar Islands and, to a lesser extent, the humid mountains forests of Northeastern Himalayas, e.g., refs. [429,430,431,432]. In addition to these taxonomic studies, ecological studies were also carried out on litter colonisers and endophytes in plant species from the forests of the Western Ghats in Goa, which included palm trees species, such as Calamus thwaitesii. C. urens, and Elaeis guineensis [433,434].
A vast area of the peninsular India still remains underexplored for fungal biodiversity [435]. However, the fungi of the forests of the Western Ghats in Goa and the rainforests of the Andaman-Nicobar Islands, two of the biological hotspots of India, have been explored to some extent, e.g., refs. [436,437,438,439,440,441]. In turn, many palm fungi have been documented, including several new taxa (Figure 1, Table 1). Most of these studies were reviewed by Bhat [437], who considered the forests of the Western Ghats as “an abode of novel and interesting microfungi” and presented a list of new hyphomycetes discovered in them, where several palmicolous fungi are cited. Further studies on the diversity of microfungi from these forests in Goa and in some parts of Karnataka, Kerala, Tamil Nadu, and Maharashtra were carried out by Pratibha et al. [442,443], which resulted in the documentation of a few more palmicolous hyphomycetes. Still, today studies on the fungi of the forests of the Western Ghats hills and the rainforests of the Andaman-Nicobar Islands continue to be carried out and to report an exceptional diversity of palm fungi. For instance, Dubey and Moonnambeth [306,444,445,446,447,448], Dubey [449], and Dubey and Neelima [450] documented several dematiaceous palmicolous hyphomycetes, including new taxa, during an investigation of fungi from the forests of the Western Ghats of Maharashtra (Table 1). More recently, Niranjan and Sarma [451] compiled a checklist of fungi reported from the Andaman-Nicobar Islands, where many palmicolous fungi have been documented. Several other palmicolous fungi, including new species, have been reported from the rainforests of these islands by Ram and Sinha [452] and Niranjan and Sarma [453,454,455,456,457].
Although information on Indian palm fungi is scarce and difficult to review and compile properly, fungi collected from living palm leaves, diseased palm foliage, and palm litter by various researchers have been included in periodically published lists and compilations of Indian fungi, for example “List of Indian fungi 1952–1956” [404], “List of Indian fungi 1956–1960” [458], “Fungi of India 1989–2001” [459], “Ascomycetes of Peninsular India” [460], and, more recently, “Bilgrami’s Fungi of India List and References (1988–2020)” [461]. One of these compilations was dedicated exclusively to palm fungi under the title “Fungal records on palms from India” by Pande et al. [462], who made a list of fungi recorded on palm trees up to 1999 and listed 355 species distributed in 188 genera described from 29 species of palm trees from different Indian states.
Brazil is a country rich in palm species, some of which have great socio-economic value [463]. Even so, few taxonomic studies have been carried out on Brazilian palm fungi (Figure 1) and most of them have focused on palm hyphomycetes. However, the few studies available have also revealed the trend towards the presence of a rich fungal diversity, from which many new palmicolous fungi have been introduced. A summary of some examples of these studies is given herein. In 1978, Hennen and Ono [464] identified the first rust fungus on a palm tree, the new genus and species Cerradoa palmaea on Attalea ceraensis from Brasília. Several palm fungi were gathered from collections of Astrocaryum from Amazonas and published by Farr [465,466,467] in his series of papers “Amazonian foliicolous fungi”. Later, Rodrigues and her colleagues recorded some new palmicolous taxa when studying the endophytic fungi that inhabit the tissues of the Amazonian palm Euterpe oleracea [291,468,469,470,471,472,473] (Table 1). Rodrigues [468,472] published the first study on the fungal endophytes inhabiting the foliage of E. oleracea from Combu Island, growing in the Brazilian Amazon estuary and in Amazonian floodplains, and listed 57 species of palm ascomycetes, including several hyphomycetes. In addition, several fungi recorded on arecaceous hosts from Brazil have been compiled by Silva and Minter [474] and Mendes et al. [475,476].
Many other sparse reports of palm fungi from Brazilian regions have been made. However, similar to the studies on palm fungi from Argentina, most of the studies on Brazilian palm fungi were not carried out as systematic studies of descriptive taxonomy. Instead, these studies aimed to better understand the diversity of conidial fungi, especially hyphomycetes, on woody palms of the Amazon rainforest and other biomes, where biodiversity research and conservation programs were being conducted. An overview of these studies is presented here.
The investigation on conidial fungi associated with the decomposition of palm leaves in the Amazon rainforest has been restricted to three areas, namely the National Forest of Caxiuanã [274,387,477,478,479,480,481] and the Combu Island [482,483] in Pará, and the National Forest of Amapá in Amapá [264,387,479,481]. Several Arecaceae hosts have been sampled, including members of Astrocaryum, Attalea, Bactris, Euterpe, Geonoma, Maximiliana, Oenocarpus, Socratea, and Syagrus. Most of these studies were motivated both by the accelerated deforestation of the Amazon rainforest and by the lack of research into the diversity of palm fungi in the tropical regions of South America. In addition to the importance of these studies for expanding the knowledge about the composition and distribution of palm fungi in the Amazon rainforest, several new records for these Brazilian regions, as well as new genera and species, have been reported, e.g., refs. [264,274,387,479] (Table 1).
Surveys of microfungi on palm trees have also been carried out in Bahia, Brasília, and Pernambuco, particularly in biomes in areas of Northeast Brazil where palm trees are a major component [484,485,486,487,488,489,490,491,492,493,494,495,496,497,498,499,500,501,502,503,504,505,506,507,508,509,510,511,512,513,514,515,516]. These studies aimed to understand the fungal composition of important biomes of Northeast Brazil, such as the Atlantic rainforest, e.g., refs. [496,499,500], the Caatinga, e.g., refs. [497,501,502,503,504,507,508,509,512], and the Cerrado, e.g., refs. [485,486,489], as well as to uncover the mycota that inhabit the tissues of important palms that can be found there, including Acrocomia intumescens, Attalea funifera, Bactris acanthocarpa, Cocos nucifera, Elaeis guineensis, Euterpe edulis, E. oleracea, Mauritia flexuosa, Polyandrococos caudescens, Syagrus botryophora, and S. coronata. In turn, several new palmicolous taxa have been reported, including members of the typical palmicolous mycota found by Hyde and co-workers on their extensive investigation on East and Southeast Asian and Australasian countries. For instance, Vitória et al. [491,495] introduced the new species Arecomyces attaleae and Neolinocarpon attaleae on dead rachides of A. funifera from Bahia.
2.1.5. Palm Fungi and Reflections on the Recent Input from Molecular Era
This comprehensive overview of the literature shows that palms support a vast array of fungi, especially ascomycetes. In the well-studied tropical regions of East and Southeast Asia, Australasia, and, to a lesser extent, Central America (Figure 1), where these fungi have received considerable attention, a remarkable diversity of fungi has been revealed with the description of numerous new taxa, e.g., refs. [6,8,104,329,352]. Much of this diversity can be attributed to the tropical and subtropical habitats surveyed, where the diversity of fungi is known to be higher [10]. However, the few studies carried out on palms that thrive in temperate regions have also revealed a considerably rich fungal diversity, of which some taxa have been described as new to science. For instance, several palmicolous fungi have been recorded in New Zealand (Figure 1), particularly from collections of Rhopalostylis spp. [54,517,518,519,520,521]. McKenzie et al. [54] noted that 147 named fungal species and 50 fungal records identified only to genus have been recorded on Rhopalostylis, mainly on R. sapida from New Zealand. Thus, the great diversity of palmicolous fungi recovered can be also attributed to the wide variety of palm hosts and habitats studied, including many different palm species and tissues in terrestrial, freshwater, and marine or mangrove ecosystems. While the first studies by Hyde and co-workers covered the diversity of fungi on mangrove palms, e.g., ref. [168], most subsequent studies were dedicated to surveying saprobic fungi that develop on palm substrata from tropical rainforests, e.g., ref. [150]. In addition, few studies were conducted to survey palm phytopathogens, e.g., ref. [212].
Up to 2003, the intensive research carried out by Hyde and co-workers has reported more than 320 new species and more than 45 new genera of palmicolous fungi. However, in all these earlier studies, all the taxa reported were introduced, described, and arranged in different taxonomic ranks within the Ascomycota based solely on their morphology. This is currently known as a subjective approach and many taxa have been assigned to Ascomycota genera incertae sedis. Given that palm trees are important hosts that harbour potential novel taxa, it is critical that these palmicolous fungi are recollected, epitypified where needed, and isolated so that molecular data can be obtained and used to establish their natural phylogenetic placements [522]. For example, regarding the initial examples of some of the most common fungal genera found on palms, such as Anthostomella, Astrosphaeriella, Linocarpon, and Oxydothis, several species have been recorded on palms, e.g., refs. [111,134,138], but only a small percentage of these have associated sequence data available. As a result, their position in a natural taxonomic framework is poorly supported and, consequently, their evolutionary relationships in higher taxonomic ranks are poorly understood.
Some of the first studies on palmicolous fungi that combined molecular data with morphological data were those by Pinruan et al. [270,282,301,307,523] and Pinnoi et al. [524,525] in Thailand, who reported several new taxa of saprobic fungi on Calamus species and on the peat swamp palms Licuala longecalycata and Eleiodoxa conferta. These studies were a continuation of previous solely morphological studies that had already yielded a remarkably rich fungal diversity from palms in the Sirindhom Peat Swamp Forest at Narathiwat, Southern Thailand [236,241,242,243,244,245,246,247]. Four new genera have been introduced (Table 1) and their phylogenetic relationships among extant taxa have begun to be unveiled. An interesting example worth mentioning is the description of the new species Astrocystis eleiodoxae on petioles of E. conferta submerged in a peat swamp from Thailand by Pinnoi et al. [525]. Much of the early studies by Hyde and co-workers focused on the treatment of several genera that occur on palms and their taxonomic relationships and placement based mainly on morphological characters related to asci, ascospores, and associated features. The taxonomic placement of the genus Astrocystis, as well as several other xylariaceous-related genera commonly recorded on palms, has been analysed, although it has often remained obscure due to the assessment of its morphology alone, e.g., ref. [150]. The description of A. eleiodoxae by morphological and phylogenetic means, as well as other new xylariaceous taxa from palms, such as Rosellinia capetribulensis on decaying rachides of Calamus sp. from Northern Queensland, Australia by Bahl et al. [526], underlines the importance of introducing molecular data into the study of palm fungi as a highly diverse fungal group. Although the assignment of these taxa to higher taxonomic ranks remained ambiguous and undetermined, the relationship with other xylariaceous genera started to become clearer than when these studies were conducted solely on the basis of morphology.
The introduction of DNA sequence data to study fungal biodiversity in the early 1990s has served as a stimulus for the description of new taxa from palms. Hence, several of the old collections have been accessed and their taxonomic placement clarified. Over the last 15 years, Hyde and co-workers have been revisiting their studies on palms, recollecting and epitypifying some of the taxa and clarifying their phylogenetic position among extant and new taxa [522]. This, in turn, has refined the identity of the most common mycota that inhabits palm tissues by providing information on the natural grouping of palmicolous genera based on sequence analyses (Table 2). The number of reports has been increasing rapidly, especially in collections from Northern Thailand (Figure 1), revealing several new palmicolous taxa that have complexified the taxonomic structure of palm fungi (Table 1 and Table 2).
Several new species of the common palm mycota were introduced, along with new genera, including members of the Sordariomycetes and Dothideomycetes frequently found on palms. This ultimately led to the establishment of new families to accommodate and clarify their phylogenetic relationships. Indeed, new taxa of some of the most common fungal genera found on palms, including Oxydothis [527,528,529], Linocarpon, Neolinocarpon [530], Astrosphaeriella sensu lato [267,281,531,532,533], and Roussoella [534], are continuously reported on palm tissues and the new families Oxydothidaceae [528], Linocarpaceae [530], Astrosphaeriellaceae [267], and Roussoellaceae [534], respectively, have been introduced to accommodate them (Table 2). A historical account of molecular studies carried out on palm fungi is presented herein. In addition, some case studies of common palm taxa are highlighted to reflect on the impact of molecular data on the taxonomy and biodiversity of this group of fungi.
Astrosphaeriella-like Taxa: A Polyphyletic Nature Hiding Cryptic Genera
Astrosphaeriella is an interesting case study of how phylogenetics and its inputs had implications in the taxonomic structure of palm fungi. Although there have been many morphological-based studies of Astrosphaeriella, including those major morphological studies by Hyde and Fröhlich [144] and Hyde et al. [148], based particularly on species of Astrosphaeriella occurring on palms in tropical regions, no thorough molecular investigation of the genus has been carried out. The taxonomy of Astrosphaeriella and its natural placement, especially at family level, stayed unresolved until recently. The frequent collection of Astrosphaeriella-like species on palms and other monocotyledonous trees has made it possible to move towards their natural classification. Thus, in addition to understanding the polyphyletic nature of Astrosphaeriella sensu lato, Liu et al. [281] erected two new genera of Astrosphaeriella-like species in Aigialaceae, viz. Fissuroma and Neoastrosphaeriella (Table 1). Later, Phookamsak et al. [267] recognised that Astrosphaeriella-like species can be distinguished into three families and established Astrosphaeriellaceae for typical Astrosphaeriella species (sensu stricto), Pseudoastrosphaeriellaceae to accommodate the new genus Pseudoastrosphaeriella, and the new genus Astrosphaeriellopsis for a distinct Astrosphaeriella-like lineage basal to Aigialaceae (Table 1). Astrosphaeriellopsis was later accommodated in Astrosphaeriellaceae by Wanasinghe et al. [532] following a multigene phylogeny and respective taxonomic circumscription of Astrosphaeriella species and allied genera with the introduction of several novel palmicolous taxa in the genera Astrosphaeriellopsis, Fissuroma, Neoastrosphaeriella, and Pithomyces isolated from Calamus, Caryota, and Licuala species in Northern Thailand and Southwest China. Further additions to Fissuroma and Neoastrosphaeriella were made by Konta et al. [533] and Zhang et al. [531] from palms in mangrove and terrestrial habitats in Thailand. Another genus, Xenoastrosphaeriella (Table 1), was introduced in Astrosphaeriellaceae to accommodate saprobic fungi on bamboo and palms that previously represented a basal lineage in Astrosphaeriellaceae [535]. More recently, the new genus Triseptatospora was introduced in Astrophaeriellaceae to accommodate T. calami found on dead petioles of Calamus sp. in Thailand [310] (Table 1).
Several new Dothideomycetes, along with Astrosphaeriella sensu lato and Roussoellaceae, are also being recorded from palm tree collections. Mapook et al. [536] introduced the new family Palawaniaceae to accommodate the Palawania species, which are saprobes common on palms, occurring on dried fronds and spines. Later, Jayasiri et al. [535] introduced two new species of palmicolous Dothideomycetes in Delitschia and Vaginatispora on fallen fruit pericarp of Nypa fruticans from Thailand. The new genus Pseudopalawania was introduced in Muyocopronaceae by Mapook et al. [303] to accommodate P. siamensis on a dead rachis of Caryota sp. from Thailand (Table 1). Recently, Yu et al. [537] made new additions to Occultibambusaceae based on collections of decaying petioles of Trachycarpus fortunei from China. Several aquatic Dothideomycetes, as well as Sordariomycetes, have also recently been described from palm trees and will be discussed later.
Xylarialean and Related Sordariomycetes: The Enigmatic Anthostomella and Allied Genera
Regarding Sordariomycetes, along with Oxydothidaceae and Linocarpaceae, many other new ascomycetes are being introduced and their phylogeny resolved based on collections from palms. However, several palmicolous genera are poorly represented with sequence data. In addition, several sequences are of poor quality, which can compromise the resolution of the phylogenetic placement of taxa, especially at higher taxonomic levels, such as family and class [538]. This has often been observed in studies on palm fungi. For example, while introducing Linocarpaceae to accommodate Linocarpon and Neolinocarpon, Konta et al. [530] also introduced the new family Leptosporellaceae in Chaetosphaeriales to accommodate Leptosporella, including two new species collected from palms, and provided a comparative morphological list of species in Leptosporella, Linocarpon, and Neolinocarpon. However, as the authors stated, fresh collections of several of the known taxa with associated molecular data are needed to establish and strengthen their natural phylogenetic placements, since all the diagnosed clades are sparsely populated. Similarly, a checklist of Diatrypaceae that occur on palms was presented by Konta et al. [263], with the introduction of many new species and a new genus, Allodiatrype (Table 1). However, the generic taxonomic resolution of several lineages remained unclear, probably due to a lack of sequence data or previous misidentifications, so their phylogenetic placement will only be clear with fresh collections and adequate sequence data [538].
The need to recollect and isolate some of the old collections of palm fungi was also reinforced by Daranagama et al. [539], who re-examined the type specimens of some Sordariomycetes genera to determine their family placement according to modern taxonomic concepts. As previously mentioned, the taxonomic placement of several xylariaceous genera, including those commonly recorded on palms, is often obscure due to the assessment of their morphology alone and the lack of sequence data to properly place them in a natural taxonomic framework. Moreover, novel taxa are often introduced when new collections and their respective molecular data are obtained. For instance, Konta et al. [293] introduced the new genus Neoxylaria (Table 1) to accommodate the new species N. arengae, as well as the morphological species Xylaria juruensis (as N. juruensis) from palm material in Brazil and X. queenslandica (as N. queenslandica) from Archontophoenix alexandrae in Australia, using both their fresh collection of N. arengae and morphology data, as there is no molecular data for these old collections. More recently, two new families were introduced for genera that frequently occur on palms, along with the introduction of new species. Appendicosporaceae was introduced to accommodate Appendicospora, with the analysis of a new fresh collection designed as a reference specimen for A. hongkongensis on dead fronds of Livistona chinensis from China [540]. Fasciatisporaceae was introduced to accommodate Fasciatispora, with the introduction of the new species F. cocoes on decaying rachides of Cocos nucifera from Thailand [541].
Several species of Fasciatispora have been introduced from palm trees since the first studies by Hyde et al. in the 1990s, and a synopsis of Fasciatispora species, along with Anthostomella, from decaying palm fronds collected in Indonesia and Thailand was presented by Hidayat et al. [542]. However, only a few of them have molecular data available for inclusion in modern taxonomic concepts and most of the new species from the 1990s remain described based only on their morphology. Recently, the type species of the genus, F. nypae, was re-collected on a frond of Nypa fruticans from Thailand and a reference specimen was designated, which made it possible to begin to resolve the phylogeny of Fasciatispora as a basal clade in Xylariaceae [543], which was later introduced as the family Fasciatisporaceae [541].
Likewise, several Anthostomella species have been introduced from palm trees and, since its first synopsis by Hyde [133], who accepted twenty-seven species (of which nine were new) occurring on palms, several other palmicolous Anthostomella species have been described, e.g., refs. [143,209,218,219,220,221]. However, the phylogeny of this species-rich genus is still ambiguous. Although its polyphyletic nature has already been recognised, a significant phylogenetic and morphological re-assessment of Anthostomella is still needed, including fresh collections and associated molecular data [539,544,545]. None of the Anthostomella species described from palms have been re-evaluated to gain knowledge about their phylogenetics, so it can be assumed that several new taxa have yet to be described under morphological Anthostomella-like species. In fact, several Anthostomella-like species have been analysed and placed in existing and new genera to properly accommodate them, e.g., refs. [539,541,544,545]. For instance, recently, Konta et al. [284] introduced the new genus Haploanthostomella from palms based on H. elaeidis on dead leaves and rachides of Elaeis guineensis from Thailand (Table 1) and provided a key to genera with Anthostomella-like characteristics. Konta et al. [284] also provided a family replacement of Endocalyx to Cainiaceae. Endocalyx is a genus of coelomycetes almost exclusively reported on palm hosts [546,547]. Although Delgado-Rodríguez et al. [546] made a recent phylogenetic assessment of Endocalyx, some species still lack molecular data, as is often the case with xylariaceous genera.
Palmicolous “Anamorphs”: A Plethora of Botryosphaeriaceae and Other Dothideomycetes
Following the trends of the late 1990s, along with the description of the Ascomycota coverage through the presence of its sexual morphs on the host, reports on palmicolous “anamorphs” began to become more frequent, e.g., refs. [241,243,244,245,246]. In recent years, several species of Botryosphaeriaceae are being introduced as new to science based on palm collections from different regions of the world, particularly species of Neodeightonia [505,547,548,549,550,551,552,553]. Moreover, some new pestalotioid-like species have recently been described on palms, including species of Neopestalotiopsis, Pestalotiopsis, Pseudopestalotiopsis [296,310,554,555,556,557,558,559], Seridium [560], and Morinia [561].
Dematiaceous hyphomycetes, as demonstrated by studies on palmicolous hyphomycetes from Central American countries and India, are also frequently reported on palm tissues and their phylogenetics is continuously revealing new taxa. Li et al. [562] introduced the new family Zygosporiaceae to accommodate Zygosporium, a widespread genus usually associated with monocotyledonous, including palms. Delgado-Rodríguez, in his series of papers on “South Florida microfungi”, in which many new species of palm hyphomycetes were described, e.g., refs. [290,312,563,564,565,566], introduced a new species of Taeniolella on the petiole of a dead leaf of Sabal palmetto, expanding the concept of the strong polyphyly of the genus among different classes [567]. Later, Delgado-Rodríguez and co-workers introduced the new genus Castanedospora to accommodate Sporidesmium pachyanthicola based on an epitype specimen collected on the petiole of a dead leaf of S. palmetto, redefining its placement at family level in the Extremaceae [273] (Table 1). Moreover, the phylogenetic placement of Ernakulamia cochinensis, one of the saprobic hyphomycetes taxa commonly found associated with palm hosts, as a member of Tetraplosphaeriaceae in Pleosporales, was investigated based on a representative specimen collected on Astrocaryum standleyanum in Panama [568]. Species of Hermatomyces are being described on palms from different regions of the world, including Panama [569], Thailand [570], and Texas (USA) [571], in addition to other exclusively morphological studies that have reported Hermatomyces species from palm trees collections in India [572] and Sierra Leone [573] (Figure 1, Table 2). More recently, Konta et al. [285] described Helminthosporium-like taxa from palms in Thailand and introduced two new genera in Massarinaceae, viz. Haplohelminthosporium and Helminthosporiella (Table 1). Konta et al. [285] also provided a checklist for Helminthosporium reported worldwide and most Helminthosporium species described from palms were based on morphology alone and only one species, H. livistonae on leaves of Livistona australis from Australia [302], was based on both morphology and sequence data. Chen et al. [574] also introduced the new species H. chinense on a decaying branch of an unidentified palm tree from China.
Many other occasional reports of palmicolous “anamorphs”, including new genera and species, are continuously published, revealing the plethora of genera that make up the hyphomycetous and coelomycetous assemblage that inhabit palm tissues, e.g., refs. [268,279,310,575,576,577,578] (Table 2). For example, Hongsanan et al. [279] described the new genus and species Discopycnothyrium palmae on the branches of an unidentified palm from Thailand (Table 1). A number of new hypocrealean members have been recently introduced based on palm collections from French Guiana, including species of Chaetopsina [579], Clonostachys [580,581], Hydropisphaera [582], Ijuhya [583,584], Lasionectria [585], and Volutella [586]. Several novel anamorphic chaetosphaereaceous fungi, including new genera, were reported from palm collections from China [268,575,577,578] and Thailand [310] (Table 1).
Crane and Miller [587] introduced some new species in Torula-like genera from palms, such as Bahusaganda and Bahusandhika, but no molecular data was obtained to establish their natural placements. Although the importance of acquiring sequence data to identify fungal taxa is now widely recognised and advisable [588,589,590], there are still several publications that have been and continue to be published without the support of molecular methods, introducing new morphological palmicolous taxa. For example, Wulandari et al. [591], while synopsising the species of Phyllosticta (as Guignardia) described from palm trees, introduced two new species from Northern Thailand based on morphology. Lechat and Fournier [592] introduced the new species Lasionectria marigotensis on a decaying leaf of Cocos nucifera from Guadeloupe (French West Indies) by comparing its morphology with a previous collection of Lasionectria, L. calamicola, from palms in Australia and Brunei Darussalam by Fröhlich and Hyde [6]. Later, the new species Dictyocheirospora indica (as Dictyosporium indicum) was collected on a petiole of Phoenix rupicola from India [593] and the new species Endophragmiella licualae was collected on dead branches of Licuala fordiana from China [594]. More recently, several palmicolous fungi, including new species, have been reported from the rainforests of the Andaman-Nicobar Islands (India) by Niranjan and Sarma [453,454,455,457] based on morphology alone, including important palmicolous genera whose taxonomic resolution highly relies on DNA sequence data, such as members of Aigialaceae, viz. Fissuroma and Neoastrosphaeriella, and Astrosphaeriellaceae, viz. Astrosphaeriella. Likewise, as already mentioned, many new species and genera of palmicolous hyphomycetes have been described on the basis of their morphology in Central and South American countries, including Mexico, Argentina, and Brazil.
The consistent new discoveries of fungi from palm hosts suggest their considerable potential for the identification of novel fungal taxa. Thus, using morphology alone to introduce new palmicolous taxa is strongly discouraged here. Although many fungal species have unique and distinctive characters that make them easily identifiable, especially in genera of hyphomycetes, the existence of cryptic species and species complexes has highlighted the importance of molecular methods in fungal identification [590,595]. In fact, the morphological species concept is thought to underestimate the number of species, since morphological characters can be very plastic (phenotypic plasticity) and often defines groups of cryptic species [590].
From Aquatic to Phytopathogenic Fungi: The Broad Taxonomic Spectrum of Palm Fungi
Freshwater and marine or mangrove ecosystems have also revealed a remarkable diversity of new palmicolous taxa with the incorporation of phylogenetics into the previous regular morphological studies [9,261,269,278,596,597,598,599,600,601,602,603,604]. The mangrove palm Nypa fruticans has proven to be a very distinctive habitat for fungal research since 1988, when Hyde began his studies to understand the assemblage of fungi that colonise palm tissues [154,162]. In fact, many new palmicolous fungi are continuously being described from N. fruticans. For instance, Zhang et al. [261] introduced the new genus Acuminatispora isolated from decayed petioles and rachides of palms in mangrove habitats in Thailand, including N. fruticans and Phoenix paludosa. More recently, Jones et al. [604] described the new species Salsuginea phoenicis on a decaying petiole of P. paludosa from the intertidal zone in Thailand. Thus, mangrove and peat swamp palms have been the substratum for the collection of various new aquatic fungi (freshwater and/or marine ascomycetes), whose phylogenetic resolution has often led to the description of new families and genera. In fact, four families have been recently introduced to accommodate new and extant taxa of aquatic fungi with affiliations to palm habitats. Falciformispora and Trematosphaeria, with representatives found on palms, were assigned to the new family Trematosphaeriaceae [596,598]. Suetrong et al. [599] introduced the new family Tirisporellaceae in Diaporthales to accommodate the genera Tirisporella and Thailandiomyces collected from palms in freshwater streams or peat swamps, and Abdel-Wahab et al. [269] added the new genus Bacusphaeria isolated from the petiole base of N. fruticans from Malaysia. The new family Dictyosporiaceae was established to accommodate saprobic fungi that occur on decaying wood and plant debris in terrestrial and freshwater habitats, including Dictyosporium, often found on palm trees, and the new genus Dictyopalmispora described from decaying leaves of Licuala longecalycata in a peat swamp forest in Thailand [278,602,603] (Table 1). Later, Zhang et al. [9] introduced another new pleosporalean family, Striatiguttulaceae, to accommodate two monophyletic lineages described from decayed rachides of N. fruticans and P. paludosa, the new genera Longicorpus and Striatiguttula.
In the last decade, microfungi that occur as saprobes, pathogens, and endophytes on palm hosts are commonly introduced and reanalysed in mycological series, such as Fungal Diversity notes, e.g., refs. [296,605,606,607,608], Fungal Planet description sheets, e.g., refs. [609,610,611,612,613,614,615,616], and Mycosphere notes, e.g., refs. [617,618,619,620], as well many other similar publications, e.g., refs. [621,622,623,624,625,626,627,628,629]. A remarkable taxonomic novelty published recently based on a palm collection was the new order Pararamichloridiales introduced to accommodate the new family Pararamichloridiaceae based on the new genus and species Pararamichloridium livistonae on leaves of Livistona australis from New South Wales, Australia [298]. Therefore, these occasional publications continue to expand both the broad taxonomic spectrum of palm fungi and the importance of studying them as phytopathogens and other ecological groups. In fact, in recent years, a series of new palmicolous phytopathogens have been introduced and have highlighted the lack of knowledge about the ecology of palmicolous fungi. For instance, the new species Cercospora arecacearum has been found associated with necrotic leaflets of areca palms in Thailand by To-anun et al. [630]. Kinge and Mih [631] described the new basidiomycete Ganoderma ryvardenii (as G. ryvardense) associated with basal stem rot disease of oil palm in Cameroon, one of the main production constraints faced by agro-industries and smallholders’ farmers in oil palm production. Moreover, Mbenoun et al. [632] described the new species Thielaviopsis cerberus (as Ceratocystis cerberus) on the stump of a felled Elaeis guineensis tree from Cameroon, while studying species boundaries in the C. paradoxa complex, a serious constraint to the cultivation of monocotyledonous crops, including the trunk rot affecting almost all palm species. Considerations on the importance of these diseases as one of the main causes of losses in oil and ornamental palms have been put forward by Aiello et al. [633], while introducing the new phytopathogen species Ilyonectria palmarum. More recently, the new genus and species Palmeiromyces chamaeropicola was collected from diseased foliage of Chamaerops humilis from Portugal [295], revealing a new insight into Teratosphaeriaceae leaf diseases, which are caused by important phytopathogens of various plant hosts.
Research into palm fungi continuously reveals the importance of the Arecaceae family as host plants to search for novel taxa. In addition, recent studies have been important in showing the imperative need to apply molecular data to resolve the taxonomic structure of palm fungi as an ecologically diverse and important assemblage.
2.2. History of Biodiversity and Ecological Studies on Palm Fungi
Palm trees have proven to be a diverse habitat, exhibiting intense fungal colonisation. Most studies on microfungi that colonise palms are taxonomic, insomuch as they have primarily focused on cataloguing fungi and describing new taxa collected on palm substrata from various regions of the world, especially in the tropics (Figure 1). However, there are a number of studies on the ecology of palm fungi, which have focused on different aspects of their biodiversity, mainly with regard to saprobes and endophytes. Although the approach of these studies is different, the description of new taxa has often resulted from initially ecological approaches that yielded several interesting fungi for further analysis. This suggests that both approaches are essential to explore the knowledge on fungi and their biodiversity. In fact, the implications of biodiversity and ecological data on the global numbers of fungi and their general knowledge have been pointed out, which will be discussed later in relation to palm fungi and their key role in biodiversity surveys.
A review of the literature on the ecology of palm fungi reveals that studies have been conducted over the past three decades. These have mainly resulted from the intensive research carried out by Hyde and his co-workers. They have made substantial contributions to knowledge of both the taxonomy and biodiversity of palm fungi, not only by identifying the fungal assemblage that occur on palms, but also by exploring the extent of their diversity and the factors that affect it. However, some ecological studies have also been carried out on Indian palm fungi, as well as palm fungi from Central American regions. A historical account of research into the ecology and biodiversity of palm fungi is presented herein.
Fröhlich and Hyde [10] studied the biodiversity of palm fungi in the tropics, forecasting that the estimate of 1.5 million species would be a “very conservative estimate of the number of fungal species extant on the planet”. Taylor et al. [12] investigated the biogeographical distribution of microfungi from temperate and tropical palms. They revealed that differences in fungal assemblages were more related to climatic influences than to the hosts sampled, as well as to the status of these hosts at the site sampled. Subsequently, Yanna et al. [13,14,634] assessed the composition of palm fungal communities and their succession over time, pointing out that differences in fungal assemblages could be related to different collection sites, hosts, stages of decomposition, and tissues sampled. Later, Pinnoi et al. [16] studied saprotrophic fungal communities associated with Calamus spp. and reported differences in the assemblages of fungi inhabiting different microhabitats and tissues, with dry petioles supporting a greater diversity of species.
Several studies have been dedicated to endophytic palm fungi, which was one of the first ecological issues to be investigated in palm fungi [10,11,468,469,472,473,635,636,637,638]. These studies often report significant differences in the number of isolates and the taxonomic composition of fungi in respect to the plant growth stages, season, site, and tissues sampled, not only in tropical palms, but also in temperate palms, such as Trachycarpus fortunei [215]. Even so, research into palm endophytes, which began in the early 1990s, was primarily motivated by the lack of knowledge about endophytes in plants from tropical regions when compared with those in plants from temperate regions [472,473].
The first study on palm endophytes was by Rodrigues and Samuels [469], who documented the occurrence of endophytes inhabiting the leaves of the Australian fan palm Licuala ramsayi, pointing out preliminary differences in the endophyte assemblages recovered from different parts of L. ramsayi leaves. In addition, the existence of a fungal assemblage composed mainly of xylariaceous fungi was documented. This was also later documented by Rodrigues and her colleagues when they published the first studies on the fungal endophytes that inhabit the foliage of the Amazonian palm Euterpe oleracea from Combu Island (Belém, Pará). In fact, some of these studies were based on systematic and descriptive taxonomy, particularly for xylariaceous species commonly recorded as endophytes of palm tissues, including Xylaria and Idriella [291,470,471]. However, a comprehensive ecological approach was conducted by Rodrigues [468], who found that colonisation of E. oleracea by endophytes was positively correlated with leaf age, plant growth stages, site, and season, as well as with the interactive effect of some of these factors. Later, a similar study was carried out by Fröhlich et al. [11], who investigated the endophyte communities inhabiting different parts of the fronds of L. ramsayi individuals from Australia and Brunei. They noted differences between the fungal assemblages of different palm tissues and tissues with different ages. Likewise, Taylor et al. [215] observed differences in the endophytic communities recovered from different tissues of the frond of the temperate palm Trachycarpus fortunei. They also documented important information regarding the effect of climate on endophytes assemblages by analysing individuals growing inside and outside their natural geographical range.
The importance of these studies and their implications for fungal biology and biodiversity rapidly increased interest in unveiling more details of the ecology of palmicolous endophytes. Molecular data began to be applied in such studies and made endophytes one of the main issues of palm fungi to be explored until recent years [523,639,640,641,642,643,644,645,646,647,648]. Subsequently, the array of questions has diversified greatly. While some studies have invested in documenting the maximum possible diversity of palm endophytes communities, others have targeted particularly important palms and the impacts that endophytic communities can have on their pests and diseases. For instance, Guo et al. [639,640,641] tried to decrease the percentage of sterile mycelium that often lacks identification in endophyte studies by using both morphology and rDNA sequences in a study of endophytes on Livistona chinensis fronds. Later, Rungjindamai et al. [643] and Pinruan et al. [523] used both morphology and molecular techniques to characterise basidiomycete endophytes isolated from the leaves, rachides, and petioles of the oil palm Elaeis guineensis, and to reflect on how these fungi can be used as a biocontrol management strategy against the palm pathogen Ganoderma boninense. Similarly, Mahmouda et al. [644] examined the endophytic fungal diversity associated with the roots of the date palm Phoenix dactylifera growing in coastal dunes to generate a collection of strains that can be used as biocontrol agents against date palm root diseases. More recently, Azuddin et al. [646] studied the fungal endophytes on the spines of Calamus castaneus and evaluated their antagonistic activity against phytopathogens.
Although the importance of palm pathogens and their management are often mentioned, particularly in palms that are important in international trade, relatively few studies have focused on these fungi and their ecology, e.g., refs. [199,212]. Most publications on palmicolous pathogens have been motivated by systematic and descriptive taxonomic studies that have identified new taxa associated with palm diseases, e.g., refs. [187,195,196,197,198,208,295,556,630,631,632,633]. In fact, although the main fungal diseases that affect palm trees have virtually all been identified, e.g., refs. [649,650,651] little is known about minor pathogens, including those that cause leaf spots, as shown by the investigation of Hyde and co-workers. Recently, Douanla-Meli and Scharnhorst [551], while describing botryosphaeriaceous taxa associated with palm foliage from Mexico, reflected on the risk these taxa may pose to temperate countries, since palm foliage can be an import route for potentially phytopathogenic fungi. In addition, a number of studies have addressed the identification of fungal pathogens associated with P. dactylifera in regions where this palm is an important agricultural crop, e.g., refs. [652,653,654,655,656].
Ecological studies on palm fungi are often directed at palm species that are important world crops in international trade. For example, Asensio et al. [657] investigated the mycobiota of the phylloplane of the date palm P. dactylifera and their interactions. Kirkman et al. [658] studied the diversity and ecological association of the oil palm E. guineensis fungal microbiome across root, rhizosphere, and soil compartments, while Seephueak et al. [659] studied the diversity of microfungi that occur in different tissues of the oil palm frond litter in a plantation in Southern Thailand. Very few studies on palm fungi have explored fungal diversity in roots and, as a result, palm root fungi are virtually unknown. However, few ecological studies on palm fungi have addressed the biodiversity of arbuscular mycorrhizal fungi (AMF) and reflected on the importance of these symbiotic associations to the health and growth of some palm species, such as Attalea speciosa [660], Coccothrinax spp. [661,662,663], Cocos nucifera [664], Desmoncus orthacanthos [665], Metroxylon sagu [666], and P. dactylifera [667,668].
Many ecological studies have focused on the biodiversity of fungi from peat swamp [248,249,669,670] and mangrove palms [7,162,671,672,673,674,675]. Such studies often reflect on the assemblage of freshwater and marine fungi that inhabit palm tissues, compared with the fungi typically associated with terrestrial palms. In addition, the presence of different fungal assemblages from different microhabitats and palm trees tissues is often addressed. For instance, Pinnoi et al. [248], studying the fungal biodiversity of Eleiodoxa conferta, reported that fungi are more abundant on the petioles of wet palm material. Similar ecological patterns were reported by Pinruan et al. [249], who indicated that the petioles of dry material of Licuala longicalycata supported the most diverse fungal communities. Furthermore, questions regarding host- and tissue-specificity are frequently addressed, particularly in studies on the brackish water palm Nypa fruticans, whose colonisation by fungi has been well documented [671,674]. Hyde and Alias [7] reported differences in the fungal composition of different palm structures of N. fruticans, including leaves, leaf veins, rachides, petioles, and inflorescences, collected from intertidal and terrestrial habitats. Likewise, Hyde and Sarma [672] reported differences in the fungal assemblages inhabiting N. fruticans along a river, addressing some ecological observations regarding the horizontal and vertical distribution of fungi. One of the most recent publications in the ecology of palmicolous fungi reported the co-occurrence of certain species of fungi on N. fruticans. This study revealed some aspects related to the structure of the fungal communities on tissues of the brackish water palm and helped to understand the dynamics of the ecosystem, suggesting, for example, the potential interaction established between the fungi of these communities [675].
There are numerous studies on palm fungi and a plethora of data has been collected on different aspects of their biodiversity—from their taxonomic and systematic structure to ecological traits of their lifestyles inhabiting different palm tissues, microhabitats, and geographic regions. However, review studies on palm fungi are scarce. Most are confined to proceedings of mycological conferences or chapters in books that explore broader themes, e.g., refs. [17,676,677,678]. With the advent of DNA sequencing, information on palm fungi, particularly their taxonomy and systematics, has become considerably more complex. Thus, defining or understanding them as a complex and diverse group of fungi may not be an easy task. In this sense, the question “what are palm fungi?” arose and will be discussed below.
3. What Are Palm Fungi? Global Figure and Taxonomic Structure
Palm fungi have been widely documented in Australia, Brunei, Ecuador, Hong Kong, Thailand and, to a lesser extent, in Cuba, Mexico, and India (Figure 1). The data obtained indicate that palm fungi are undoubtedly a taxonomically diverse group. However, their precise taxonomic structure within a natural phylogenetic framework still requires further studies, especially those using DNA sequence data. Many publications have outlined the studies that have been carried out on palm fungi, e.g., refs. [8,17,104,676,677,678]. Nonetheless, to date, there is no recent comprehensive review on this group of fungi. Therefore, most of the figures presented are outdated and uncertain, as they are only based on the intensive research carried out in the 1990s.
3.1. Global Figure of Palm Fungi
According to Hyde et al. [17], by 1997 the global figure of palm fungi was ca. 1580 species, including 650 ascomycetes (41%), 270 basidiomycetes (17%), and 660 “anamorphs” (42%), i.e., 400 hyphomycetes (25%) and 260 coelomycetes (17%). However, given the intensive research carried out since then, it is easy to see that these figures are considerably outdated. In fact, the extensive studies carried out by Hyde and co-workers from the early 1990s to the present day have documented ca. 500 new taxa from palms, almost all of them ascomycetes (the term ascomycetes is used here to refer to species of Ascomycota reported through the presence of their sexual morphs in Arecaceae host tissues), in addition to a number of new host records for Arecaceae. Moreover, several new taxa and host records have been reported from other research groups, including, for example, the palmicolous hyphomycetes collected from palms in Cuba and Mexico by Castañeda-Ruiz, Holubová-Jechová, Mena-Portales, Mercado-Sierra, and many other co-workers.
In view of the present comprehensive review of the literature, it is assumed that the global figure of palm fungi is more than 2500 species. Specifications about each group of fungi, i.e., ascomycetes, basidiomycetes, and asexual morphs, must be carefully made. The global figure produced by Hyde et al. in the 1990s noted that around 41% of the fungi described on palms were ascomycetes [17]. As most of their reports since then were ascomycetes, it can be forecasted that the proportion of ascomycetes in the global figure of palm fungi should be higher, ca. 1370 (55%) species. Similarly, Hyde et al. [17] noted that around 42% of the fungi described from palms were asexual morphs. However, research since then has not revealed as many palmicolous “anamorphs” as ascomycetes, so it is considered that this proportion should be lower, ca. 870 (35%) species. As almost no basidiomycete has been described from palms since the investigation carried out by Hyde et al. in the 1990s, it is considered here that the number of basidiomycetes in the global figure of palm fungi is still ca. 270 (10%) species.
Although these numbers may seem nonsensical, trying to pinpoint them has a great impact on our understanding of the taxonomic structure of palm fungi. In turn, they can have implications for the way mycology studies and searches for them, especially when trying to answer central biodiversity questions, such as “where are the missing fungi?” or “how many fungi are there?”. Before diving into what is currently considered to be the ecology and taxonomy of palm fungi, a few considerations should be made.
The lack of molecular data. As previously mentioned, most studies carried out on palm fungi have been exclusively morphological, which means that most of the species, and higher taxonomic ranks, described have not considered molecular data. As is well known, combining molecular data with morphology is essential for establishing a natural phylogenetic framework. Morphology alone is subjective and can mislead species identification or incorrectly assign them to higher taxonomic ranks, disrupting their true identity and phylogenetic relationships. Furthermore, due to the phenotypic plasticity of morphological traits, the global figure of palm fungi is likely to be underestimated or overestimated. The recollection, epitypification, and isolation of these fungi is critical to establish a natural taxonomic framework for palm fungi.
The lack of studies on palmicolous “anamorphs”. Very few studies on palm fungi have focused on asexual morphs, especially coelomycetes. In fact, most studies on palm fungi have studied the Ascomycota coverage through the presence of its sexual morphs on the hosts. Thus, it is not surprising that palmicolous “anamorphs” are less represented in the global figure of palm fungi than ascomycetes “teleomorphs”. Recent reports predicted that the number of asexually reproducing fungi is greater than the number of sexually reproducing fungi [590]. It can therefore be predicted that palmicolous “anamorphs” are highly understudied and, consequently, underestimated. Since palms are fungi-rich host plants, it is expected that many new asexually reproducing species of palm fungi are awaiting to be documented. Furthermore, the above-mentioned numbers clearly demonstrate how far from the truth the global figure of palm fungi is, since more Ascomycota “teleomorphs” have been recorded on palms than “anamorphs”.
The lack of studies on palmicolous basidiomycetes. As with palmicolous “anamorphs”, very few studies on palm fungi concern basidiomycetes. Although it can be assumed that their proportion in palm fungal assemblages is low, given the previous studies that have treated these fungi as communities through their isolation in culture (traditional methodology), almost no study has specifically aimed to evaluate palm basidiomycetes. For example, Pinruan et al. [523] studied the occurrence and diversity of endophytic and saprophytic basidiomycetes on the oil palm Elaeis guineensis in Thailand and confirmed a rich fungal diversity. As the authors discuss, traditional isolation methods are known to hinder the detection of basidiomycetes. Especial concerns about the selectivity of the methods and the temporal and spatial variability of the basidiomycetes’ mycelium should be considered when studying these fungi [523]. Thus, the lack of studies aimed specifically at palmicolous basidiomycetes can be the reason why the proportion of these fungi is so low in the global figure of palm fungi. It can therefore be predicted that palmicolous basidiomycetes are also highly understudied and underestimated.
The difficulty of compiling data. Predicting an exact global figure of palm fungi and their specific groups is a difficult task. Many studies are constantly being published on new species documented on palm substrata and it can be difficult to keep track of them all. Similarly, an overwhelming number of studies have reported on palm fungi by randomly studying certain groups of fungi, hosts, or habitats and compiling these reports is an almost impossible task. Furthermore, the extent of these publications and, consequently, the rich diversity of palm fungi, makes it impossible to use well-known databases such as the US National Fungus Collections Fungus–host Database, which often cannot cope with the number of reports due to the enormous amount of information on palm fungi.
3.2. Taxonomic Structure of Palm Fungi
This overview summarises the main taxa found on palm trees, which typically make up what is referred to as palm fungi. Although the expression “palm fungi” has been used here to denote records of fungi on Arecaceae hosts, it was originally applied to a particular mycota that was consistently found in association with palms in the tropics. Thus, this overview does not represent an exhaustive list of taxa that have been reported from Arecaceae at any taxonomic rank (i.e., species, genera, or higher taxonomic ranks). As a result, some genera already recorded on palms may not be mentioned either because their frequency is not particularly significant, or mainly because they represent ubiquitous and plurivorous taxa, which are associated with several different hosts and present a cosmopolitan distribution (e.g., Alternaria, Aspergillus, Penicillium, and Phoma). The taxonomic structure of palm fungi presented here follows the most recent taxonomic updates and phylogenetic treatments available in the literature (i.e., refs. [313,314,596,621,627,679,680,681,682,683,684,685,686,687,688,689,690,691,692]).
Palm fungi are a taxonomically diverse group with more than 2500 species, including representatives of all the major classes of the fungal kingdom (Table 2). The most representative group of palm fungi is the ascomycetes, a diverse assemblage in which the best represented class is the Sordariomycetes, with four commonly recorded genera, namely Anthostomella (Xylariaceae, Xylariales), Linocarpon (Linocarpaceae, Chaetosphaeriales), Oxydothis (Oxydothidaceae, Amphisphaeriales), and Phomatospora (Phomatosporaceae, Phomatosporales). According to Hyde [111], these correspond to the main genera that invariably colonise fallen palm rachides and leaves in the tropics.
3.2.1. Palmicolous Sordariomycetes
Since the extensive studies carried out by Hyde and co-workers in the 1990s, it has become clear that xylarialean fungi are commonly encountered on palm hosts. In fact, the most well-represented order and family of palm fungi is Xylariales and Xylariaceae, respectively. However, several species of many xylarialean genera still lack molecular data. Thus, to predict their taxonomic structure in Xylariaceae and allied families is often difficult and based in subjective approaches (i.e., comparison of morphological characters). In turn, many of these genera are placed in Xylariales incertae sedis and, therefore, the taxonomic structure of several xylarialean genera of palm fungi is still obscure and needs molecular-based studies [681,682,683].
In addition to Anthostomella, a number of xylarialean genera have been recorded on palms, including Astrocystis, Kretzschmaria, Nemania, Rosellinia, Stilbohypoxylon, Xylaria (Xylariaceae), Biscogniauxia (Graphostromataceae), Idriella (Microdochiaceae), and Hypoxylon (Hypoxylaceae), e.g., ref. [693]. However, according to Smith and Hyde [150], although palm litter is a major component of many lowland rainforests, comparatively few of most of these xylarialean fungi seem to exploit this substratum. Yet, some genera are exceptions to the rule and, in addition to Anthostomella, Hyde and co-workers found that several other xylarialean fungi are common on collections of rainforest palms, particularly clypeosphaeriaceous and diatrypaceous fungi, e.g., refs. [134,263,694]. These include Annulohypoxylon (Hypoxylaceae), Apioclypea, Brunneiapiospora, Palmaria (Clypeosphaeriaceae), Allocryptovalsa, Allodiatrype, Anthostoma, Cryptovalsa, Diatrype, Diatrypella, Eutypa, Eutypella, Frondisphaeria, Peroneutypa (Diatrypaceae), Arecophila, Seynesia, Endocalyx (Cainiaceae), Fasciatispora (Fasciatisporaceae), Neoxylaria (Xylariaceae), Zygosporium (Zygosporiaceae) and many other genera, such as Capsulospora, Circinotrichum, Cyanopulvis, Guestia, Haploanthostomella, Lasiobertia, Nipicola, Palmicola, Pemphidium, Pulmosphaeria, and Sabalicola (Xylariales genera incertae sedis). Several of these genera are found exclusively or almost exclusively on palms and represent morphological genera, whose phylogenetic resolution is still needed for a precise taxonomic structuring of palm fungi, e.g., ref. [539].
Many other Sordariomycetes occur on palms and many genera are considered to be typical in the tropical assemblage of palm fungi, especially in Amphisphaeriales, Chaetosphaeriales, Meliolalles, Phyllachorales, and, to a lesser extent, Sordariales and Diaporthales.
In Amphisphaeriales, apart from Oxydothis (Oxydothidaceae), which is the most commonly found genus of palm fungi, e.g., ref. [528], a great diversity of taxa has been recorded on palms, particularly in Amphisphaeriaceae, e.g., ref. [136], Hyponectriaceae, e.g., ref. [138], and Apiosporaceae, e.g., refs. [125,540]. Several of these genera, like the xylarialean genera mentioned, are almost exclusively palm taxa and include only morphological species, so their phylogenetic resolution is still needed. Amphisphaeriales members occurring on palms include Amphisphaeria, Lepteutypa (Amphisphaeriaceae), Arecomyces, Frondicola, Hyponectria, Rachidicola (Hyponectriaceae), Appendicospora (Appendicosporaceae), Arthrinium, Dictyoarthrinium (Apiosporaceae), Iodosphaeria (Iodosphaeriaceae), and Leiosphaerella (Pseudomassariaceae). In addition to members of Apiosporaceae and Appendicosporaceae, several genera of ascomycetes with unitunicate asci and apiospores are often found on palm tissues [143]. These include some of the xylarialean genera mentioned above (i.e., Anthostomella. Apioclypea, Brunneiapiospora, Palmaria) and other amphisphaeriaceous members, such as Pseudomassaria (Pseudomassariaceae).
In Chaetosphaeriales, the genera Linocarpon, Neolinocarpon (Linocarpaceae), and Leptosporella (Leptosporellaceae) are often found on palms, and their natural placements are beginning to be resolved as new collections are made, although they are still poorly represented with sequence data, e.g., ref. [530]. In addition to these three genera, other Chaetosphaeriales genera commonly recorded on palms include Chaetosphaeria, Chloridium, Sporoschisma (Chaetosphaeriaceae), and Caudatispora (Chaetosphaeriales genus incertae sedis), e.g., ref. [695].
In Phyllachorales, most of the members that occur in association with palms are in Phaeochoraceae, which was introduced to accommodate saprotrophic or biotrophic ascomycetes on plant leaves apparently restricted to Arecaceae hosts, including the genera Cocoicola, Phaeochora, Phaeochoropsis, and Serenomyces, e.g., refs. [212,696]. These genera of Phaeochoraceae, along with members of Phyllachoraceae, including Brobdingnagia, Camarotella, Coccodiella, Coccostromopsis, Maculatifrondes, Malthomyces, Ophiodothella, Oxodeora, Phyllachora, Sphaerodothis and Tribulatia, and Catabotrys (Catabotryaceae, Catabotryales), accommodate species that cause tar spots or lesions on palm leaves and can cause substantial diseases in the hosts, e.g., refs. [212,697].
In Meliolalles, species of Meliola (Meliolaceae) are commonly represented as pathogens on palms, e.g., ref. [6]. In Sordariales, members occurring in association with palms are essentially represented by Cercophora, Lasiosphaeria (Lasiosphaeriaceae) and Lockerbia (Sordariales genus incertae sedis), e.g., ref. [185].
Most of the previously discussed taxa commonly found on palms are represented in two subclasses of the Sordariomycetes, namely Xylariomycitidae and Sordariomycetidae. However, a great diversity of Sordariomycetes is collected from palms and dispersed among many other taxonomic ranks, including a number of members of the Diaporthomycetidae, such as the frequently reported palmicolous genus Phomatospora (Phomatosporaceae, Phomatosporales), and Hypocreomycetidae, such as genera in the Microascales, viz. Triadelphia (Triadelphiaceae) and Wardomycopsis (Microascaceae), as well as other examples that will be discussed below, e.g., ref. [110].
Some genera of the Sordariomycetes recorded on palms are represented by a single or a few collections and their phylogeny is still vague and of uncertain placement within subclasses or orders, for example, Arecacicola, Curvatispora, Nigromammilla, Paracapsulospora, Mangrovispora (Sordariomycetidae genera incertae sedis), Cannonia (Coniochaetales genus incertae sedis), Frondispora, Manokwaria (Xylariomycitidae genera insertae sedis), Myelosperma (Myelospermataceae, Xylariomycetidae family incertae sedis), Neobarrmaelia (Xylariales genus incertae sedis), and Thyridium (Thyridiaceae, Diaporthomycetidae family incertae sedis). Thus, their recollection and epitypification is crucial to establish and clarify their natural placements among extant well-known taxa of Sordariomycetes [681]. In some cases, such as Paracapsulospora and Neobarrmaelia, its natural placement is unclear due to the limited sequence data available to populate surrounding clades [292,296]. Less frequently, members of other subclasses of Sordariomycetes are also reported from palm hosts. For example, members of Conioscyphales, Pleurotheciales, and Savoryellales (Savoryellomycetidae), including the hyphomycete genera Conioscypha (Conioscyphaceae) and Monotosporella (Pleurotheciaceae), and Ascotaiwania, Canalisporium, and Savoryella (Savoryellaceae), respectively, have occasionally been found on palms, e.g., refs. [619,698,699].
In Diaporthales, the members that occur in association with palms are essentially represented by Diaporthe (Diaporthaceae) and mostly restricted to typically temperate palms, although no study has yet dealt with their diversity in depth, e.g., ref. [276]. Other genera of Diaporthales have been recorded on palms, but are represented by single, old collections, whose reanalysis and/or recollection is necessary to properly resolve their phylogeny, namely Apiosphaeria (Diaporthaceae), Coniella (Schizoparmaceae), Maculatipalma (Gnomoniaceae), Durispora, and Phruensis (Diaporthales genera incertae sedis), e.g., ref. [143].
Many Nectria-like and allied species from a wide range of genera are found on palms. These include several members of the Hypocreales, such as Calonectria, Chaetopsina, Cosmospora, Dactylonectria, Ilyonectria, Nectriopsis, Nectria, Ophionectria, Pleiocarpon, Volutella (Nectriaceae), Clonostachys, Hydropisphaera, Ijuhya, Lasionectria, Nectriella (Bionectriaceae), Niesslia (Niessliaceae), Stachybotrys (Stachybotryaceae), Trichoderma, and Verticimonosporium (Hypocreaceae), as well as members of the Magnaporthales, such as Gaeumannomyces (Magnaporthaceae), Ophioceras (Ophioceraceae), Pyricularia (Pyriculariaceae), and Pseudohalonectria (Pseudohalonectriaceae), e.g., refs. [200,207,592]. Likewise, many Acremonium-like fungi and related genera have common representatives on palms, some of which have recently been introduced, including members in Hypocreales, such as Neoacremonium (Neoacremoniaceae), Acremonium, Gossypinidium, Hydropisphaera, Lasionectriella, and Paracylindrocarpon (Bionectriaceae), and Glomerellales, such as Brunneomyces and Acremoniisimulans (Plectosphaerellaceae), e.g., refs. [310,582].
In Tirisporellales, all members are freshwater ascomycetes described from palms, including Bacusphaeria, Thailandiomyces, and Tirisporella (Tirisporellaceae), e.g., ref. [599]. In Annulatascales, the genera Annulatascus and Submersisphaeria (Annulatascaceae), which are typical freshwater taxa, have representatives collected from terrestrial and intertidal palm samples, e.g., ref. [247].
Several genera with common representatives on terrestrial palms also have some marine and freshwater palmicolous species. These have often been recorded on intertidal samples of Nipa palms, which can be wetted daily by tidal inundations and consequently colonised by marine fungi. These include species of Anthostomella, Fasciatispora, Linocarpon, Neolinocarpon, Nipicola, Oxydothis, and Phomatospora, e.g., ref. [110]. In addition to genera typically associated with terrestrial palm samples and members of Tirisporellaceae and Annulatascaceae, a number of aquatic Sordariomycetes are commonly found colonising mangrove and peat swamp palms. These include the halosphaeriaceous genera Aniptodera, Fluviatispora, and Lignincola (Halosphaeriaceae, Microascales), Baipadisphaeria (Nectriaceae, Hypocreales), Flammispora (Sordariomycetes genus incertae sedis), Savoryella (Savoryellaceae, Savoryellales), Trichocladium (Chaetomiaceae, Sordariales), and Unisetosphaeria (Trichosphaeriaceae, Diaporthomycetidae family incertae sedis). Many other genera, whose phylogeny within the ascomycetes is still unresolved, are reported from freshwater and intertidal palm samples, such as Nypaella and Helicorhoidion (Ascomycota genera incertae sedis), e.g., refs. [166,700].
3.2.2. Palmicolous Dothideomycetes
Apart from the above-mentioned genera, most aquatic palmicolous fungi are representatives of the Dothideomycetes [596,686], including several members of Pleosporales, some of which also include terrestrial species, viz. Acuminatispora, Plectophomella (Pleosporales genera incertae sedis), Astrosphaeriella (Astrosphaeriellaceae), Carinispora (Pseudoastrosphaeriellaceae), Falciformispora, Trematosphaeria (Trematosphaeriaceae), Helicascus (Morosphaeriaceae), Herpotrichia (Melanommataceae), Leptosphaeria (Leptosphaeriaceae), Lolia (Lindgomycetaceae), Massarina (Massarinaceae), and Salsuginea (Salsugineaceae), e.g., refs. [116,166]. Recently, the new pleosporalean family Striatiguttulaceae was established to accommodate two new manglicolous fungi from palms, Longicorpus and Striatiguttula [9]. Moreover, other members of Dothideomycetes include aquatic representatives from palm samples, such as members in Jahnulales, including Jahnula (Aliquandostipitaceae) and Manglicola (Manglicolaceae).
In recent years, a series of Dothideomycetes have been frequently described from palm trees based on morpho-molecular data [687,688]. This, in turn, has clarified the structure of the taxa of Dothideomycetes that make up the assemblage of palm fungi and the best represented order is Pleosporales. Some of these taxa are part of the genera most frequently found on palm hosts, particularly the Astrosphaeriella-like species. The polyphyletic nature of Astrosphaeriella sensu lato has recently been resolved in different families and/or genera to include typically palmicolous taxa. Astrosphaeriella-like species on palms include different genera in three families, viz. Astrosphaeriella, Astrosphaeriellopsis, Pteridiospora, Pithomyces, Javaria, Xenoastrosphaeriella (Astrosphaeriellaceae), Fissuroma, Neoastrosphaeriella (Aigialaceae), and Pseudoastrosphaeriella (Pseudoastrosphaeriellaceae), e.g., refs. [267,533,701]. Moreover, members in Botryosphaeriales are also found on palms, including Barriopsis, Botryosphaeria, Diplodia, Lasiodiplodia, Neodeightonia (Botryosphaeriaceae), and Phyllosticta (Phyllostictaceae), and their potential as phytopathogens has occasionally been discussed, e.g., refs. [549,550,702,703]. Likewise, members of the Occultibambusaceae have recently been recorded in palms collections, including Brunneofusispora and Neooccultibambusa [537].
Several other Dothideomycetes are commonly found on palms and mostly reside in Pleosporales, especially in the Didymosphaeriaceae, such as Didymosphaeria, Montagnula, Paraconiothyrium, Paraphaeosphaeria, Pseudopithomyces, and Spegazzinia, and Roussoellaceae, such as Appendispora, Neoroussoella, and Roussoella, e.g., refs. [222,223,534]. Some of these genera are part of one of the most interesting groups of Dothideomycetes found on palms, as they include truly terrestrial ascomycetes with extracellular, often gelatinous appendages on the ascospores, which are mainly known from aquatic habitats, e.g., ref. [147]. However, many other pleosporalean fungi have been recorded on palms from a wide range of families. These include members of the Coniothyriaceae, such as Coniothyrium; Delitschiaceae, such as Delitschia; Lophiostomataceae, such as Lophiostoma and Vaginatispora; and Neophaeosphaeriaceae, such as Neophaeosphaeria, e.g., ref. [616]. Other members of Pleosporales found on palm include the genus Corynesporasca (Corynesporascaceae), which is a morphological genus introduced based on palm collections. Although it has been shown that Corynesporasca has a Corynespora-like asexual morph, the phylogenetic relationships of these two genera are unclear until molecular data of the type species are available [275].
3.2.3. Palmicolous “Anamorphs”
In addition to the previously mentioned anamorphic taxa, many families and members of Pleosporales that include asexually reproducing fungi, especially hyphomycetes, are typically found on palms. These include members of the Dictyosporiaceae, such as Dictyocheirospora, Dictyopalmispora, Dictyosporium, Pseudocoleophoma, and Sporidesmiella, e.g., ref. [602]; Hermatomycetaceae, including Hermatomyces; Leptosphaeriaceae, including Chaetoplea and Quasiphoma, e.g., refs. [569,573]; Massarinaceae, including Haplohelminthosporium, Helminthosporiella, and Helminthosporium, e.g., ref. [285]; Melanommataceae, including Asymmetricospora, Byssosphaeria, Camposporium, and Herpotrichia, e.g., refs. [575,704]; Periconiaceae, including Periconia, e.g., ref. [705]; Phaeosphaeriaceae, including Amarenographium, Parastagonospora, Phaeosphaeria, Septoriella, and Wojnowiciella, e.g., ref. [292]; Pleosporaceae, including Bipolaris, Curvularia, and Exserohilum, e.g., ref. [706]; Pseudoberkleasmiaceae, namely Pseudoberkleasmium, e.g., ref. [576]; Teichosporaceae, including Parateichospora, e.g., ref. [299]; and Tetraplosphaeriaceae, including Ernakulamia and Tetraploa, e.g., ref. [541]; as well as other taxa, for instance, the genus Repetophragma (Pleosporales genus incertae sedis). Moreover, species of Torula-like genera are also frequently recorded on palms from tropical countries, including Bahusandhika (Lentimurisporaceae), Cylindrotorula, and Torula (Torulaceae), e.g., ref. [587].
Studies on palmicolous “anamorphs” are scarce and are mostly restricted to dematiaceous hyphomycetes and botryosphaeriaceous coelomycetes, which seem to be an important assemblage of fungi that inhabit palm tissues, especially in the tropics. However, several palmicolous “anamorphs” have recently been described and introduced in mycological series as occasional discoveries. In turn, the taxonomic structure of palmicolous “anamorphs” is becoming better known and more complex, including many Sordariomycetes and particularly Dothideomycetes.
Concerning Sordariomycetes, the genera Ascotaiwania, Canalisporium (Savoryellaceae, Savoryellales), Distoseptispora (Distoseptisporaceae, Distoseptisporales), Monotosporella (Pleurotheciaceae, Pleurotheciales), Melanconis, Melanconium (Melanconidaceae, Diaporthales), Melanographium (Sordariomycetes genus incertae sedis), Pararamichloridium (Pararamichloridiaceae, Pararamichloridiales), and Spadicoides (Xenospadicoidaceae, Xenospadicoidales) are some of the hyphomycetes typically recorded on palms, e.g., refs. [15,238,612].
In addition, some of the most common Sordariomycetes taxa found on palms also include palmicolous “anamorphs”. These comprise several members of the Xylariales, including the genera Ascotricha, Diabolocovidia (Xylariaceae), Barrmaelia (Barrmaeliaceae), Circinotrichum, Gyrothrix (Xylariales genera incertae sedis), Endocalyx (Cainiaceae), Hansfordia (Hansfordiaceae), Microdochium (Microdochiaceae), and Zygosporium (Zygosporiaceae), e.g., refs. [546,562]. Likewise, several anamorphic fungi of the Chaetosphaeriales are recorded on palms, including Chloridium, Codinaea, Craspedodidymum, Cryptophiale, Dictyochaeta, Kionochaeta, Rattania, Sporoschisma, Thozetella (Chaetosphaeriaceae), and Endophragmiella (Helminthosphaeriaceae), e.g., refs. [243,707]. Moreover, a plethora of asexual Hypocreales genera have common representatives on palm hosts, such as Acremonium, Gossypinidium, Hydropisphaera, Lasionectriella, Paracylindrocarpon (Bionectriaceae), Chaetopsina, Dactylonectria, Pleiocarpon, Volutella (Nectriaceae), Neoacremonium (Neoacremoniaceae), Alfaria, Stachybotrys, and Virgatospora (Stachybotryaceae), e.g., refs. [283,317]. In addition, species of Fusarium (Nectriaceae) are also common on palm hosts and have been associated with important diseases, e.g., ref. [708]. Other Sordariomycetes orders with palmicolous asexual morphs include the Magnaporthales genus Pyricularia (Pyriculariaceae); the Microascales members Ceratocystis, Thielaviopsis (Ceratocystidaceae), Cirrenalia (Halosphaeriaceae), Custingophora (Gondwanamycetaceae), Triadelphia (Triadelphiaceae), and Wardomycopsis (Microascaceae); and many other genera, such as Diaporthe (Diaporthaceae, Diaporthales), Coniella (Schizoparmaceae, Diaporthales), Koorchaloma (Trichosphaeriaceae, Diaporthomycetidae family incertae sedis), Paraproliferophorum (Diaporthomycetidae genus incertae sedis), and Pararamichloridium (Pararamichloridiaceae, Pararamichloridiales), e.g., refs. [8,226].
Species of pestalotioid fungi in Amphisphaeriales have recently been described from palm collections, including some members of the Sporocadaceae, such as Bartalinia, Morinia, Neopestalotiopsis, Pestalotiopsis, Pseudopestalotiopsis, Robillarda, and Seiridium. Other “anamorphs” in Amphisphaeriales with representatives on palms include, for instance, Beltrania (Beltraniaceae), Arthrinium, and Dictyoarthrinium (Apiosporaceae), e.g., refs. [556,559].
Several other palmicolous “anamorphs” are Sordariomycetes, such as the hyphomycetes genera Acrodictys (Acrodictyaceae, Sordariomycetes family incertae sedis), Apogaeumannomyces (Sordariomycetes genus incertae sedis), and Hyalobelemnospora (Ophiostomataceae, Ophiostomatales). Even so, most of palmicolous “anamorphs” are Dothideomycetes or, like some of the above-mentioned Sordariomycetes, represent morphological, monotypic genera introduced based on palm collections and are known only from palms or almost exclusively from palms and their phylogeny is still uncertain and reside in Ascomycota genera incertae sedis. These include the hyphomycetes genera Acarocybellina, Acarocybiopsis, Agrabeeja, Anabahusakala, Ashtaangam, Atrosetaphiale, Basauxia, Bhadradriella, Botryomonilia, Brachysporiopsis, Bulbocatenospora, Ceratosporella, Cheiromyceopsis, Consetiella, Delortia, Dwibahubeeja, Endosporoideus, Helensiella, Hemisynnema, Holubovaea, Kalamarospora, Mackenziella, Nusia, Sawantomyces, Septosporiopsis, Setophiale, Spiculostilbella, Stratiphoromyces, Paradactylella, Phragmospathulella, Polybulbophiale, Rogergoosiella, Tretendophragmia, Tretocephala, Venustisporium, Venustocephala, Veramycella, Veramyces, and Waihonghopes (for references and details of some of these genera, see Table 1). The recollection of these taxa is imperative for the knowledge of the assemblage of palmicolous “anamorphs”. Other asexual morphs known from palms that reside in Ascomycota genera incertae sedis include Argopericonia, Barnettella, Bharatheeya, Bhatia, Capitorostrum, Ceratosporella, Drepanospora, Endomelanconium, Everhartia, Grallomyces, Goidanichiella, Haplobasidion, Helicoubisia, Kostermansinda, Lacellina, Lomachashaka, Lylea, Megalodochium, Phaeomonilia, Podosporium, Polytretophora, Pseudotorula, Sporidesmiopsis, Stauriella, Staurophoma, Tharoopama, and Vanakripa, e.g., refs. [229,236,517].
Numerous Dothideomycetes are recorded on palm trees, many of which include asexual morphs. In addition to some of the above-mentioned genera, such as Cirrenalia, Delortia, Drepanospora, Everhartia, Helicorhoidion, and Helicoubisia, many other allied genera of helicosporous hyphomycetes are found in association with palms, including Hymenoscyphus (Helotiaceae, Helotiales), Xenosporium (Dothideomycetes genus incertae sedis), and members of the Tubeufiaceae (Tubeufiales), such as Helicoma, Helicomyces, Helicosporium, and Thaxteriella, along with other non-helicosporous genera, such as Berkleasmium, e.g., refs. [43,319,321,324]. Other members in Tubeufiales encountered on palms include Aquaphila (Tubeufaceae) and Wiesneriomyces (Wiesneriomycetaceae). In Asterinales, species of Asterina, Cirsosia, Discopycnothyrium (Asterinaceae), Lembosia (Lembosiaceae), and Morenoina (Morenoinaceae) were collected from palms, some of which included known pathogens commonly represented on palms, e.g., refs. [152,279,606]. Several palmicolous “anamorphs”, particularly hyphomycetes, with Mycosphaerella-like sexual morphs reside in Mycosphaerellales, including some species of Cercospora, Distocercospora, Exosporium, Pallidocercospora, Passalora, Phaeophleospora, Pseudocercospora, Ramularia, Scolecostigmina, Uwemyces, Zasmidium (Mycosphaerellaceae), and Pseudoepicoccum (Mycosphaerellales genus incertae sedis), some of which have been recorded on palms associated with foliar diseases, e.g., refs. [630,709,710,711]. Recently Palmeiromyces (Teratosphaeriaceae) was recorded as an obligate biotroph causing palm leafspots [295]. Other members of Mycosphaerellales, especially dematiaceous hyphomycetes, have been recorded on palms, such as Castanedospora (Extremaceae) and Stenella (Teratosphaeriaceae), e.g., ref. [273]. Moreover, in Kirschsteiniotheliales, the hyphomycetes genera Kirschsteiniothelia (Kirschsteiniotheliaceae) and Taeniolella (Kirschsteiniotheliales genus incertae sedis) have been recorded on palms, e.g., ref. [618].
Table 2.
Synopsis of the taxonomic structure of palm fungi: genera and respective families in subclasses of Sordariomycetes and Dothideomycetes with common representatives found on Arecaceae hosts.
Table 2.
Synopsis of the taxonomic structure of palm fungi: genera and respective families in subclasses of Sordariomycetes and Dothideomycetes with common representatives found on Arecaceae hosts.
| Class | Subclass | Order | Family | Genera |
|---|---|---|---|---|
| Dothideomycetes | Dothideomycetidae | Dothideales | Dothideaceae | Uleodothis |
| Mycosphaerellales | Extremaceae | Castanedospora | ||
| Mycosphaerellaceae | Cercospora, Distocercospora, Exosporium, Pallidocercospora, Passalora, Phaeophleospora, Pseudocercospora, Ramularia, Scolecostigmina, Uwemyces, Zasmidium | |||
| Teratosphaeriaceae | Palmeiromyces, Stenella | |||
| Incertae sedis | Pseudoepicoccum | |||
| Pleosporomycetidae | Acrospermales | Acrospermaceae | Gonatophragmium | |
| Pleosporales | Acrocalymmaceae | Acrocalymma | ||
| Aigialaceae | Fissuroma, Neoastrosphaeriella | |||
| Arthopyreniaceae | Mycomicrothelia | |||
| Astrosphaeriellaceae | Astrosphaeriella, Astrosphaeriellopsis, Pteridiospora, Pithomyces, Javaria, Triseptatospora, Xenoastrosphaeriella | |||
| Coniothyriaceae | Coniothyrium | |||
| Corynesporascaceae | Corynesporasca | |||
| Delitschiaceae | Delitschia | |||
| Dictyosporiaceae | Dictyocheirospora, Dictyopalmispora, Dictyosporium, Pseudocoleophoma, Sporidesmiella | |||
| Didymosphaeriaceae | Didymosphaeria, Montagnula, Paraconiothyrium, Paraphaeosphaeria, Pseudopithomyces | |||
| Hermatomycetaceae | Hermatomyces | |||
| Lentimurisporaceae | Bahusandhika | |||
| Leptosphaeriaceae | Chaetoplea, Leptosphaeria, Quasiphoma | |||
| Lindgomycetaceae | Lolia | |||
| Dothideomycetes (cont.) | Dothideomycetidae (cont.) | Pleosporales (cont.) | Lophiostomataceae | Lophiostoma, Vaginatispora |
| Massarinaceae | Haplohelminthosporium, Helminthosporiella, Helminthosporium, Massarina | |||
| Melanommataceae | Asymmetricospora, Byssosphaeria, Camposporium, Herpotrichia | |||
| Morosphaeriaceae | Helicascus | |||
| Neophaeosphaeriaceae | Neophaeosphaeria | |||
| Occultibambusaceae | Brunneofusispora, Neooccultibambusa | |||
| Periconiaceae | Periconia | |||
| Phaeosphaeriaceae | Amarenographium, Parastagonospora, Phaeosphaeria, Septoriella, Wojnowiciella | |||
| Pleosporaceae | Bipolaris, Curvularia, Exserohilum | |||
| Pseudoastrosphaeriellaceae | Carinispora, Pseudoastrosphaeriella | |||
| Pseudoberkleasmiaceae | Pseudoberkleasmium | |||
| Roussoellaceae | Appendispora, Neoroussoella, Roussoella | |||
| Salsugineaceae | Salsuginea | |||
| Striatiguttulaceae | Longicorpus, Striatiguttula | |||
| Trematosphaeriaceae | Falciformispora, Trematosphaeria | |||
| Teichosporaceae | Parateichospora | |||
| Tetraplosphaeriaceae | Ernakulamia, Tetraploa | |||
| Torulaceae | Cylindrotorula, Torula | |||
| Incertae sedis | Acuminatispora, Plectophomella, Repetophragma | |||
| Hysteriales | Hysteriaceae | Gloniopsis | ||
| Incertae sedis | Asterinales | Asterinaceae | Asterina, Cirsosia, Discopycnothyrium | |
| Lembosiaceae | Lembosia | |||
| Morenoinaceae | Morenoina | |||
| Botryosphaeriales | Botryosphaeriaceae | Barriopsis, Botryosphaeria, Diplodia, Lasiodiplodia, Neodeightonia | ||
| Dothideomycetes (cont.) | Incertae sedis (cont.) | Botryosphaeriales (cont.) | Phyllostictaceae | Phyllosticta |
| Jahnulales | Aliquandostipitaceae | Jahnula | ||
| Manglicolaceae | Manglicola | |||
| Kirschsteiniotheliales | Kirschsteiniotheliaceae | Kirschsteiniothelia | ||
| Incertae sedis | Taeniolella | |||
| Muyocopronales | Muyocopronaceae | Muyocopron, Pseudopalawania | ||
| Tubeufiales | Tubeufiaceae | Aquaphila, Berkleasmium, Helicoma, Helicomyces, Helicosporium, Thaxteriella | ||
| Wiesneriomycetaceae | Wiesneriomyces | |||
| - | Palawaniaceae | Palawania | ||
| - | Trichopeltinaceae | Acrogenotheca | ||
| - | - | Letendraeopsis, Xenosporium, Brooksia, Dianesea, Leptomeliola, Scolionema | ||
| Sordariomycetes | Diaporthomycetidae | Annulatascales | Annulatascaceae | Annulatascus, Submersisphaeria |
| Diaporthales | Diaporthaceae | Diaporthe | ||
| Gnomoniaceae | Maculatipalma | |||
| Melanconidaceae | Melanconis, Melanconium | |||
| Schizoparmaceae | Coniella | |||
| Incertae sedis | Durispora, Phruensis | |||
| Distoseptisporales | Distoseptisporaceae | Distoseptispora | ||
| Magnaporthales | Magnaporthaceae | Gaeumannomyces | ||
| Ophioceraceae | Ophioceras | |||
| Pseudohalonectriaceae | Pseudohalonectria | |||
| Ophiostomatales | Ophiostomataceae | Hyalobelemnospora | ||
| Phomatosporales | Phomatosporaceae | Phomatospora | ||
| Tirisporellales | Tirisporellaceae | Bacusphaeria, Thailandiomyces, Tirisporella | ||
| Sordariomycetes (cont.) | Diaporthomycetidae (cont.) | Xenospadicoidales | Xenospadicoidaceae | Koorchaloma, Spadicoides |
| Incertae sedis | Mesnieraceae | Bondiella | ||
| Trichosphaeriaceae | Unisetosphaeria | |||
| Thyridiaceae | Thyridium | |||
| - | Paraproliferophorum | |||
| Hypocreomycetidae | Glomerellales | Plectosphaerellaceae | Acremoniisimulans, Brunneomyces | |
| Hypocreales | Bionectriaceae | Acremonium, Clonostachys, Gossypinidium, Hydropisphaera, Ijuhya, Lasionectria, Nectriella, Paracylindrocarpon | ||
| Hypocreaceae | Verticimonosporium | |||
| Nectriaceae | Baipadisphaeria, Calonectria, Chaetopsina, Cosmospora, Dactylonectria, Fusarium, Ilyonectria, Nectria, Nectriopsis, Ophionectria, Pleiocarpon, Volutella | |||
| Neoacremoniaceae | Neoacremonium | |||
| Niessliaceae | Niesslia | |||
| Stachybotryaceae | Alfaria, Stachybotrys, Virgatospora | |||
| Microascales | Gondwanamycetaceae | Custingophora | ||
| Halosphaeriaceae | Aniptodera, Cirrenalia, Fluviatispora, Lignincola | |||
| Microascaceae | Wardomycopsis | |||
| Triadelphiaceae | Triadelphia | |||
| Ceratocystidaceae | Ceratocystis, Thielaviopsis | |||
| Pararamichloridiales | Pararamichloridiaceae | Pararamichloridium | ||
| Savoryellomycetidae | Conioscyphales | Conioscyphaceae | Conioscypha | |
| Pleurotheciales | Pleurotheciaceae | Monotosporella | ||
| Savoryellales | Savoryellaceae | Ascotaiwania, Canalisporium, Savoryella | ||
| Sordariomycetidae | Chaetosphaeriales | Chaetosphaeriaceae | Chaetosphaeria, Chloridium, Codinaea, Craspedodidymum, Cryptophiale, Dictyochaeta, Kionochaeta, Rattania, Sporoschisma, Thozetella | |
| Sordariomycetes (cont.) | Sordariomycetidae (cont.) | Chaetosphaeriales (cont.) | Helminthosphaeriaceae | Endophragmiella |
| Leptosporellaceae | Leptosporella | |||
| Linocarpaceae | Linocarpon, Neolinocarpon | |||
| Incertae sedis | Caudatispora | |||
| Coniochaetales | Incertae sedis | Cannonia | ||
| Meliolalles | Meliolaceae | Meliola | ||
| Phyllachorales | Phaeochoraceae | Cocoicola, Phaeochora, Phaeochoropsis, Serenomyces | ||
| Phyllachoraceae | Brobdingnagia, Camarotella, Coccodiella, Coccostromopsis, Maculatifrondes, Malthomyces, Ophiodothella, Oxodeora, Phyllachora, Sphaerodothis, Tribulatia | |||
| Sordariales | Chaetomiaceae | Trichocladium | ||
| Lasiosphaeriaceae | Cercophora, Lasiosphaeria | |||
| Incertae sedis | Lockerbia | |||
| Incertae sedis | - | Arecacicola, Curvatispora, Nigromammilla, Paracapsulospora | ||
| Xylariomycetidae | Amphisphaeriales | Amphisphaeriaceae | Amphisphaeria, Lepteutypa | |
| Apiosporaceae | Arthrinium, Dictyoarthrinium | |||
| Appendicosporaceae | Appendicospora | |||
| Beltraniaceae | Beltrania | |||
| Hyponectriaceae | Arecomyces, Frondicola, Hyponectria, Rachidicola | |||
| Iodosphaeriaceae | Iodosphaeria | |||
| Oxydothidaceae | Oxydothis | |||
| Pseudomassariaceae | Leiosphaerella, Pseudomassaria | |||
| Sporocadaceae | Bartalinia, Morinia, Neopestalotiopsis, Pestalotiopsis, Pseudopestalotiopsis, Robillarda, Seiridium | |||
| Xylariales | Barrmaeliaceae | Barrmaelia | ||
| Sordariomycetes (cont.) | Xylariomycetidae (cont.) | Xylariales (cont.) | Cainiaceae | Arecophila, Seynesia, Endocalyx |
| Clypeosphaeriaceae | Apioclypea, Brunneiapiospora, Palmaria | |||
| Diatrypaceae | Allocryptovalsa, Allodiatrype, Anthostoma, Cryptovalsa, Diatrype, Diatrypella, Eutypa, Eutypella, Frondisphaeria, Peroneutypa | |||
| Fasciatisporaceae | Fasciatispora | |||
| Graphostromataceae | Biscogniauxia | |||
| Hansfordiaceae | Hansfordia | |||
| Hypoxylaceae | Annulohypoxylon, Hypoxylon | |||
| Microdochiaceae | Idriella, Microdochium | |||
| Oxydothidaceae | Oxydothis | |||
| Robillardaceae | Robillarda | |||
| Xylariaceae | Anthostomella, Ascotricha, Astrocystis, Diabolocovidia, Kretzschmaria, Nemania, Neoxylaria, Rosellinia, Stilbohypoxylon, Xylaria | |||
| Zygosporiaceae | Zygosporium | |||
| Incertae sedis | Capsulospora, Circinotrichum, Cyanopulvis, Gyrothrix, Guestia, Haploanthostomella, Lasiobertia, Neobarrmaelia, Nipicola, Palmicola, Pemphidium, Pulmosphaeria, Sabalicola | |||
| Incertae sedis | Myelospermataceae | Myelosperma | ||
| - | Frondispora, Manokwaria | |||
| Incertae sedis | Catabotryales | Catabotryaceae | Catabotrys | |
| - | Acrodictyaceae | Acrodictys | ||
| - | - | Apogaeumannomyces, Flammispora, Mangrovispora |
3.2.4. Miscellaneous Palm Taxa
Several other Dothideomycetes are reported from palms and their taxonomy is either incertae sedis or spread over a plethora of orders and families, unlike the palmicolous Sordariomycetes, whose taxonomy, although highly diverse, seems to be more concentrated in some specific orders. These Dothideomycetes include, for example, Acrogenotheca (Trichopeltinaceae, Dothideomycetes family incertae sedis), Bondiella (Mesnieraceae, Dothideomycetes family incertae sedis), Brooksia, Dianesea, Leptomeliola, Scolionema (Dothideomycetes genera incertae sedis), Gonatophragmium (Acrospermaceae, Acrospermales), Gloniopsis (Hysteriaceae, Hysteriales), Letendraeopsis (Dothideomycetes genus incertae sedis), Muyocopron, Pseudopalawania (Muyocopronaceae, Muyocopronales), Mycomicrothelia (Arthopyreniaceae, Pleosporales), Palawania (Palawaniaceae, Dothideomycetes family incertae sedis), Uleodothis (Dothideaceae, Dothideales), and many other taxa occasionally reported, some of which are morphological genera only known from palms, e.g., refs. [303,536,620].
Although most palm fungi belong to the Dothideomycetes and Sordariomycetes, a series of taxa from other classes of Ascomycota are also often encountered on palm collections, including, for example, the genera Mazosia (Roccellaceae, Arthoniales, Arthoniomycetes), Morchella (Morchellaceae, Pezizales, Pezizomycetes), Stictis (Stictidaceae, Ostropales, Lecanoromycetes), and various Leotiomycetes taxa. These include members of Helotiales, such as, Diplococcium (Vibrisseaceae), Hymenoscyphus (Helotiaceae), Phialocephala (Mollisiaceae), Porodiplodia (Porodiplodiaceae), many species of Lachnum and Lachnellula (Lachnaceae), the aquatic genus Vibrissea, Strossmayeria (Vibrisseaceae), and some genera incertae sedis, viz. Cenangiumella, Sorokina, and Sorokinella. Fungal members of other Leotiomycetes orders are also represented on palms, such as Chalara (Pezizellaceae, Rhytismatales), Dactylaria (Calloriaceae, Rhytismatales), Lophodermium (Rhytismataceae, Rhytismatales), Phacidium (Phacidiaceae, Phacidiales), and Phlyctema (Dermateaceae, Medeolariales), e.g., refs. [240,241,622,712,713]. However, considering the comprehensive review of literature carried out here, the great diversity of palmicolous ascomycetes is basically restricted to the subphylum Pezizomycotina.
Very few basidiomycetes have been reported from palms and, as a result, their knowledge is practically unknown. However, some palmicolous basidiomycetes are worth mentioning due to their recurrent or specific association with palms. These include the genus Ganoderma (Ganodermataceae, Polyporales, Agaricomycetes, Agaricomycotina), which is an important phytopathogen that rots the lower part of palm trunks, e.g., ref. [714], and the genus Graphiola (Graphiolaceae, Exobasidiales, Exobasidiomycetes, Ustilaginomycotina), which parasites almost exclusively Arecaceae hosts, causing leaf spots on wide range of palm species, e.g., ref. [715]. Several other genera of Basidiomycota have occasionally been isolated from palms and studies on the oil palm basidiomycete assemblage have shown that members of Polyporales, such as Fomitopsis (Fomitopsidaceae), Pycnoporus, and Trametes (Polyporaceae), and Agaricales (Agaricomycetidae, Agaricomycetes, Agaricomycotina), such as Schizophyllum (Schizophyllaceae), may be some of the common basidiomycetes that inhabit palm substrata [523,643]. Nonetheless, more studies are needed to gain knowledge about the common basidiomycete assemblage of palm hosts.
4. Palm Trees as Model Plants for the Study of Fungal Biodiversity
Biodiversity is the variety of life on Earth and, consequently, depends on both taxa and their biotic and abiotic interactions [716,717]. Thus, ecological and biodiversity studies are extremely important, along with systematic and taxonomic approaches, to assess a complete and integrated perspective of the complex assemblages that make up biological communities and their ecosystems. Ecological and systematic research on palm fungi indicates that they are remarkably diverse and complex biological communities that exhibit a variety of different lifestyles. Therefore, the great diversity of palm fungi plays an important role in different aspects related to biodiversity and makes it possible to address several questions of great importance in biodiversity surveys.
Many studies have described palms as important substrata for exploring fungal diversity, particularly due to their intense fungal colonisation, e.g., refs. [7,10,12,352]. In addition, the close association and intrinsic relationship between palm hosts and palm fungi have occasionally been discussed, e.g., refs. [12,329,352,718]. Furthermore, the high fungal diversity recorded on Arecaceae hosts seems to be related to specific ecological and biodiversity issues observed in palm fungal communities. These include any kind of host- and tissue-specificity, or any kind of established biotic or abiotic relationships, e.g., refs. [12,13,14,329,634,718]. Here, palm trees are regarded as model plants for the study of fungal biodiversity and, therefore, the key role of palm fungi in biodiversity surveys is discussed.
4.1. Palm Fungi and the Search for the “Missing Fungi”
Perhaps one of the main aspects for which research on palm fungi is acknowledged is the fact that palm trees seem to harbour numerous undescribed microfungi, e.g., refs. [9,532]. To search for the undescribed mycota around the world is currently one of the main objectives of mycologists, who try to fill the gap between the number of fungal species currently described and the number of species that the most recent estimates predict. In addition, studies on fungal diversity are fundamental to increase awareness of the critical role of fungi in ecosystems. Therefore, to determine the magnitude and patterns of fungal diversity is an ongoing challenge in fungal biodiversity surveys [719,720,721].
The regular discovery of new fungal species has prompted mycologists to wonder about the number of fungi that exists worldwide. Since Fries [722], who established a comparison between the diversity of fungi and that of insects, fungi are known as one of the most speciose groups of organisms. Estimates of the number of fungal species worldwide has varied over time, ranging from relatively low numbers of 100,000 [723], 250,000 to 270,000 [724], and 712,000 [720] to impressive higher estimates of 3.5 to 5.1 [725] and almost 10 [726] and 12 million [721]. Until recently, the most widely cited and recognised number was the 1.5 million fungal species hypothesised by Hawksworth [727]. Hawksworth based his conclusions on observed ratios between fungal and plant species diversity in regions where fungi were considered to be well studied. However, even Hawksworth [727] considered this figure to be a conservative estimate. Moreover, it has been revisited several times in the literature as the worldwide description rate of new fungal species has increased over the last decades [10,728,729,730,731,732].
While molecular data is becoming the standard approach for identifying most fungal groups, environmental metabarcoding via high-throughput sequencing (HTS) is increasing the number of sequence data documenting fungal diversity worldwide [721,733,734,735,736,737,738]. Thus, predicting the number of fungal species worldwide has taken on a new dimension and divergent numbers and opinions are continuously expressed [739,740,741]. One of the ongoing debates among taxonomists is how to formally describe the so called “dark taxa”, i.e., lineages represented only by sequence data and for which no individual voucher specimens or cultures exist [589,742,743,744].
Although there is no universal approach to identifying fungi and accurately predicting fungal diversity on Earth, the currently accepted estimate of species richness is between 2.2 and 3.8 million [745]. This estimate was based on different datasets, including publication rates of new taxa, species recognition studies, extrapolations of ratios between plants and fungi, and consideration of “dark taxa” known as molecular sequence data from environmental samples [589,745]. Considering that around 150,000 accepted fungal species are currently known [746,747], these figures indicate that less than 10% of the world’s mycota have been named so far. Thus, more than 90% of all fungal species remain to be discovered. Taking into account that new species are reported at an average rate of 1500 to 2000 species per year [746,747], it could take more than 2000 years before all the missing fungi are discovered and named. More positive scenarios have been recently published, following updates to the number of named fungal species and the average rate at which fungal species are being published. Yet, the undescribed mycota will only be known in about 200 to 1800 years [746].
For all the above-mentioned reasons, the question “where are the missing fungi?” has often been asked and therefore locating and describing these taxa is a major task among mycologists [10,718,719,748,749,750]. Previous studies have highlighted that understudied biodiversity hotspots, less studied habitats and life modes, as well as less studied or fungi-rich and geographically widespread host plants (and their families), should be explored and may contain many of the missing taxa [595]. In fact, the inventory of fungal species from different substrata, especially those that seem to support a high species richness, is undoubtedly responsible for describing some of the missing fungal diversity [590].
Along with other host plants, such as bamboos (Poaceae) [751], Clematis (Ranunculaceae) [752], Eucalyptus (Myrtaceae) [753], Musa (Musaceae) [752], Pandanus (Pandanaceae) [754], and Rosa (Rosaceae) [755], Arecaceae hosts have been shown to be hyperdiverse substrata for fungal diversity, as well as a rich source of new fungal taxa. In the last three decades, a remarkable number of new taxa have been described from Arecaceae, e.g., refs. [6,8,9,10,17,104]). In fact, Hyde et al. [17] reported that 75% of all fungi collected on palms were new to science. In this sense, the evidence gained from the extensive palm fungi research undoubtedly indicates that many of the missing fungi can be found on palms. However, the extent of this assumption is yet to be determined and only predictions can be made.
Most of the taxa introduced from palm substrata over the last 30 years have been based solely on morphological diagnosis (Table 1). Thus, the identity of these fungi within a natural taxonomic framework has yet to be resolved. This is well illustrated when accessing Anthostomella-like species from palms. More than 30 species of Anthostomella have been introduced from palm collections, insomuch that around 60 palm species are known to be hosts of Anthostomella and allied genera [133,203,209,218,219,220,221,542]. However, none of the Anthostomella species described from palm trees have been re-evaluated in terms of their phylogenetics. Considering that recent studies have described this genus as polyphyletic and that several new taxa were hidden under Anthostomella-like species [539,544,545], the morphological species of Anthostomella described from palms may reveal a much greater diversity than initially observed. In addition, several morphological species are known to be cryptic species, i.e., distinct species that are misidentified and hidden under one species name [590,595,756]. This consequently increases the likelihood that several Anthostomella morphological species described from palms are unknown and misidentified taxa. Likewise, several other taxa described from palm trees have been based on morphology, which includes many of the taxa that are only known from Arecaceae hosts. In the last decade, many studies that used polyphasic approaches, i.e., morpho-phylogenetic analyses, have revealed numerous new taxa from widely studied regions, habitats, and hosts, e.g., ref. [757]. Recent studies have revisited the identification of palmicolous taxa and many new species are now being introduced based on morphological and sequence data, while others are being redefined according to modern DNA sequence-based classifications, e.g., refs. [522,530,531,532,533,541,607]. In turn, this has broadened and structured knowledge about the taxonomy of palm fungi (Table 2) (see Section 3). Thus, collections of palm fungi can help not only to record old taxa that need to be recollected and placed in a natural taxonomic framework, but also, undoubtedly, to reveal some of the world’s undescribed mycota.
In addition to being recognised as fungi-rich host plants, palm trees are also a geographically widespread and highly diverse group of plants. They comprise around 2600 species in 181 genera [1]. However, only a small number of palm species have been investigated for their associated microfungi. It has long been recognised that many more fungal species are reported from plants of economic importance than ones that are not recognised as of human interest [758]. This is also the case with palm trees. There is a disparity in knowledge about fungi on economically important palms compared with other palms. Thus, while economically important palm genera such as Cocos, Elaeis, Phoenix, and Calamus have a reported fungal richness ranging from around 650 to 1300 records, with an average value of ca. 1100 records, most palm genera have a reported fungal richness ranging from less than 10 to less than 300 records [759] (Table 3). This was well illustrated by Taylor and Hyde [8], who observed a pattern of relative species richness between different palm species—151 species for Archontophoenix alexandrae, 144 for Trachycarpus fortunei, and 77 for Cocos nucifera—which differed from the impression gained from species previously described as new taxa from these hosts—177 from C. nucifera, 12 from A. alexandrae, and 5 from T. fortunei. Given that palm trees are host plants rich in fungi, it seems obvious, looking at these figures, that there is a large amount of unstudied data on most palm species. Hence, although palm fungi have been investigated to some extent, their knowledge is still underexplored and the fungal composition for most palm species is virtually unknown.
By accessing the collection of palm fungi, especially through the reporting of taxonomic novelties, it is possible to see that more than 260 palm species in more than 95 genera have been studied for their associated microfungi [759] (Table 3). However, the majority represent occasional collections in the field, insomuch that the great majority of palm species have a reported fungal richness ranging from less than 20 to less than 50 (Table 3). However, some have been chosen for biodiversity studies because they represent palms of particular interest and/or palms that inhabit ecosystems of particular interest. Some examples are considered herein.
Table 3.
Synopsis of fungal records on Arecaceae hosts retrieved from the U.S. National Fungus Collections Fungus–Host Database [759]. Palm species with less than 100 associated fungal records were disregarded from the detailed list but were considered in the summary figures.
Table 3.
Synopsis of fungal records on Arecaceae hosts retrieved from the U.S. National Fungus Collections Fungus–Host Database [759]. Palm species with less than 100 associated fungal records were disregarded from the detailed list but were considered in the summary figures.
| Palm Species 1 | Total Number of | |||||||
|---|---|---|---|---|---|---|---|---|
| Fungal Records 2 | Fungal Species 2,3 | Ascomycetes | Asexual Morphs | Coelomycetes | Hyphomycetes | Basidiomycetes | Zygomycetes | |
| Cocos nucifera | 1296 | 526 | 149 (28.33%) | 275 (52.28%) | 91 (17.30%) | 184 (34.98%) | 96 (18.25%) | 6 (1.14%) |
| Elaeis guineensis | 1256 | 427 | 100 (23.42%) | 235 (55.04%) | 50 (11.71%) | 185 (43.33%) | 80 (18.74%) | 12 (2.81%) |
| Phoenix dactylifera | 560 | 197 | 48 (24.37%) | 123 (62.44%) | 39 (19.80%) | 84 (42.64%) | 23 (11.68%) | 3 (1.52%) |
| Archontophoenix alexandrae | 355 | 178 | 87 (48.88%) | 86 (48.31%) | 11 (6.18%) | 75 (42.13%) | 5 (2.81%) | 0 |
| Areca catechu | 298 | 155 | 26 (16.77%) | 111 (71.61%) | 33 (21.29%) | 78 (50.32%) | 16 (10.32%) | 2 (1.29%) |
| Trachycarpus fortunei | 297 | 154 | 58 (37.66%) | 94 (61.04%) | 41 (26.62%) | 53 (34.42%) | 2 (1.30%) | 0 |
| Roystonea regia | 225 | 153 | 19 (12.41%) | 123 (80.39%) | 16 (10.46%) | 107 (69.93%) | 11 (7.19%) | 0 |
| Livistona chinensis | 189 | 95 | 47 (49.47%) | 35 (36.84%) | 10 (10.53%) | 25 (26.32%) | 13 (13.68%) | 0 |
| Phoenix loureiroi | 173 | 92 | 27 (29.35%) | 63 (68.48%) | 9 (9.78%) | 54 (58.70%) | 2 (2.17%) | 0 |
| Phoenix canariensis | 160 | 91 | 24 (26.37%) | 43 (47.25%) | 12 (13.19%) | 31 (34.07%) | 24 (26.37%) | 0 |
| Chamaerops humilis | 128 | 64 | 32 (50.00%) | 22 (34.38%) | 12 (18.75%) | 10 (15.63%) | 10 (15.63%) | 0 |
| Sabal palmetto | 128 | 88 | 45 (51.14%) | 28 (31.82%) | 6 (6.82%) | 22 (25.00%) | 15 (17.05%) | 0 |
| Arenga engleri | 122 | 64 | 14 (21.88%) | 50 (78.13%) | 4 (6.25%) | 46 (71.88%) | 0 | 0 |
| Licuala longicalycata | 119 | 89 | 49 (55.06%) | 40 (44.94%) | 3 (3.37%) | 37 (41.57%) | 0 | 0 |
| Rhopalostylis sapida | 113 | 88 | 36 (40.91%) | 41 (46.59%) | 0 | 41 (46.59%) | 11 (12.50%) | 0 |
| Summary figures 1,2,3 | Total number of palm genera from which associated fungi have been studied: 97 | |||||||
| Palm genera with a total number of fungal records ≥ 100: Cocos (1296 fungal records), Elaeis (1286), Phoenix (1146), Calamus (658), Archontophoenix (395), Areca (333), Rhopalostylis (318), Trachycarpus (306), Livistona (278), Sabal (274), Roystonea (270), Licuala (244), Arenga (229), Caryota (176), Chamaerops (128), Syagrus (112), Chamaedorea (108), and Borassus (105) | ||||||||
| Total number of palm species from which associated fungi have been studied: 262 | ||||||||
| Total number of palm species with a total number of fungal records ≥ 100: 15 | ||||||||
| Total number of palm species with 100 < total number of fungal records ≥ 50: 12 | ||||||||
| Total number of palm species with 50 < total number of fungal records ≥ 20: 26 | ||||||||
| Total number of palm species with a total number of fungal records < 20: 209 | ||||||||
| Total number of fungal records associated with Arecaceae hosts: 9339 | ||||||||
| Total number of fungal species recorded from Arecaceae hosts: 2932, including 1182 ascomycetes (40.31%), 332 basidiomycetes (11.32%), 1398 anamorphic fungi (47.68%), namely 984 ascomycetes (33.56%) and 413 coelomycetes (14.09%), and 20 zygomycetes (0.68%) | ||||||||
1 All palm species names annotated in the US National Fungus Collections Fungus–Host Database were checked against the Plants of the World Online Database [760]. The fungal records that were reported from palm species identified only to genus or from unidentified Arecaceae hosts were only counted in the summary figures, regardless of whether their total number was more than 100. 2 For the total number of records and species, only the taxa of the Fungi kingdom were considered, so the records of Oomycota and Myxomycota associated with Arecaceae hosts annotated in the US National Fungus Collections Fungus–Host Database were excluded. The total number of fungal records includes records of taxa identified only to genus, as well as records of the same taxa that were obtained from different studies. 3 The total number of fungal species excludes taxa identified only to genus, as well as taxa that have been annotated more than once. The latter may correspond either to different collections of the same taxa on a certain Arecaceae host, to collections of the same taxa from different Arecaceae hosts, or to the same collection reported in different studies. Note: the palm species are listed in descending order of the total number of associated fungal records and species. The fungal species annotated in the US National Fungus Collections Fungus–Host Database and used to construct the table have not been verified in official nomenclatural repositories, so the current classification of some taxa is likely to be different. Taxa currently synonymised under other taxa and taxa for which “teleomorph-anamorph” connections have been established are likely to be overestimating the figures presented. This is likely to be the reason why the summary figure presented for the total number of fungal species recorded from Arecaceae hosts is substantially different from the global figure of palm fungi estimated in the present review (see Section 3.1).
The fungi colonising peat swamp palms have been relatively well documented, as these palms inhabit unique ecosystems comprising very distinct abiotic conditions, which are extremely important worldwide due to their rich biodiversity. The numbers of fungi and novel taxa recorded on collections of palms in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand, are presented in Table 4 and Table 5, respectively.
Table 4.
Numbers of fungi recorded on collections of palms in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand.
Table 4.
Numbers of fungi recorded on collections of palms in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand.
| Palm Species | Total Number of | Reference | ||||
|---|---|---|---|---|---|---|
| Fungal Records | Fungal Species | Ascomycetes | Asexual Morphs | Basidiomycetes | ||
| Eleiodoxa conferta | 462 | 112 | 43 (38%) | 67 (60%) | 2 (2%) | [248] |
| Licuala longicalycata | 358 | 147 | 79 (53%) | 65 (45%) | 3 (3%) | [249] |
| Metroxylon sagu | 82 | 45 | 21 (47%) | 24 (53%) | 0 | [669] |
| Nenga pumila | 184 | 47 | 19 (40%) | 28 (60%) | 0 | [669] |
Pinnoi et al. [248] reported a total of 462 fungal records following six field collections of Eleiodoxa conferta, including 112 fungal species, among which 9 new species and 1 new genus were introduced (Table 4 and Table 5). Pinruan et al. [249] reported a total of 358 fungal records following 6 field collections of Licuala longicalycata, including 147 fungal species, among which 9 new species and 4 new genera were introduced (Table 4 and Table 5).
Table 5.
Novel taxa described from palm substrata collected in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand.
Table 5.
Novel taxa described from palm substrata collected in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand.
| Genus | Species | Substratum | Reference |
|---|---|---|---|
| Astrocystis eleiodoxae | On a submerged petiole of Eleiodoxa conferta | [525] | |
| Baipadisphaeria | Baipadisphaeria spathulospora | On a submerged trunk of Licuala longicalycata | [270] |
| Chalara siamensis (as C. siamense) | On submerged dead petioles of E. conferta | [241] | |
| Craspedodidymum licualae | On a decaying trunk of L. longicalycata | [243] | |
| Cras. microsporum | On a decaying trunk of L. longicalycata | [243] | |
| Cras. siamense | On a decaying sheath of L. longicalycata | [243] | |
| Dactylaria flammulicornuta | On a terrestrial petiole of Nenga pumila | [245] | |
| D. palmae | On terrestrial sheath of N. pumila | [245] | |
| D. uliginicola | On a submerged rachis of E. conferta | [245] | |
| Dictyopalmispora | Dictyopalmispora palmae | On decaying leaves of L. longicalycata | [602] |
| Flammispora | Flammispora bioteca | On submerged decaying leaves of L. longicalycata | [282] |
| Goidanichiella fusiformis (as G. fusiforma) | On a submerged dead petiole of E. conferta | [236] | |
| Jahnula appendiculata | On a submerged trunk of L. longicalycata | [242] | |
| Knoxdaviesia undulatistipes (as Custingophora undulatistipes) | On a submerged dead petiole of E. conferta | [246] | |
| Phruensis | Phruensis brunneispora | On a dead trunk of L. longicalycata | [301] |
| Stachybotrys palmae | On a decaying rachis of L. longicalycata | [244] | |
| Submersisphaeria palmae | On submerged petioles, rachides, and trunks of E. conferta, N. pumila and L. longicalycata | [247] | |
| Thailandiomyces | Thailandiomyces bisetulosus | On submerged senescent trunk of L. longicalycata | [307] |
| Unisetosphaeria | Unisetosphaeria penguinoides | On a submerged dead petiole of E. conferta | [245] |
| Vanakripa minutiellipsoidea | On a submerged dead petiole of E. conferta | [246] |
Similarly, fungi from mangrove palms, especially Nypa fruticans, have been widely investigated. Mangrove forests are highly specialised habitats adapted to extreme salinity conditions, which play an important role in the ecology of tropical and subtropical coastal waters. A total of 142 taxa were reported from collections of N. fruticans, among which 45 new species and 11 new genera were introduced [9,261,535,597,674,761] (Table 6). Although many genera and species of fungi have been well documented from N. fruticans and some peat swamp palms, very few molecular studies of fungi associated with these palms have been carried out (see Section 2). Thus, molecular analysis will certainly be able to provide the mycota that have not yet been discovered in the fungal communities that occur on these Arecaceae hosts.
Table 6.
Novel taxa described from collections of Nypa fruticans.
| Genus | Species 1 | Substratum (Collection Site) | Reference |
|---|---|---|---|
| Acuminatispora | Acuminatispora palmarum | On a submerged decayed petiole (Thailand) | [261] |
| Aniptodera intermedia * | On an intertidal petiole (Malaysia) | [166] | |
| A. nypae * | On intertidal fronds (Malaysia) | [116] | |
| Anthostomella nypae * | On an intertidal petiole (Malaysia) | [166] | |
| A. nypensis * | On an intertidal petiole (Malaysia) | [166] | |
| A. nypicola * | On an intertidal petiole (Malaysia) | [166] | |
| Apioclypea nypicola * | On an intertidal rachis (Malaysia) | [143] | |
| Arecophila nypae * | On intertidal palm tissues (Malaysia) | [131] | |
| Astrocystis nypae * | On an intertidal frond (Malaysia) | [150] | |
| A. selangorensis * | On a dead intertidal rachis (Malaysia) | [150] | |
| Astrosphaeriella nipicola (as A. nipaecola) (basio. Melanopsamma nipicola) | On palm tissues (Indonesia) | [144] | |
| A. nypae * | On decaying intertidal fronds (Brunei) | [162] | |
| Bacusphaeria | Bacusphaeria nypae * | On petiole base (Malaysia) | [269] |
| Carinispora | Carinispora nypae * | On decaying intertidal fronds (Brunei) | [162] |
| Delitschia nypae * | On a decaying fruit pericarp (Thailand) | [535] | |
| Fasciatispora | Fasciatispora nypae * | On intertidal rotten fronds (Brunei) | [161] |
| Frondicola | Frondicola tunitricuspis * | On decaying fronds | [162] |
| Helicascus nypae * | On intertidal dead fronds (Brunei) | [160] | |
| Helicorhoidion nypicola * | On intertidal palm tissues (Brunei) | [166] | |
| Herpotrichia nypicola * | On an intertidal petiole (Malaysia) | [166] | |
| Leptosphaeria nypicola * | On an intertidal petiole (Malaysia) | [166] | |
| Lignincola nypae * | On an intertidal petiole (Malaysia) | [166] | |
| Linocarpon angustatum * | On an intertidal petiole base (Malaysia) | [165] | |
| L. appendiculatum * | On rotten fronds (Brunei) | [154] | |
| L. bipolare (as L. bipolaris) * | On intertidal fronds (Brunei) | [105] | |
| L. longisporum * | On intertidal fronds (Brunei) | [105] | |
| L. nipae (syn. Ophiobolus nipae) * | On dead petioles (Philippines) | [154] | |
| Longicorpus | Longicorpus striatisporus (syn. Astrosphaeriella striatispora) | On fronds (Brunei) | [9] |
| Neolinocarpon | Neolinocarpon globosicarpum * | On decaying intertidal fronds (Brunei) | [162] |
| N. nypicola * | On an intertidal petiole base (Malaysia) | [165] | |
| Nipicola | Nipicola carbospora * | On immersed fronds (Brunei) | [163] |
| N. selangorensis * | On an intertidal frond (Malaysia) | [116] | |
| Nypaella | Nypaella frondicola * | On intertidal fronds (Brunei) | [164] |
| Oxydothis nypae * | On rotten fronds (Brunei) | [156] | |
| O. nypicola * | On a decayed petiole (Brunei) | [117] | |
| Phomatospora nypae * | On dead intertidal leaves (Malaysia) | [110] | |
| P. nypicola * | On an intertidal petiole (Malaysia) | [166] | |
| Plectophomella nypae * | On intertidal fronds (Brunei) | [164] | |
| Pleurophomopsis nypae * | On intertidal fronds (Brunei) | [164] | |
| Savoryella nypae (basio. Trichocladium nypae) * | On intertidal palm tissues (Brunei) | [166,619] | |
| Striatiguttula | Striatiguttula nypae * | On a decayed rachis (Thailand) | [9] |
| Tirisporella | Tirisporella beccariana * | On decaying leaf bases (Malaysia and Phlippines) | [167] |
| Vaginatispora nypae * | On a decaying fruit pericarp (Thailand) | [535] | |
| V. palmae * | On an immersed rachis (Thailand) | [761] | |
| Vibrissea nypicola * | On an intertidal petiole (Malaysia) | [166] |
1 The species only known from Nypa fruticans are noted with a superscript asterisk (*).
Trachycarpus fortunei has also been relatively well studied, as it is capable of thriving in warm temperate regions and occurs naturally in the warm temperate areas of China. Thus, this makes it possible to explore the differences between fungal diversity in temperate and tropical regions. Taylor et al. [215] isolated a total of 1728 identifiable fungal endophytes from 3256 frond samples of 10 mature T. fortunei individuals, including 75 species in 43 genera.
Some palm species are also more studied because they are commonly found in tropical rainforests and may be geographically restricted to certain regions, i.e., endemic. Thus, they are studied as a means of expanding knowledge about tropical mycology and addressing the biodiversity of fungi that inhabit tropical hotspots. In addition, some palm species that are geographically restricted could be recognised as a valuable source of new taxa, as discussed by Taylor et al. [194] for Archontophoenix alexandrae in Australia. As Taylor et al. [194] stated “its endemic nature and the relative geographic isolation of its natural habitat makes it a suitable candidate for studies relating to host-specificity of fungi and fungal biogeography”. In fact, it is likely that endemic host plants that have evolved in geographic isolation can be colonised by many novel fungi when studied in their natural environment. An impressive number of more than 35 new fungal species have been introduced based on collections of A. alexandrae, which is certainly associated with the endemic nature of this palm tree (Table 7). A similar pattern has also been observed in other palm species. For instance, Licuala palms have been frequently sampled in Brunei and Australia, where they are found naturally on tropical forests. Fröhlich and Hyde [10] recorded a total of 242 taxa, including 189 species of fungi, from 2672 isolates of six Licuala palms. Moreover, they reported a surprising estimate of 240 fungal species occurring on 3 individual Licuala palms and 155 on a Licuala single palm in a Brunei rainforest. This investigation considered only the endophytes, saprophytes, and pathogens that develop on the leaves and petioles. Likewise, a remarkable diversity of fungal endophytes was recorded by Fröhlich et al. [11] from 6 Licuala palms, including 73 species in 48 genera of 2237 isolates. Similar hight species richness has also been observed in palm species endemic to temperate regions, such as the New Zealand palm Rhopalostylis sapida [54]. Similarly, the subendemic Cuban palm Roystonea regia has often been investigated in Cuba for the isolation of a huge diversity of palmicolous “anamorphs”, particularly hyphomycetes. According to Mercado-Sierra et al. [329], more than 150 species and 60 genera of fungi have been recorded from collections of R. regia from Cuba, among which 15 new species and 5 new genera were introduced.
Table 7.
Novel taxa described from collections of Archontophoenix alexandrae.
| Genus | Species | Substratum (Collection Site) | Reference |
|---|---|---|---|
| Aegerita queenslandica | On a rotten leaf (Queensland, Australia) | [63] | |
| Anthostomella clypeosa | On a dead rachis (Queensland, Australia) | [8] | |
| Apioclypea nonapiospora | On a dead rachis (Hong Kong, China) | [8] | |
| Astrosphaeriella immersa | On a dead petiole (Hong Kong, China) | [148] | |
| Barriopsis archontophoenicis | On dead woody tissues (Thailand) | [549] | |
| Botryosphaeria archontophoenicis | On a dead petiole (Hong Kong, China) | [8] | |
| Chaetopsina alexandrae | On a dead rachis (Queensland, Australia) | [8] | |
| Heteroconium queenslandicum | On a rotten leaf (Queensland, Australia) | [63] | |
| Hydropisphaera ciliata | On a dead sheath (Queensland, Australia) | [8] | |
| Iodosphaeria hongkongensis | On a dead petiole (Hong Kong, China) | [146] | |
| Lasiosphaeria alexandrae | On a submerged rachis (Queensland, Australia) | [185] | |
| L. alexandricola | On a dead sheath (Hong Kong, China) | [185] | |
| Linocarpon australiense * | On palm tissues (Queensland, Australia) | [172] | |
| L. luteocollum | On a dead rachis (Queensland, Australia) | [8] | |
| Maculatipalma * | Maculatipalma fronsicola * | On a living (Queensland, Australia) | [197] |
| Manokwaria * | Manokwaria notabilis * | On a dead rachis on rainforest floor (Queensland, Australia) | [109] |
| Melanographium palmicola (as M. palmicolum) | On a decaying rachis (Hong Kong, China) | [182] | |
| Muyocopron hongkongense | On a dead rachis (Hong Kong, China) | [8] | |
| Neolinocarpon inconspicuum (as N. inconspicuus) | On a dead rachis (Queensland, Australia) | [140] | |
| N. nonappendiculatum (as N. nonappendiculatus) | On a dead petiole (Queensland, Australia) | [140] | |
| Neoxylaria queenslandica (as Xylaria queenslandica) | On a dead rachis (Queensland, Australia) | [8] | |
| Oxydothis alexandrarum | On a rotten rachis (Queensland, Australia) | [112] | |
| O. australiensis | On a rachis in forest litter (Queensland, Australia) | [112] | |
| Palmicola | Palmicola archontophoenicis | On a basal sheath of a fallen rachis (Queensland, Australia) | [108] |
| P. bipolaris | On a dead petiole (Queensland, Australia) | [8] | |
| Phomatospora archontophoenicis | On a dead rachis (Queensland, Australia) | [8] | |
| Pseudohalonectria eubenangeensis | On a dead rachis (Queensland, Australia) | [200] | |
| Pulmosphaeria | Pulmosphaeria archontophoenicis | On a dead petiole (Queensland, Australia) | [194] |
| Selenosporella queenslandica | On a rotten leaf (Queensland, Australia) | [63] | |
| Sorokina frondicola | On dead rachis (Queensland, Australia) | [8] | |
| Sporidesmium queenslandicum | On a rotten leaf (Queensland, Australia) | [63] | |
| Triadelphia archontophoenicicola (as T. australiensis) | On a dead rachis (Queensland, Australia) | [8] | |
| Tribulatia | Tribulatia appendicospora | On a dead petiole (Queensland, Australia) | [8] |
| Trichoconis queenslandica | On a rotten leaf (Queensland, Australia) | [63] | |
| Volutella queenslandica | On a rotten leaf (Queensland, Australia) | [63] |
* New taxa whose designated holotype material corresponds to collections of other palm species or unidentified palms and not Archontophoenix alexandrae, but which were also isolated from collection of A. alexandrae when introduced as new to science.
Several studies have also investigated the fungal communities on tissues of selected palms in tropical and subtropical regions, e.g., refs. [13,14], as well as in tropical and temperate regions, e.g., ref. [8]. Yanna et al. [13] identified 288 different taxa from fungal communities on decaying fronds of Livistona australis, Oraniopsis appendiculata (Australia), Arenga engleri, L. chinensis (Hong Kong), A. undulatifolia, Salacca affinis, and Oncosperma horridum (Brunei), among which 17 undescribed species were found. Similarly, Taylor and Hyde [8] studied the microfungi associated with three palm species in areas where they were native and where they had been introduced. They identified a total of 288 different taxa, including one new genus and 34 undescribed species, 26 of which showed host-specificity at species level.
All these figures demonstrate the extraordinary richness of palms for research into fungal biodiversity. The high number of fungal taxa found confirms that the fungi on palms are diverse and can be a source of many undescribed species. Furthermore, many palm species inhabit some of the world’s biodiversity hotspots, including areas of South America and India, where many palm species are native or even endemic. Although considered biodiversity-rich areas, both South America and India are poorly explored regions in terms of fungal diversity. Some studies have reported a high diversity of palm fungi from India and Brazil, e.g., refs. [437,501]. However, no comprehensive study has yet been carried out on the composition of palm fungi in these regions (see Section 2). Since a large number of fungi inhabit biodiversity hotspots, it is to be expected that many unknown fugal taxa inhabit palm trees native to these regions, especially considering the lack of studies in this regard.
Collecting fungi based on a chosen host is one of the most popular methods for studying fungal diversity. In addition to being fungi-rich and geographically widespread hosts, palms are mostly distributed in the tropical and subtropical regions of the world, some of which are underexplored biodiversity hotspots [590,595]. Thus, palms have several characteristics that increase the possibility of discovering new fungal species, which makes them ideal hosts for searching part of the worldwide unknown mycota. Therefore, palm trees should be prioritised for seeking new taxa and studying fungal diversity, given the ecological possibilities they can represent and reflect in the composition of their fungal assemblages. A number of studies have documented the role of palm trees and the corresponding palm fungal communities in biodiversity surveys. A notable example is the investigation carried out by Hyde and co-workers, which allowed estimates of fungal biodiversity to be questioned and adjusted to more reasonable values.
4.2. Palm Fungi and the Fungal Biodiversity Estimates
The studies carried out by Hyde and co-workers resulted in a wealth of data that provided new information for estimating fungal biodiversity. Much of these data were discussed and revised by Fröhlich and Hyde [10], who wondered whether the estimates of global fungal biodiversity at the time were realistic after the remarkable diversity observed in the fungal communities of palm trees in the tropics. For this reason, they considered that the estimate of 1.5 million species proposed by Hawksworth was a “very conservative estimate of the number of fungal species extant on the planet”. Furthermore, fungus to plant ratios seem to be noticeably higher on palms compared with those estimated by Hawksworth [727,728], when revisiting the numbers of fungal diversity on Earth. Thus, determining fungus to plant ratios on palms and, consequently, its contribution to estimates of fungal numbers is of great importance in fungal biodiversity surveys.
Hyde [762,763], reviewing his extensive work on palm fungi in North Queensland, estimated that there are about 3 pathogens, 10 saprophytes, and 100 endophytes that can develop on each palm species. In addition, Hyde [762,763] considered that 25% of these fungi are likely to be host-specific, i.e., restricted to a single host species (compared with the 67% host-specificity assumed by Hawksworth [727]). As a consequence, about 28 fungal taxa are likely to be associated with each palm species. This astounding plant to fungus ratio of 1:28 would imply the existence of almost 73,000 species of fungi on palms worldwide, of which only less than 3% (ca. 1580 species) were known [8]. However, following detailed investigation on six palm trees in Australia and Brunei carried out by Fröhlich and Hyde [10], the 1:28 ratio was subsequently revised upwards to 1:33. As Hyde [762,763] stated, after years of experience with palm fungi, it appears that “with palms the host species to fungi ratio is much higher”. Therefore, values ranging from 1:28 to 1:33 would be a more accurate estimate than the much lower and conservative plant:fungus ratios that have been estimated over the years [10,762].
The figures for plant:fungus ratios on palms are of marked significance for the total numbers of fungi worldwide. Plant:fungus ratios rely heavily on the concept of whether fungi are host- and/or tissue/organ-specific or have host- and/or tissue/organ-recurrence. Thus, host-specificity or -recurrence is probably the most important single factor used in estimating global fungal numbers [728,764]. The concepts of host-specificity and host-recurrence are not distinguished in the context of this review and are often used synonymously. However, host-specificity may be an inappropriate term for saprobic fungi. Zhou and Hyde [764] suggested host-exclusivity and host-recurrence as more appropriate terms (for a definition and discussion of these concepts, see Zhou and Hyde [764]).
There is now much circumstantial evidence that many palm fungi are host- and/or tissue-specific, and their impact on palm fungi numbers are discussed here. Many palm fungi have only been recorded on Arecaceae or sometimes on other large woody monocotyledons, such as Pandanaceae hosts. This could be due to similarities in the physical nature of the substratum of these plant families, which produce relatively large, thick leaves, which offer a range of microhabitats for fungal growth. Moreover, palm fungi differ widely from the taxa recorded on other monocotyledons, such as grasses.
4.2.1. Fungal Specificity at Family, Genus, and Species Levels
Host-specificity infers a relationship between hosts and fungi and has mostly been applied to plant pathogens. There are numerous examples of host- and tissue-specific plant pathogens [765,766,767,768]. However, most fungi on palms are not pathogens, and therefore are unlikely to be host-specific. They may, however, exhibit a host-recurrence, i.e., occur repeatedly on the same host, but be absent or rare on adjacent hosts of the same family [764].
An extremely high diversity of palm saprophytes was found developing on a wide range of dead palm material. Saprobic fungi are less likely to be host-specific [764]. However, in the great diversity of saprophytic fungi supported by palm tissues, many species are found exclusively or recurrently on palms. Therefore, it would be expected that some saprophytic fungi be selective to specific palm species or genera. This was found to be true and is well documented on mangrove palm trees, e.g., refs. [7,165]. At which level this specificity occurs, i.e., host genus, subtribe, tribe, or subfamily, is not yet obvious, but should become clear as the mycota of more palm hosts are systematically investigated. In fact, following his studies on the fungi on palms in North Queensland, Australia, Hyde [769] listed several species and genera of fungi that are thought to be unique to Archontophoenix alexandrae and other palm genera, as they have not been identified in detailed studies of other hosts occupying the same habitat. As Hyde [769] stated “these fungi are almost certainly genus-specific and some may also be host-specific” (Table 8).
Table 8.
Possible host-specific fungi known from a single palm species or genus in Australia (adapted and updated from [769]).
Table 8.
Possible host-specific fungi known from a single palm species or genus in Australia (adapted and updated from [769]).
| Palm Species/Genus | Fungal Species | Reference |
|---|---|---|
| Archontophoenix alexandrae | Hydropisphaera ciliata | [8] |
| Lasiosphaeria alexandrae | [185] | |
| Lockerbia palmicola * | [114] | |
| Neolinocarpon inconspicuum | [140] | |
| N. nonappendiculatum | [140] | |
| Oxydothis alexandrarum | [112] | |
| O. australiensis | [112] | |
| Palmicola archontophoenicis | [194] | |
| P. bipolaris | [8] | |
| Phomatospora archontophoenicis | [8] | |
| Pseudohalonectria eubenangeensis | [200] | |
| Pulmosphaeria archontophoenicis | [194] | |
| Calamus | Anthostomella bipileatus | [6] |
| Astrosphaeriella australiensis | [144] | |
| Cyanopulvis australiensis | [6] | |
| Neolinocarpon australiense | [140] | |
| Oxydothis calami | [117] | |
| O. luteaspora | [112] | |
| O. rubella | [112] | |
| O. uniseriata | [6] | |
| Pemphidium calamicola | [135] | |
| P. rattanicola | [6] | |
| Pseudohalonectria palmicola | [200] | |
| Roussoella calamicola | [147] | |
| Cocos nucifera | Mycosphaerella palmicola | [198] |
| Licuala | Ascotaiwania licualae | [6] |
| Capsulospora angustispora | [6] | |
| Nectriella erythroclypea | [121] | |
| Nipicola licualae | [6] | |
| Oxydothis angustispora | [6] | |
| O. cyrtospora | [6] | |
| O. extensa | [6] | |
| O. parasitica | [195] | |
| Linospadix | Oxydothis linospadicis | [195] |
| O. obducens | [117] | |
| Oraniopsis appendiculata | Monotosporella palmicola | [15] |
| M. sphaerica | [15] | |
| Palmaria montanea | [143] | |
| Sporidesmiella oraniopsidis | [230] | |
| Pinanga sp. | Phyllosticta candeloflamma | [187] |
* Although Lockerbia palmicola has been included in the present list as a possible host-specific fungal species of Archontophoenix alexandrae, the species was introduced by [114] on dead palm rachides on forest floors from North Queensland, Australia, which were referred to as “possibly Archontophoenix”. Thus, this fungal species may be specific to other Archontophoenix species, rather than A. alexandrae, or even to the genus Archontophoenix. Note: only fungal species known from more than one collection were included.
Although most palmicolous taxa will not be specific to a particular palm species, most of them belong to genera that specialise on palm hosts [17]. Many fungi that are saprobes on palms appear to be unique or occur disproportionately on palms relative to other hosts. Many genera typically found on palms, such as Arecomyces, Arecophila, Ascotaiwania, Manokwaria, Myelosperma, Neolinocarpon, Palmicola, and Pemphidium, comprise species known only from palms, while many speciose genera, such as Astrosphaeriella, Linocarpon, and Oxydothis, are predominantly found on palms [17,718,764]. Some of these genera were originally described from palm substrata (Table 1) and remain taxa that are apparently restricted to palms. In some cases, specificity seems to have a wider taxonomic range. For instance, members of the Phaeochoraceae (Phyllachorales), such as Cocoicola, Phaeochora, Phaeochoropsis, and Serenomyces, are saprotrophic or biotrophic ascomycetes on plant leaves that are apparently restricted to the Arecaceae [212]. Some examples of host-specificity suggested in palmicolous taxa are presented below, along with comments on the factors that can justify their specificity.
Although several Oxydothis species are known from more than one palm host, some are only known from a single palm host. This has been observed with O. alexandrae, which was frequently collected on decaying petioles of A. alexandrae but was not found on adjacent host palms, including Calamus or Licuala palms inhabiting the same habitat or region [12,117]. Thus, this taxon is a good example of an apparently saprobic fungus showing host-specificity. Hyde et al. [180] observed that species of Oxydothis and Cocoicola develop on the fronds before they completely dry out, suggesting that they may be endophytes that convert to a saprobic lifestyle with the onset of senescence. A similar situation occurs with Neolinocarpon nypicola on Nypa fruticans. The blackened stroma of this fungus forms throughout the senescing palm material, long before it has the appearance of being rotten [165]. In addition, it has recently been shown that saprobic Oxydothis species inhabiting dead palm tissues can produce appressoria by germinating ascospores [528]. Appressoria are specialised infection structures used by pathogenic taxa to infect their hosts and have rarely been observed in saprobic fungi, e.g., ref. [770]. The production of appressoria by saprobic taxa suggests that they may be adapted to an endophytic lifestyle and become active after host senescence [718,771]. The appressoria found on germinating ascospores of Oxydothis species from palms suggest that these taxa can infect healthy plants as endophytes, making them the first colonisers of dead palm material as saprobes [211].
Species of Astrosphaeriella are more commonly associated with the climbing or rattan palms, such as Calamus, Daemonorops, and Livistona, which indicate some degree of host-specificity [12]. For example, A. bakeriana is one of the first and most common taxa to appear on dead fronds and leaves of Livistona chinensis in Hong Kong and is hardly known from any other host, although it seems to be common throughout the Asian region [144]. Guo et al. [639], while studying the endophytes of L. chinensis, showed that this common saprobe is also an endophyte at an earlier stage. Many other fungi that were isolated as endophytes from palms have been shown to occur as common saprobes [639]. If many of the saprobes develop from endophytes, then it is likely that many saprobic fungi on palms are host-specific. As Guo et al. [639] pointed out “some endophytes and saprotrophs are interrelated, i.e., some saprotrophs have a latent period inside plant tissues, or some endophytes become saprotrophs after plants scenesce”. It is imperative to understand the mechanism of life mode conversion in fungi, as such conversions would have a significant impact on fungal diversity. The extraordinarily rich mycota found on palm trees in the tropics is likely to reveal new insights into this suggested change in life mode between endophytes and saprobes, or even between endophytes and pathogens. This fungus–host relationship, in which the plant tolerates the fungus in its tissues as an endophyte, is likely to have evolved over a long period, so it is likely to have resulted in saprobes having a host preference [763]. Thus, given that many fungi are specific or recurrent on palms, this may be one of the reasons why so many undescribed taxa are found on Arecaceae hosts. As Zhou and Hyde [764] stated, common fungal genera on palms may be endophytes that become saprobes on senescent plant parts, which would justify such high plant:fungus ratios in these hosts.
Most Neodeightonia species are exclusively or almost exclusively found on arecaceous hosts, and the co-evolution of Neodeightonia species as endophytes with these hosts to adapt to new environmental conditions has been recently discussed [547]. For example, N. phoenicum has only been reported from Phoenix spp. to date, so it is apparently restricted to palms and may represent an example of host-specificity at the genus level, e.g., ref. [548].
Many examples of host-specific fungi have also been suggested in palmicolous hyphomycetes. Brachysporiella species are mainly found on palms, although they also grow in other habitats [103]. Other genera, such as Ceratosporella, Endocalix, Piricauda, Phragmospathula, and Phragmospathulella, are practically exclusive to palms. In some cases, such as Holubovaea and Consetiella, specificity even appears at the level of host genus or host species, in this case Roystonea regia [329,352]. However, in cosmopolitan or speciose genera of hyphomycetes, this specificity is often lost. Speciose genera are more heterogeneous and, in turn, their species are more genetically diverse. Greater genetic diversity leads to greater adaptability to the environment, so they can be found in palm trees and other substrata [329]. An interesting case of host-specificity is the species Holubovaea roystoneicola described on petioles of R. regia from Cuba [73]. There is no record of this species on a different host plant since its introduction 40 years ago, nor in any other region outside Cuba. However, it has been collected more than 140 times in different Cuban localities and always inhabiting R. regia [352].
One remarkable study that gave new insights into the issue of host-specificity was that of Fröhlich and Hyde [10]. They compared the fungal communities on three Licuala ramsayi palms in Northern Queensland, Australia, and on a different unidentified species of Licuala in a pristine tropical rainforest in Brunei Darussalam. Only 30 of 242 taxa overlapped between the fungal communities recorded on both Licuala species, although some of these taxa may have been misidentified, as it was not possible to resolve them using molecular data. As Hyde et al. [756] asked, if fungal species were not mainly host- or genus-specific, how did almost completely different communities occur on these palm species of the same genus but in different countries?
Studies on the fungal communities of different terrestrial palm species have suggested that both host genera and host species affect the composition of these communities in relation to the fungal species recovered from palm tissues (Table 9). Yanna et al. [13] studied the fungal composition of communities recovered from decaying fronds of seven palm species in Australia, Brunei, and Hong Kong and reported that few fungi were common to palms of the same genus and the number was lower on palms of different genera. In Brunei and Hong Kong, only 10% and 17%, respectively, of the fungi recorded were common to palms of different genera, while in Australia the numbers were even lower, ranging from 6 to 9%. Furthermore, only 5 to 23% of the fungi were common to two of the palms studied. These figures strongly suggest evidence for host-specificity on different palm host genera. Similarly, Yanna et al. [634] and Taylor and Hyde [8] studied fungal communities on Phoenix hanceana and A. alexandrae in Hong Kong, respectively, and fungi overlapping with those on other palm hosts in Hong Kong was very low.
Table 9.
Ten most common species recorded on selected terrestrial palm species in ecological studies conducted in different countries (adapted from [16]).
Table 9.
Ten most common species recorded on selected terrestrial palm species in ecological studies conducted in different countries (adapted from [16]).
| Arenga engleri (Hong Kong) | Arenga undulatifolia (Brunei) | Calamus sp. (Thailand) | Livistona chinensis (Hong Kong) | Oncosperma horridum (Brunei) | Phoenix hanceana (Hong Kong) | Salacca affinis (Brunei) |
|---|---|---|---|---|---|---|
| Piricauda cochinensis | Piricauda cochinensis | Tetraploa sp. | Astrosphaeriella bakeriana | Linocarpon livistonae | Diplococcium stoveri | Zygosporium minus |
| Diplococcium stoveri | Melanographium selemiodes | Morenoina palmicola | Lachnum palmae | Craspedodydimum nigroseptatum | Endocalyx cinctus | Linocarpon livistinae |
| Helminthosporium solani | Trichoderma harzianum | Circinoconis paradoxa | Appendicospora hongkongensis | Zygosporium minus | Cryptophiale udagawae | Peltistromella anomala |
| Melanographium palmicola | Zygosporium minus | Diaporthe sp. | Monodictys putredinis | Monotosporella setosa var. macrospora | Penzigomyces nodipes | Helicosporium griseum |
| Melanographium selenioides | Pleurophragmium sp. | Helminthosporium sp. | Oxydothis elaeicola | Neolinocarpon australiense | Thozetella effusa | Volutella ciliata |
| Monodictys putredinis | Helmithosporium velutimum | Linocarpon sp. | Trichoderma harzianum | Trichoderma harzianum | Pseudospiropes simplex | Oxydothis luteaspora |
| Oxydothis ragai | Volutella ciliata | Phaeosphaeria sp. | Neolinocarpon australiense | Oxydothis luteaspora | Dictyochaeta simplex | Periconiella sp. |
| Pestalotiopsis palmarum | Peltistromella anomala | Anthostomella sp. | Fasciatispora petrakii | Oxydothis licualae | Serenomyces shearii | Arecomyces bruneiensis |
| Guignardia manokwaria | Stachylidium sp. | Astrosphaeriella sp. 1 | Corynesporopsis isabelicae | Oxydothis elaeicola | Capsulospora brunneispora | Sporidesmium parvum |
| Dischoridium roseum | Anthostomella minutoides | Goidanichiella fusiformis | Dictyosporium elegans | Brachysporiella gayana | Harknessia globosa | Codinaea intermedia |
Note: the taxa are listed in descending order of their percentage abundance. The taxa listed are only those identified at least to genus level; unidentified taxa have been disregarded. The taxa names are presented according to the literature used to construct the table, regardless of whether their current classification is different.
Host-specificity has also been strongly suggested on the fungal communities of palms inhabiting peat swamp and mangrove forests. Pinnoi et al. [248] and Pinruan et al. [249] documented the fungal communities that occur on the peat swamp palms Eleiodoxa conferta and Licuala longicalycata, respectively. They observed a very low overlap between the fungi and the dominant mycota found on terrestrial palms. While genera such as Anthostomella, Arecomyces, Linocarpon, Oxydothis, and Sorokinella are generally common on terrestrial palms, but do not usually constitute a dominant group on peat swamp palms, showing a marked difference in the percentage of occurrence, species of Astrosphaeriella tend to be common to both terrestrial and peat swamp palms. Recently, these data were revisited and analysed by Pinruan et al. [669], who also documented the diversity of fungi occurring on two other peat swamp palms, Metroxylon sagu and Nenga pumila. These studies report either some taxa common on submerged wood (e.g., Brocchiosphaera brocchiata, Dictyochaeta gyrosetula and Thozetella nivea) and many taxa common to those known to come from terrestrial palms (e.g., Astrosphaeriella spp., Linocarpon spp., Massarina and Oxydothis). However, the percentage of overlap between these communities is low and most of the taxa found are exclusive to peat swamp palms and have never been documented in other habitats. When analysing the percentage of overlap in fungal biodiversity between the four peat swamp palms, Pinruan et al. [669] reported that of the ten most common fungal species, or even genera, occurring on each of them, no taxa was found to be common to the four palms (Table 10). The results reinforce that each of the four peat swamp palms supports its own specific fungal community. Moreover, the overlap of fungal species between the four palms was less than 1%, while between three and two it was less than 2% and less or 6%, respectively, even though they all grew in close proximity in a peat swamp forest in Thailand (Figure 2).
Table 10.
Ten most common genera and species recorded on each and all of the four peat swamp palm species in ecological studies conducted in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand (adapted from [669]).
Table 10.
Ten most common genera and species recorded on each and all of the four peat swamp palm species in ecological studies conducted in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand (adapted from [669]).
| Taxonomic Rank | Eleiodoxa conferta | Licuala longicalycata | Metroxylon sagu | Nenga pumila | Peat Swamp Palms |
|---|---|---|---|---|---|
| Genera | Astrosphaeriella | Astrosphaeriella | Nawawia | Diplococcium | Astrosphaeriella |
| Stilbohypoxylon | Oxydothis | Anthostomella | Dinemasporium | Microthyrium | |
| Cancellidium | Annulatascus | Oxydothis | Linocarpon | Stilbohypoxylon | |
| Xylomyces | Massarina | Apiospora | Arecomyces | Cancellidium | |
| Lophiostoma | Microthyrium | Cylindrocladium | Spadicoides | Diplococcium | |
| Microthyrium | Phaeoisaria | Dinemasporium | Lophodermium | Oxydothis | |
| Morenoina | Nectria | Tetraploa | Sporidesmium | Xylomyces | |
| Phaeoisaria | Phruensis | Apioclypea | Dactylaria | Lophiostoma | |
| Jahnula | Submersisphaeria | Ornatispora | Oxydothis | Phaeoisaria | |
| Annulatascus | Thozetella | Massarina | Jahnula | Annulatascus | |
| Species | Cancellidium applanatum | Microthyrium sp. | Anthostomella bipapillispora | Diplococcium stoveri | Microthyrium sp. |
| Xylomyces aquaticus | Phaeoisaria clematidis | Nawawia filiformis | Dinemasporium sp. | Cancellidium applanatum | |
| Astrosphaeriella aquatica-like | Annulatascus velatispora | Oxydothis-like | Arecomyces epigeni | Diplococcium stoveri | |
| Stilbohypoxylon elaeicola | Massarina bipolaris | Apioclypea eccentricospora | Linocarpon sp. 4 | Xylomyces aquaticus | |
| Lophiostoma frondisubmersa | Phruensis brunneispora | Apiospora sp. | Lophodermium sp. | Phaeoisaria clematidis | |
| Microthyrium sp. | Solheimia costaspora | Dinemasporium lanatum | Dactylaria palmae | Astrosphaeriella aquatica -like | |
| Morenoina palmicola | Thailandiomyces bisetulosus | Tetraploa aristata | Lophiostoma sp. | Stilbohypoxylon elaeicola | |
| Phaeoisaria clematidis | Nectria sp. 1 | Ornatispora sp. | Oxydothis sp. 8 | Jahnula appendiculata | |
| Stilbohypoxylon eleiodoxae | Helicoma sp. 1 | Massarina bipolaris | Spadicoides sp. 4 | Lophiostoma frondisubmersa | |
| Jahnula appendiculata | Astrosphaeriella malayensis | Acrogenospora sphaerocephala | Jahnula appendiculata | Morenoina palmicola |
Note: the taxa are listed in descending order of their percentage abundance. The taxa names are presented according to the literature used to construct the table, regardless of whether their current classification is different.
A very low overlap in fungal diversity is similarly observed between peat swamp palms and the mangrove palm N. fruticans. None of the 10 most common fungal species on each peat swamp palm studied has been recorded on N. fruticans, even though its fungal diversity has been well documented in Brunei, e.g., refs. [154,162], Malaysia, e.g., refs. [110,165], the Philippines, e.g., refs. [673,772], Thailand, e.g., refs. [9,162,261], and Indonesia, e.g., ref. [117] (Table 11). Likewise, few of the fungi recorded on N. fruticans have been recorded inhabiting peat swamp palms [674]. When compared with the peat swamp palm E. conferta, only a few species are common to both palms in Astrosphaeriella, Linocarpon, and Oxydothis. However, the genera Carinispora, Fasciatispora, Halocyphina, Helicascus, Lignincola, and Lulworthia, which are common on N. fruticans, have not been recorded on E. conferta, as these genera are more commonly found on substrata in marine habitats and may require salt for growth, while those on E. conferta may not be salt tolerant. The latter may be more tolerant to acidic waters, while marine fungi tend to occur in more alkaline waters [248,674]. A similar pattern of low overlap in fungal composition is observed between the peat swamp palm L. longicalycata and N. fruticans, although some taxa are common to both palms, including species of Helicoma, Helicosporium and Thozetella [249,674]. Thus, most of the fungi found on Nipa palm is intertidal and do not appear to occur on other palms. In addition, species composition on this palm also differs from that on mangrove wood [168,671].
Nypa fruticans grows in the brackish waters of the intertidal region along marine coastlines in the tropics and extends into freshwater zones. This highly specialised habitat makes N. fruticans an interesting host for fungal colonisation. Fungi occurring on N. fruticans can be categorised into three main groups: typically marine/mangrove fungi (e.g., Aniptodera chesapeakensis, Halocyphina villosa, Kallichroma tethys, Marinosphaera mangrovei, Lignincola laevis, Lulworthia spp., Savoryella paucispora, Saagaromyces ratnagiriensis, Sammeyersia grandispora and Verruculina enalia), many of which appear to be host-specific (e.g., Aniptodera nypae, Fasciatispora nypae, Helicascus nypae, Helicorhoidion nypicola, Lignincola nypae, Savoryella nypae, and Tirisporella beccariana); typically freshwater fungi (e.g., Anthostomella eructans, Annulatascus velatisporus, Helicoma hongkongense, H. hyalonemum, H. pannosum, and Thozetella nivea); and fungi from typical palm-inhabiting fungal genera, most of which also appear to be exclusive to this palm (e.g., Anthostomella nypae, Linocarpon angustatum, L. appendiculatum, L. bipolare, L. nipae, Oxydothis nypae, and O. nypicola) [671,672,673]. To-date, 142 fungi have been documented growing on N. fruticans, of which 42 are only known from this host [9,261,535,597,674,761] (Table 6). Thus, one can assume ca. 30% of host-specificity for the fungal species recorded on N. fruticans. In fact, N. fruticans appears to support a large number of unique fungi, which do not overlap with those occurring on terrestrial palms [13].
Table 11.
Ten most common species recorded on Nypa fruticans in different ecological studies conducted in different countries.
Table 11.
Ten most common species recorded on Nypa fruticans in different ecological studies conducted in different countries.
| Thailand 1 | Brunei 2 | Philippines 3 |
|---|---|---|
| Trichocladium nypae | Linocarpon bipolare | Linocarpon appendiculatum |
| Linocarpon appendiculatum | Linocarpon appendiculatum | Microthyrium sp. |
| Lulworthia grandispora | Oxydothis nypae | Astrosphaeriella striatispora |
| Oxydothis nypae | Astrosphaeriella striatispora | Oxydothis nypicola |
| Astrosphaeriella striatispora | Trichocladium nypae | Halocyphina villosa |
| Helicorhoidion nypicola | Lignincola nypae | Didymella sp. |
| Aniptodera nypae | Neolinocarpon globosicarpum | Lignincola nypae |
| Lignincola laevis | Sporidesmium crassisporum | Helicorhoidion nypicola |
| Dictyosporium elegans | Helicorhoidion nypicola | Aniptodera intermedia |
| Anthostomella cf. rehmii | Aniptodera nypae | Massarina sp. |
It should be noted that most of the palms studied were only one species of a given genus, so it is unclear whether the data obtained is the result of host- or genus-specificity. Some cases of host-specificity seem to be well established, such as that observed in fungal communities of N. fruticans, which is the only species in the Nypa genus. There are other intertidal mangrove palms, such as Calamus erinaceus, Oncosperma tigillarium, and Phoenix paludosa. It is therefore necessary to examine the fungi on these hosts to determine whether there is overlap in fungal communities. This will certainly fine-tune the percentage of host-specificity predicted for N. fruticans. However, N. fruticans is notable for the fact that more than 40 fungal species are exclusive to this host. Thus, although it is unclear whether the same fungi also occur on the other three intertidal palms, even if they did, the ratio of these palm hosts to specific intertidal fungi is extremely high.
4.2.2. Fungal Specificity at Organ/Tissue Level
In addition to host-specificity at the plant family, genus, and species levels, studies on palm fungi have also revealed host-specificity at the organ/tissue level. Palm trees comprise several different types of tissue with different textures and chemistries, including roots, trunks, petioles, rachides, leaves, and flowers. For instance, species of Astrosphaeriella are particularly common on the aerial stems or trunks of climbing or rattan palms, which suggests some degree of tissue-specificity [12]. In fact, different palm tissues have been found to support different assemblages of saprobic and endophytic fungi [17], and this difference is probably due to substratum structure. For instance, Hyde et al. [17] reviewed the ascomycetes reported on palms and compiled a list of some common genera reported from different palms structures. These include species of Anthostomella, Lembosia, Meliola, Mycosphaerella, Phyllachora, and Sphaerodothis predominantly found on leaves, and species of Anthostomella, Astrosphaeriella, Linocarpon, Oxydothis, Rosellinia, and Xylaria predominantly found on rachides. Likewise, Hyde et al. [17] reported that the few taxa described from palm inflorescences were not found in other tissues. Thus, fungi associated with the reproductive tissues of palms are likely to form a distinct assemblage of species when compared with other palm fungal communities [10]. This was found to be true in several palm fungal communities recovered from different palm tissues.
Tissue-specificity has been suggested in endophytic palm fungal communities. A higher isolation rate, i.e., the recovery of a greater number of isolates, is often observed in vein rather than intervein tissues in both tropical [11,291,469] and temperate [215] palmicolous endophytes. Moreover, differences are often observed between the endophytic mycotas of different palm tissues and the tissues of different ages [11,215]. Tissue-specificity has also been suggested by the preference of xylariaceous taxa for leaf tissues [11,215] and of coelomycetes taxa for petioles [11].
Hyde and Alias [7] found that different fungi colonised different parts of the fronds of Nypa fruticans, including the leaves, leaf midribs, petioles, and petiole bases, indicating that some fungi may develop preferentially on certain types of tissue. Similar results were found on terrestrial palms. Yanna et al. [13] investigated the effect of different parts of the decaying fronds of seven different palm species on fungal communities. They reported that distinct fungal communities occurred on the leaves and rachis-tips, mid-rachides, and rachis-bases of most of the palm species examined. The exception was Livistona australis in which the fungal communities occurring on different parts of the rachides were more similar, probably due to their similar structures. However, the fungi on the leaves were distinct. Yanna et al. [14] investigated the effect of different parts of the decaying fronds of L. chinensis from Hong Kong on fungal communities. Distinct fungal communities were also observed, insomuch that 25 to 70% of the fungi recorded during different periods of decay were restricted on either leaves or petioles, which included 20 and 54 of 91 species confined to leaves and to petioles, respectively. For example, Appendicospora hongkongensis and Cocoicola livistonicola have only been recorded on petioles, while Pseudospiropes arecacensis and Vesiculozygosporium echinosporum (syn. Zygosporium echinosporum) have only been recorded on leaves [14]. Pinnoi et al. [16] also found that the petioles of Calamus spp. supported a greater species diversity than rachides (61% versus 39%, respectively) and while many species, such as Melanographium citri, Astrosphaeriella vesuvius, and Berkleasmium micronesiacum (syn. Coleodictyospora micronesiaca), were confined to petioles, only Lachnellula sp. occurred exclusively on rachides. Likewise, fungi were found to be more abundant and diverse on the petioles of the peat swamp palms Eleiodoxa conferta [248] and Licuala longicalycata [249] than on their trunks, rachides, and leaves (53% on petioles versus 30% on rachides and 17% on leaves for E. conferta; 61% on petioles versus 24% on trunks and 15% on leaves for L. longicalycata). Moreover, the percentage of overlapping fungi found in all the tissues examined of L. longicalycata were only 0.3%, while 69.7% of them were only found on the petioles (versus 8.9% only found on trunks and 8.9% only found on leaves) [249]. Although only preliminary results have been retrieved, a similar pattern of petioles supporting the greatest number of fungal records and diversity were also found for the peat swamp palms Metroxylon sagu and Nenga pumila [669].
Palm petioles have long been recognised as an ideal substratum for the development of a wide diversity of fungi and their structure has been identified as a crucial factor in establishing an intimate fungus–host relationship. This fungus–host relationship was first mentioned by Mercado-Sierra [73] for the rotten and large sheathing petioles of Roystonea regia and later discussed for other Cuban palms [329,352]. The petioles of R. regia are very long (1.5–2 m) and wide. Thus, upon decomposition, the adequate surface available for the development of hyphomycetes is very large, and much larger than that of other plants, which are, therefore, subjected to a higher level of competition than palm trees [329]. Furthermore, the diversity of taxa was also considered remarkable. Mercado-Sierra [73] hypothesised that this should be related to the very long period of active leaf growth in R. regia, which allows a high concentration of nutrients that can be used by the fungi that inhabit its tissues.
Some studies have also suggested that the morphological and anatomical structures of palms can affect the composition and appearance of the taxa that colonise palm fronds. For instance, Hyde and Cannon [212] reported that the heavily lignified and robust arrangement of palm vascular bundles seems to affect the development and final appearance of some fungal fructifications. These fructifications tend to be elongated, so that they fill the tissue between the veins rather than growing over or through them. They are often erumpent or inserted between the outer layers of the host tissue. The more deeply immersed ascomata can often be seen developing between vascular bundles and having their walls distorted by them [212]. The hardness and arrangement of the veins in palm tissues seem to also influence the taxa that use them, which may reveal some degree of tissue-specificity. This has been observed in fungi that cause tar spots on palms. For example, relatively few Phyllachora species are found on Arecaceae when compared with other host families, and the species that do occur are limited to tissues with low levels of structural components and small amounts of lignification, such as the leaf blade [212].
The anatomical structure of palm tissues has also been reported as the main factor that determines the greater abundance and diversity of fungal assemblages on petioles, trunks, and rachides when compared with leaves [13,14,16,248,249,669]. While leaves contain mainly thin-walled, starch-rich parenchymatous cells, petioles have more thick-walled sclerenchymatous cells with associated vascular bundles [773], which can take up water and retain moisture for a longer time. Thus, thicker cell walls can yield more nutrients, namely cellulose and lignin, for sustained fungal growth. These anatomical differences between palm tissues are likely the reason why tissue-specificity has been widely suggested in palm fungal communities. The repeated occurrence of certain fungi on different types of tissue may result from different nutritional requirements of fungi or their ability to utilize different substrata due to the production of specific enzymes [13,14,249]. In addition, palm petioles are structurally more robust, with more concentrated supportive tissue than leaves, and do not decompose as quickly [773], thus allowing time for a more complex fungal community to form and a succession of different fungi to develop [10].
Many other issues can affect the composition of palm fungal communities, which will therefore also have important implications for fungal estimates. These include, for instance, fungal succession and the existence of different microhabitats, as well as fungal co-occurrence patterns and geography. Fungal succession on palms have been studied on L. chinensis and Phoenix hanceana from Hong Kong [14,634]. These studies have shown that there is a sequential order in which fungi appear on substrata as they decay, so that different fungal communities are established on different tissues during the decomposition process, with certain species associated exclusively or primarily with certain palm tissues. These results are in line with the evidence that some of the early colonisers of palm substrata are derived from endophytes and, therefore, likely to be host-specific [764]. In fact, as Hyde et al. [718] stated, if “different fungi colonise substrata at different stages of decay, this has important implications for fungi numbers”. Similarly, studies on both terrestrial [16] and peat swamp [248,249] palms have shown that palm parts exposed under different microhabitat conditions, such as dry and wet/submerged palm material, showed differences in their fungal communities and, therefore, it is likely that the existence of these different conditions also influence the fungal diversity and the suggested taxa specificity. In addition, the specificity observed can also be influenced by the existence of certain biotic relationships, such as competition, which can inhibit the establishment of certain species and may enhance the co-occurrence of others. This has recently been discussed for fungal communities on N. fruticans [675]. Likewise, studies on saprobic and endophytic palm fungi have provided an indication that site-specific factors and geographical distance may be important in shaping fungal assemblages. In fact, if the same host taxa occurring in different countries support similar or different fungal assemblages, this will have important implications for fungal estimates.
4.3. Palm Fungi as Good Biogeographical Indicators
Palm fungal communities have shown distinct patterns in relation to their collection site and geography, which in some cases seems to strongly affect their qualitative and quantitative composition. For instance, Yanna et al. [13] showed that fungal species composition was significantly affected by the site of collection. There were few taxa common (5 to 16%) to palms from different sites, insomuch that distinct fungal communities were observed on samples from Australia, Brunei, and Hong Kong. In addition, the fungal composition on palms from either different or the same genera at different sites of collection were less coherent than those from the same sites [13]. Taylor et al. [12,215] have also shown variation between geographically separated communities of endophytic fungi in Trachycarpus fortunei, as well as saprobes on other palm hosts.
The close association of palm fungi with palm hosts suggests that they are good biogeographical indicators. Thus, they are a particularly important group of fungi for studying the biogeographical distribution of fungi, which is a challenging task in fungal biodiversity surveys. In fact, studies likely to provide good biogeographical data are those involving fungi that coevolved or are intimately associated with their hosts [774]. This is well documented in palm fungal communities, especially by the number of host-specific fungi estimated for palm hosts [10]. In addition, some palm fungi recorded as saprobes have also been recovered as endophytes, which emphasises their close relationship with palm hosts [215].
Climate has proved to be an influential factor affecting the distribution of fungi associated with palm trees. Taylor et al. [12] reported different assemblages of fungi associated with palms in temperate regions as compared with those in tropical regions, with differences being more related to climatic influences than to the hosts sampled. In temperate regions, the dominant tropical palm mycota were replaced by more ubiquitous, plurivorous ascomycetes, in addition to fungi of different groups, such as coelomycetes. The tropical palm Archontophoenix alexandrae presented a largely distinct palmicolous mycota within its natural biogeographical range, when compared with the taxa recovered outside of the palm’s natural habitat. An assemblage depauperated in typical palmicolous taxa, but with representatives of widespread tropical taxa of a more plurivorous nature, was recovered from A. alexandrae planted outside its natural habitat [12].
Fröhlich and Hyde [6] also observed that the differences and similarities between the palm mycota recovered from Australia, Brunei, and Hong Kong are likely to be chiefly influenced by three factors, namely past and present biogeography, host distribution, and climate. According to Fröhlich and Hyde [6], while biogeography and host distribution seems to have a dominant effect on the distribution of fungi at the genus level, climate seems to be more important in determining the distribution of species. Distribution of fungi in the Old World Tropics followed patterns consistent with climate, rather than past and present biogeography and host distribution. Thus, palmicolous ascomycete assemblages from Hong Kong and Australia were found more similar despite being in different hemispheres [6]. Yanna et al. [13] and Taylor and Hyde [8] also noted that geographical distribution significantly affects palm fungal communities, regardless of the host.
The data available for palm fungi biogeography are incomplete and fragmentary [12]. However, some patterns were noted by Fröhlich and Hyde [6], mostly from the collection of palm fungi in four countries, viz. Australia, Brunei, Ecuador, and Hong Kong. The typical palm fungi found in the tropics, such as Linocarpon, Astrosphaeriella, Oxydothis, Anthostomella, Arecomyces, Lophiostoma, and Capsulospora, seems to have the same pantropical distribution as their hosts. Thus, the same genera, or even species, have been recorded on both sides of the equator, in both the Old and New World Tropics. Fröhlich and Hyde [6] noted no significant variation in species richness of the different tropical countries studied. However, the relative abundance of the most common species was different in the New and Old World Tropics. For instance, species of Arecomyces are more frequently found in Ecuador, while species of Oxydothis are more frequently found in Southeast Asia and Australia.
5. Why Study Palm Fungi? Biodiversity Estimates and Their Significance
In the wealth of data obtained from the extensive investigation on palm fungi, a diverse and abundant assemblage of host- and tissue-specific fungi was found (see Section 4). This certainly accounts for the high fungal diversity recorded on palms, and subsequently accounts for the largely unknown number of fungal species estimated. Furthermore, it has been shown that many other important factors in the estimation of fungal diversity determine the occurrence of distinct palm fungal communities, including the existence of different microhabitats, site of collection, and fungal succession. However, while palm fungi appear to be an important source for the description of many of the unknown fungal taxa, the extent of their contribution is yet to be determined. Considering the figures previously overviewed, an updated estimate of the number of fungal taxa occurring on palms worldwide is presented and discussed here, and its impact on the fungal biodiversity estimates is also noted.
Several different benchmarks can be used to assess estimates of fungal diversity on palm trees. In addition, it can be assumed that their mean value may be closer to true biological reality, since it will accommodate and integrate all those differences mentioned in the literature. One might wonder why it would be important to accommodate these differences and the answer is basically related to the remarkable extent of latitude in which palm trees can thrive and exploit ecosystems. Palms occupy a great diversity of habitats, from tropical rainforests to deserts. This reflects their impressive adaptability to diverse climatic conditions, which is easily observed in the abundance of palms in temperate regions, although the overwhelming majority are native to tropical climates [5]. Considering that palm fungi are good biogeographical indicators, due to their close association with palm hosts, their communities can provide important biogeographical data, since they comprise fungi that coevolved or are closely associated with palms [12]. Thus, it is to be expected that differences in palm fungal communities, expressed in both their qualitative and quantitative composition, will be observed when accessing the fungal composition of palms that occupy extremely distinct habitats, such as those found in temperate and tropical ecosystems.
Considering the most recent literature available, palms comprise around 2,600 species in 181 genera [1]. As previously mentioned, after years of research into palm fungi, Hyde [762,763] estimated that there were approximately three pathogens, ten saprobes, and one hundred endophytes for each species of palm. Thus, the number of ca. 113 fungal taxa developing on each palm species is used here to predict the number of fungal species expected to occur on palms worldwide.
Benchmark I—Plant:fungus ratios in tropical palms. Hyde [762,763] considered that 25% of the fungi that occur on palms would be host-specific, which means that 28 specific fungal taxa are likely to be associated with each palm species. A plant:fungus ratio of 1:28 would imply the existence of 67,600 species of palm fungi worldwide (Table 12). Later, the above ratio was revised upwards, and Fröhlich and Hyde [10] predicted that 33 specific fungal taxa are likely to be associated with each palm species. A plant:fungus ratio of 1:33 would imply the existence of 85,800 species of palm fungi worldwide (Table 12). Naturally, these figures, with an average value of 76,700 species of fungi on palms, are a reference for palms inhabiting tropical regions, where fungal diversity is expected to be higher compared with temperate regions. As has long been observed, plant:fungus ratios are expected to vary depending on the geographical location [10,727]. Therefore, it is expected that the plant:fungus ratio in palms native to temperate regions will be lower, as the fungal communities of temperate palms tend to be less diverse than their tropical counterparts.
Benchmark II—Plant:fungus ratios in temperate palms. Hawksworth and Lücking [745], while revisiting estimates of fungal diversity on Earth, estimated that plant:fungus ratios range from 1:8 to 1:19.1, with an average value of 1:9.8, a considerably higher ratio than the conservative 1:6 estimated in 1991 [727]. Interestingly, this new estimated ratio of 9.8 unique fungal species to vascular plants is based on data obtained through field surveys and molecular approaches. Thus, this figure naturally includes much more information on fungal diversity than studies that are based solely on field surveys. In fact, field surveys are biased towards recording certain groups of fungi, while others remain underexplored or even undetected. Furthermore, most of the studies considered by Hawksworth and Lücking [745] were long-term investigations based on collections of fungi from temperate regions. For example, studies on Esher Common (Surrey, England, UK), the site most investigated by field mycologists in the world, have produced a plant:fungus ratio of 1:8, which is remarkably close to the average value of 1:9.8. In this sense, the plant:fungus ratio of 1:9.8 can be seen as a more appropriate value for estimating fungal diversity in temperate vascular plants. Considering that this plant:fungus ratio is accurate for temperate palm trees, this would imply the existence of 25,480 species of palm fungi worldwide (Table 12). Although the above approach represents an interpolation of the studies by Hawksworth and Lücking [745], studies on palms can also be used to assess the plant:fungus ratio in temperate palms. Taylor et al. [12] studied the biogeographical distribution of microfungi associated with palms from tropical and temperate habitats and estimated a number of potentially host-specific fungi ranging from three to thirteen species. As expected, palms from tropical regions showed a higher number of host-specific fungi, namely thirteen for Archontophoenix alexandrae and ten for Cocos nucifera, than the three specific fungi found on Chamaerops humilis, which is native to temperate regions. The average plant:fungus ratio of 1:8.7 interpolated from the data obtained by Taylor et al. [12] is lower than those suggested for tropical palm hosts (1:26–33) [10]. However, it is remarkably closer to the number of specific fungi estimated for hosts in temperate regions (1:8) [745]. Furthermore, the plant:fungus ratio estimated by Taylor et al. [12] may be highly influenced by the data obtained for Chamaerops humilis, as well as by the experimental set-up. This included palm hosts outside their natural geographic range, which will naturally influence their fungal composition and may be the reason why tropical palms showed plant:fungus ratios more typical of temperate climate hosts. In this sense, a plant:fungus ratio of 1:8.7 is considered here to be an adequate approximation for temperate palms and would imply the existence of 22,620 species of palm fungi worldwide (Table 12). Thus, an average value of 24,050 species of palm fungi can be assumed for palms inhabiting temperate regions.
Benchmark III—Plant:fungus ratios in palms inhabiting highly specialised habitats. Considering the previous extensive discussion on host-specificity in palm fungi, it appears that some palm species, particularly those inhabiting exceptionally unique and diverse habitats, may have a relatively higher percentage of host-specificity than those 25% estimated by Hyde [762,763]. Studies on peat swamp palms have revealed exceptionally diverse and distinct communities recorded on palms growing in close proximity, with the description of several new species. Only less than 1%, 2%, and 6% overlap in fungal species was observed between four, three, and two, respectively, of the peat swamp palms investigated [669] (Figure 2). Although some collections can be considered as preliminary results, the overlap between fungal communities is incredibly low. In turn, this may reflect a higher plant:fungus ratio than that considered for typically tropical terrestrial palms, which probably results in a high number of host-specific fungi. Similarly, a host-specificity of ca. 30%, higher than the 25% suggested by Hyde [762,763], has been estimated here for Nypa fruticans, a mangrove palm that inhabits an exceptionally unique ecosystem. This percentage of host-specificity was calculated taking it to account that 42 of the 142 fungal species recorded on this palm are host-specific (Table 6). Future collections, including collections from mangrove formations in other countries, may reveal an even higher percentage of host-specificity, as new species inhabiting N. fruticans tissues are continuously being described, e.g., ref. [9]. It seems reasonable to consider that 25% of host-specificity may be a conservative estimate for palms inhabiting highly specialised habitats. Using a plant:fungus ratio of 1:42 as an illustrative figure of current knowledge about fungal diversity on N. fruticans, this would imply the existence of 109,200 species of palm fungi worldwide (Table 12). Hawksworth [727] examined the number of fungi recorded associated with vascular plants in the British Isles and considered that one third of the fungal records could be considered as not host-specific, resulting in 67% host-specificity. It is not surprising that palms show lower percentages of host-specificity than that considered by Hawksworth [727] for a temperate region. As May [775] suggested, fungi, like insects, may be more generalised with regard to hosts in tropical regions due to a greater diversity of tree species, which results in lower percentages of host-specificity. As tree diversity increases, individuals of a particular species become more sparsely distributed, which probably exerts a selective pressure on fungi to become less specialised in their host requirements [775]. Even so, although tropical fungi may tend to be less host-specific than their temperate counterparts, the extraordinarily rich mycota of tropical hosts are likely to ensure higher plant:fungus ratios [10]. Thus, considering that ca. 30% of host-specificity in N. fruticans is likely to be higher as research continues to reveal new species (especially with the introduction of DNA sequence-based identifications), and that the worldwide prediction of fungal diversity is based on two/thirds host-specificity, it can be considered that host-specificity in palms inhabiting highly specialised habitats may have an intermediate value. It can be assumed that an average host-specificity value of ca. 49% occurs in palm trees inhabiting highly specialised habitats. In these cases, a plant:fungus ratio of 1:55 can be hypothesised, which would imply the existence of 143,000 species of palm fungi worldwide (Table 12). Thus, an average value of 126,100 species of palm fungi can be assumed for palms inhabiting highly specialised habitats.
Table 12.
Estimates of the total number of species of palm fungi in the world derived by different methods (see Section 5 for further explanation).
Table 12.
Estimates of the total number of species of palm fungi in the world derived by different methods (see Section 5 for further explanation).
| Benchmark | Basis | Reference and Reasoning | Plant:Fungus Ratio 1 | Estimate of Total Species Number 2 |
|---|---|---|---|---|
| I | Plant:fungus ratios in tropical palms | Hyde [762,763] based on extensive work on palm fungi in Australia | 1:28 | 67,600 |
| Fröhlich and Hyde [10] based on survey of fungi associated with six Licuala palms in Australia and Brunei Darussalam | 1:33 | 85,800 | ||
| II | Plant:fungus ratios in temperate palms | Hawksworth and Lücking [745] based on long-term investigations mainly on fungal collections from temperate regions | 1:9.8 | 25,480 |
| Present study based on the estimates presented by Taylor et al. [12] for the number of host-specific fungi in tropical and temperate palms growing inside and outside their natural geographic range | 1:8.7 | 22,620 | ||
| III | Plant:fungus ratios in palms inhabiting highly specialised habitats | Present study considering that 42 of the 142 fungal species recorded on Nypa fruticans are likely to be host-specific | 1:42 | 109,200 |
| Present study considering that palms inhabiting highly specialised habitats may have a higher percentage of host-specific fungi than typical tropical palms (25%) and be closer to the percentage of host-specificity estimated for temperate hosts (63%) | 1:55 | 143,000 | ||
| Mean I–III | 75,617 |
1 Plant:fungus ratio 1:52 for benchmark III was calculated considering the estimates presented by Hyde [762,763] for the number of fungal taxa (ca. 113) expected to develop on each palm species. 2 The estimate of the total number of fungal species was calculated considering that, according to the most recent literature available, palm trees comprise around 2600 species [1].
Based on the above-mentioned estimates, an average of benchmarks I to III yields a figure of 75,617 species of palm fungi worldwide (Table 12). However, some considerations should be made before further predictions, considering the estimation of almost 76,000 species of fungi on palms worldwide, which is remarkably close to the previous prediction of 73,000 by Taylor and Hyde [8]. The benchmarks presented are not comparable and represent different means of evaluating or predicting the same information, i.e., plant:fungus ratios in palm trees, given that plant:fungus ratios are one of the most important factors used in estimating global numbers of fungi [764]. A wide range of variation has been considered, which attempts to mimic the wide variation in habitats exploited by palms and the intimate relationship that palm fungi establish with their hosts. However, the figure of 76,000 species of fungi on palms worldwide is considered conservative and its accuracy is yet to be determined. Some considerations are discussed below.
An estimate based on an unweighted average. A determining factor for this scenario is the fact that all the benchmarks considered have the same weight when calculating the average value. However, most palm trees are native to tropical and subtropical regions, so it can be predicted that a plant:fungus ratio calculated on tropical palms (benchmarks I and III) may reveal a more realistic scenario than the one calculated on temperate palms (benchmark II). In fact, only about 130 of the 2600 palm species (ca. 5%) occur naturally beyond the tropical latitudes and would have a plant:fungus ratio like those calculated for temperate hosts [776]. Moreover, more than 90% of Arecaceae species diversity is restricted to tropical rainforests and part of the remaining 10% inhabit seasonal tropical and subtropical vegetation [777].
The overlooked fungal biodiversity. In most studies on palm fungi, particularly those that estimate plant:fungus ratios, the biodiversity of several groups of fungi are omitted from the surveys. These include fungi growing on tissues that are usually not examined, such as fungi from below-ground or reproductive organs, as well as lichen-forming fungi, whose occurrence is almost unknown on palm trees. In addition, many other species of fungi are often not recovered from samples, such as fastidious fungi, which can only be isolated using selective media (for detailed discussion on biodiversity of fungi omitted in such studies see [10]). Similarly, most studies are often biased towards the isolation or collection of ascomycetes or asexual morphs. As a consequence, certain taxonomic groups are overlooked, such as basidiomycetes, which are clearly poorly studied on palms (see Section 3).
The lack of data on certain palm fungal communities. Although considered conservative, the almost 76,000 species of fungi on palms worldwide may not be far from the truth, since the downward factors can be accommodated in benchmark III, which predicts a much higher plant:fungus ratio for some palms that inhabit hyperdiverse tropical ecosystems. However, the contribution of this benchmark is less detailed and supported by actual data than benchmarks I and II. Therefore, its contribution as a fine-tuning factor to the accuracy of the estimate may not be as reliable as the contribution of benchmarks I and II. This exceptionally high plant:fungus ratio is not expected to be found in most palms, as it is likely to be influenced by the unique abiotic conditions of the highly specialised habitats that are colonised by few palm species.
The lack of molecular-based studies. The vast majority of ecological studies on palm fungi have been based on morphological analyses. Since morphological characters are known to be an inadequate approach to identifying fungi due to phenotypic plasticity, the true diversity of documented palm fungal communities is likely to be underestimated. In addition, to date, almost no study has explored the diversity of palm fungi using palm samples for studies based on DNA metabarcoding technology via HTS. For instance, recent studies on the endophyte communities inhabiting the leaflets of mule palms (×Butiagrus nabonnandii), sampled using culture-dependent (CD) and culture-independent (CI) methods, have shown a small overlap in endophyte composition, with CI methods providing a higher estimate of species richness and composition [778]. Since palm fungal communities have proven to be extremely diverse, the assessment of environmental DNA in palm samples would probably give a more accurate idea of their true diversity and how far from the truth current predictions are. Likewise, given their diverse nature, palm fungal communities assessed using DNA metabarcoding technology could provide important new insights into the current discussion on how to formally describe “dark taxa”.
Predicting fungal diversity in palms is of great significance for estimates of the total number of fungi worldwide. In the present review, it is conservatively estimated that ca. 76,000 species of fungi can be found on palm trees worldwide, of which only just over 3% (ca. 2500 species) have been documented. This results in a total of ca. 97% of palm fungi awaiting to be documented. Considering that ca. 75% of all fungi collected on palms are new to science, this means that prioritising palm trees as host plants for fungal collections could reveal more than 55,000 new taxa to science. Therefore, given that the most currently accepted estimate of fungal species richness is between 2.2 and 3.8 million, ca. 2.5 to 1.5%, respectively, of the world’s unknown mycota could potentially be found on palm hosts.
6. Conclusions and Future Perspectives
Research carried out over the last 30 years suggests that Arecaceae hosts can be regarded as model plants for the study of fungal biodiversity. Palm tissues seem to support a vast and diverse mycobiota that can address several questions in biodiversity studies, which can be pointed out as actual fungal biodiversity challenges. Studies in Southeast Asia and Australasia have shown that there is a hyperdiverse group of fungi, referred to as palm fungi, consistently associated with palm trees in the tropics. A number of ecological issues were assessed in palm fungi, which are critical to the study of fungal communities and their biological patterns in ecosystems. These studies have emphasised the intimate relationship of palm fungi with palm hosts, insomuch as they are considered to be good biogeographical indicators, playing a key role in biodiversity surveys.
Palm fungi are considered a unique group of fungi, since many genera found to be associated with palms are host-specific or are rarely found associated with other plants. This host-specificity is far from being fully understood. Studies have revealed that it may be associated with the ecology of these microorganisms and their close association with palm hosts. In fact, some palm fungi recorded as saprobes are often recovered as endophytes, which can justify the high plant:fungus ratios estimated in Arecaceae hosts. In turn, this surely accounts for the remarkable number of new species to science that are continuously being described from palms.
The currently accepted estimate of the world’s mycota is between 2.2 and 3.8 million fungal species, yet less than 10% of them have been named so far. For this reason, the question “where are the missing fungi?” has often been asked and has motivated the persistent search for new fungal species. Evidence gained from the extensive investigation on palm fungi undoubtedly indicates that many of the missing fungi can be found on palms. In the present review, it has been conservatively estimated that more than 55,000 new taxa are expected to be found in palm collections. This means that approximately 1.5 to 2.5% of the world’s unknown mycota could potentially be found on palm hosts. Although host-specificity can be an important factor in estimating the number of palm fungal species worldwide, it is likely that conclusions are being drawn from data that are somewhat biased towards fungi, hosts, and substrata that are of human interest. This has been well illustrated in studies on palm fungi, which have largely focused on palms of international economic interest.
The estimates provided here highlight that palm fungi are an understudied assemblage. There is an enormous wealth of undiscovered and untapped palm fungi that could hold substantial potential for mankind. Therefore, the search for the undescribed palm fungi (and fungi in general) and the study of their diversity patterns are of the utmost importance in biodiversity studies. These studies have incredible economic potential in discovering microorganisms with new biotechnological and industrial uses. However, only a more complete inventory of these microorganisms will make it possible to preserve a representative collection for future research, society, and prosperity. Furthermore, this will only be possible with the development of appropriate protocols and methods to detect and understand this diversity, which will largely depend on the use of molecular data.
DNA sequence-based studies have revealed numerous additional cryptic taxa in well-known and established species and genera, suggesting that fungal biodiversity estimates may be highly underestimated. Most studies on palm fungi have been based on morphological analyses. As a result, most of the 2500 species of palm fungi have no associated molecular data. In order to successfully study and understand these fungi, a major investment is needed in their re-collection and epitypification. Only phylogenetic studies will be able to clarify the taxonomic structure of palm fungi and fill the current gaps in their knowledge. In addition, molecular analyses, including DNA metabarcoding, could provide missing links to palm fungal communities and therefore help to understand their population dynamics, such as host-specificity and biogeographical distribution.
Author Contributions
Conceptualization: A.J.L.P. and D.S.P.; Data curation: D.S.P.; Writing—original draft: D.S.P.; Writing—review and editing: A.J.L.P. and D.S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Portuguese Foundation for Science and Technology (FCT/MCTES) that finances BioISI (Biosystems and Integrative Sciences Institute) (UIDB/04046/2020 + UIDP/04046/2020) through national funds (OE) and the PhD grant to Diana S. Pereira (SFRH/BD/09742/2020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Study does not involve any huge data acquisition and the corresponding authors may be contacted for further assistance of the subjects discussed.
Acknowledgments
D.S.P. gratefully acknowledges the kind assistance of Lisa A. Castlebury of the United States Department of Agriculture (USDA)|Agricultural Research Service (ARS) in accessing information on fungus–host associations in the US National Fungus Collections Fungus–Host Database.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Baker, W.J.; Dransfield, J. Beyond Genera Palmarum: Progress and prospects in palm systematics. Bot. J. Linn. Soc. 2016, 182, 207–233. [Google Scholar] [CrossRef]
- Tomlinson, P.B. Systematics and Ecology of the Palmae. Annu. Rev. Ecol. Evol. Syst. 1979, 10, 85–107. [Google Scholar] [CrossRef]
- Johnson, D. Palms: Their Conservation and Sustained Utilization. Status Survey and Conservation Action Plan; IUCN: Gland, Switzerland, 1996; Available online: https://portals.iucn.org/library/node/7027 (accessed on 15 September 2023).
- Johnson, D.V. Non-Wood Forest Products 10/Rev.1—Tropical Palms 2010 Revision; FAO: Rome, Italy, 2011; Available online: https://www.fao.org/3/i1971e/i1971e00.htm (accessed on 15 September 2023).
- Dransfield, J.; Uhl, N.W.; Asmussen, C.B.; Baker, W.J.; Harley, M.M.; Lewis, C.E. Genera Palmarum. The Evolution and Classification of the Palms, 2nd ed.; Royal Botanic Gardens, Kew: London, UK, 2008. [Google Scholar]
- Fröhlich, J.; Hyde, K.D. Palm microfungi. Fung. Divers. Res. Ser. 2000, 3, 1–393. [Google Scholar]
- Hyde, K.D.; Alias, S.A. Biodiversity and distribution of fungi associated with decomposing Nypa fruticans. Biodivers. Conserv. 2000, 9, 393–402. [Google Scholar] [CrossRef]
- Taylor, J.E.; Hyde, K.D. Microfungi of tropical and temperate palms. Fungal Divers. Res. Ser. 2003, 12, 1–495. [Google Scholar]
- Zhang, S.-N.; Hyde, K.D.; Jones, E.B.G.; Jeewon, R.; Cheewangkoon, R.; Liu, J.-K. Striatiguttulaceae, a new pleosporalean family to accommodate Longicorpus and Striatiguttula gen. nov. from palms. Mycokeys 2019, 49, 99–129. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, J.; Hyde, K.D. Biodiversity of palm fungi in the tropics: Are global Fungal Divers. estimates realistic? Biodivers. Conserv. 1999, 8, 977–1004. [Google Scholar] [CrossRef]
- Fröhlich, J.; Hyde, K.D.; Petrini, O. Endophytic fungi associated with palms. Mycol. Res. 2000, 104, 1202–1212. [Google Scholar] [CrossRef]
- Taylor, J.E.; Hyde, K.D.; Jones, E.B.G. The biogeographical distribution of microfungi associated with three palm species from tropical and temperate habitats. J. Biogeogr. 2000, 27, 297–310. [Google Scholar] [CrossRef]
- Yanna; Ho, W.H.; Hyde, K.D. Fungal communities on decaying palm fronds in Australia, Brunei, and Hong Kong. Mycol. Res. 2001, 105, 1458–1471. [Google Scholar] [CrossRef]
- Yanna; Ho, W.H.; Hyde, K.D.; Goh, T.-K. Occurrence of fungi on tissues of Livistona chinensis. Fungal Divers. 2001, 6, 167–179. [Google Scholar]
- Yanna; Hyde, K.D. New saprobic fungi on fronds of palms from northern Queensland, Australia. Aust. Syst. Bot. 2002, 15, 755–764. [Google Scholar] [CrossRef]
- Pinnoi, A.; Phongpaichit, S.; Hyde, K.D.; Jones, E.B.G. Biodiversity of fungi on Calamus (Palmae) in Thailand. Cryptogam. Mycol. 2009, 30, 181–190. [Google Scholar]
- Hyde, K.D.; Fröhlich, J.; Taylor, J.E. Diversity of Ascomycetes on palms in the tropics. In Biodiversity of Tropical Microfungi; Hyde, K.D., Ed.; Hong Kong University Press: Hong Kong, China, 1997; pp. 141–156. [Google Scholar]
- Penzig, A.J.O.; Saccardo, P.A. Diagnoses Fungorum Novorum in Insula Java Collectorum; Tipografia di Angelo Ciminago: Genova, Italy, 1897; Series Secunda; Volume 11, pp. 491–530. [Google Scholar]
- Hennings, P. Fungi blumenaviensis. II. a cl. Alfr. Möller lecti. Hedwigia 1902, 41, 1–33. [Google Scholar]
- Hennings, P. Fungi Amazonici II. a cl. Ernesto Ule collecti. Hedwigia 1904, 43, 242–273. [Google Scholar]
- Hennings, P. Fungi Philippenses I. Hedwigia 1908, 47, 250–265. [Google Scholar]
- Rehm, H. Ascomycetes Philippinensis, II. Philipp. J. Sci. Section C. Botany 1913, 8, 251–263. [Google Scholar]
- Rehm, H. Ascomycetes Philippinensis, III. Philipp. J. Sci. Section C. Botany 1913, 8, 391–405. [Google Scholar]
- Rehm, H. Ascomycetes Philippinensis, V. Leafl. Philipp. Bot. 1914, 6, 2191–2237. [Google Scholar]
- Rehm, H. Ascomycetes Philippinensis, VIII. Leafl. Philipp. Bot. 1916, 8, 2935–2961. [Google Scholar]
- Spegazzini, C. Fungi nonnulli senegalenses et canariensis. Anales Mus. Nac. Hist. Nat. Buenos Aires 1914, 26, 117–134. [Google Scholar]
- Sydow, H.; Sydow, P. Beitrag zur Kenntnis der Pilzflora der Philppinen–Inseln. Ann. Mycol. 1917, 15, 12–268. [Google Scholar]
- Ellis, M.B. Haplobasidion, Lacellinopsis and Lacellina. Mycol. Pap. 1957, 67, 1–15. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. I. Mycol. Pap. 1960, 76, 1–36. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. II. Mycol. Pap. 1961, 79, 1–23. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes. VIII. Periconiella, Trichodochium, etc. Mycol. Pap. 1967, 111, 1–46. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. XI. Mycol. Pap. 1972, 131, 1–25. [Google Scholar]
- Deighton, F.C. Three leaf–spotting hyphomycetes on palms. Trans. Br. Mycol. Soc. 1985, 85, 739–742. [Google Scholar] [CrossRef]
- Deighton, F.C. Pseudocercospora carpentariae sp. nov. Trans. Br. Mycol. Soc. 1987, 89, 402–404. [Google Scholar] [CrossRef]
- Rogers, J.D.; Callan, B.E.; Samuels, G.J. The Xylariaceae of the rainforests of North Sulawesi (Indonesia). Mycotaxon 1987, 29, 118–172. [Google Scholar]
- Reid, D.A. Fungi venezuelani: V: The Cyphellaceae of Venezuela. Kew Bull. 1961, 15, 261–275. [Google Scholar] [CrossRef]
- Dennis, R.W.G. Fungi Venezuelani: VII. Kew Bull. 1965, 19, 231–273. [Google Scholar] [CrossRef]
- Müller, E.; Dennis, R.W.G. Fungi venezuelani: VIII: Plectascales, Sphaeriales, Loculoascomycetes. Kew Bull. 1965, 19, 357–386. [Google Scholar] [CrossRef]
- Wright, J.E. Agaricostilbum, a new genus of Deuteromycetes on palm spathes from Argentina. Mycologia 1970, 62, 679–682. [Google Scholar] [CrossRef]
- Hino, I.; Katumoto, K. Notes on fungi from western Japan (1). Bull. Fac. Agric. Yamaguchi Univ. 1956, 7, 257–266. [Google Scholar]
- Katumoto, K. Notes on fungi from western Japan (5). J. Jap. Bot. 1962, 37, 295–299. [Google Scholar] [CrossRef]
- Katumoto, K. Notes on fungi from western Japan (9). J. Jap. Bot. 1966, 41, 329–334. [Google Scholar] [CrossRef]
- Goos, R.D. Some Helicosporous fungi from Hawaii. Mycologia 1980, 72, 595–610. [Google Scholar] [CrossRef]
- Holubová-Jechová, V. Craspedodidymum, new genus of phialosporous Hyphomycetes. Czech Mycol. 1972, 26, 70–73. [Google Scholar]
- Liu, X.J.; Liao, Y.Z. Records on some species of genus Prathigada and Stenella. Acta Microbiol. Sin. 1980, 20, 116–121. [Google Scholar]
- Samuels, G.J.; Rossman, A.Y. Studies in the Amphisphaeriaceae (sensu lato) 2. Leiosphaerella cocoes and two new species of Oxydothis on palms. Mycotaxon 1987, 28, 461–471. [Google Scholar]
- Hughes, S.J. New Zealand Fungi 1. Ceratosporium Schw. N. Z. J. Bot. 1964, 2, 305–309. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand fungi 5. Trichothallus and Plokamidomyces states of Trichopeltheca. N. Z. J. Bot. 1965, 3, 320–332. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand fungi 6. Sporoschisma Berk. and Br. N. Z. J. Bot. 1966, 4, 77–85. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand fungi 7. Capnocybe and Capnophialophora, new form genera of sooty moulds. N. Z. J. Bot. 1966, 4, 333–353. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand fungi 13. Trichocladium Harz. N. Z. J. Bot. 1969, 7, 153–157. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand Fungi 25. Miscellaneous species. N. Z. J. Bot. 1978, 16, 311–370. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand Fungi 31. Capnobotrys, an anamorph of Metacapnodiaceae. N. Z. J. Bot. 1981, 19, 193–226. [Google Scholar] [CrossRef]
- McKenzie, E.H.C.; Buchanan, P.K.; Johnston, P.R. Checklist of fungi on nikau palm (Rhopalostylis sapida and R. baueri var. cheesemanii) in New Zealand. N. Z. J. Bot. 2004, 42, 335–355. [Google Scholar] [CrossRef]
- Pirozynski, K.A. Microfungi of Tanzania. I. Miscellaneous Fungi on oil palm. II. New hyphomycetes. Mycol. Pap. 1972, 129, 1–64. [Google Scholar]
- Matsushima, T. Microfungi of the Solomon Islands and Papua-New Guinea; Matsushima: Kobe, Japan, 1971. [Google Scholar]
- Matsushima, T. Icones Microfungorum a Matsushima Lectorum; Matsushima: Kobe, Japan, 1975. [Google Scholar]
- Matsushima, T. Saprophytic microfungi from Taiwan, part 1. Hyphomycetes. Matsushima Mycol. Mem. 1980, 1, 1–82. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 2. Matsushima Mycol. Mem. 1981, 2, 1–68. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 3. Matsushima Mycol. Mem. 1983, 3, 1–90. [Google Scholar] [CrossRef]
- Matsushima, T. Matsushima Mycological Memoirs 4. Matsushima Mycol. Mem. 1985, 4, 1–68. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 5. Matsushima Mycol. Mem. 1987, 5, 1–100. [Google Scholar] [CrossRef]
- Matsushima, T. Matsushima Mycological Memoirs 6. Matsushima Mycol. Mem. 1989, 6, 1–100. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 7. Matsushima Mycol. Mem. 1993, 7, 1–141. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 8. Matsushima Mycol. Mem. 1995, 8, 1–44. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 9. Matsushima Mycol. Mem. 1996, 9, 1–30. [Google Scholar]
- Matsushima, T. Matsushima Mycological Memoirs 10. Matsushima Mycol. Mem. 2003, 10, 1–214. [Google Scholar]
- Matsushima, K.; Matsushima, T. Fragmenta Mycologica I. Matsushima Mycol. Mem. 1995, 8, 45–54. [Google Scholar]
- Matsushima, K.; Matsushima, T. Fragmenta Mycologica II. Matsushima Mycol. Mem. 1996, 9, 31–40. [Google Scholar]
- Mercado-Sierra, A. El género Phragmospathula (Hyphomycetes: Fungi imperfecti) en Cuba. Acta Bot. Cub. 1980, 5, 1–6. [Google Scholar]
- Mercado-Sierra, A. Lista preliminar de hifomicetes demaciáceos de la Estación Ecológica de Sierra del Rosario y zonas adyacentes. Acta Bot. Cub. 1981, 6, 1–6. [Google Scholar]
- Mercado-Sierra, A. Taxonomía y aspectos ecológicos de algunos hifomicetes helicospóricos hallados en Cuba. Acta Bot. Cub. 1982, 11, 1–11. [Google Scholar]
- Mercado-Sierra, A. La palma real (Roystonea regia): Un sustrato idóneo para el desarrollo de hifomicetes demaciáceos. Acta Bot. Cub. 1983, 15, 1–13. [Google Scholar]
- Holubová-Jechová, V. New or interesting phialidic hyphomycetes from Cuba. Mycotaxon 1982, 15, 277–292. [Google Scholar]
- Holubová-Jechová, V.; Mercado-Sierra, A. Some new or interesting microfungi from Cuba. Mycotaxon 1982, 14, 309–315. [Google Scholar]
- Holubová-Jechová, V.; Mercado-Sierra, A. Hyphomycetes from Loma de la Coca and some localities of La Habana and Matanzas provinces, Cuba. Acta Bot. Cub. 1989, 76, 1–15. [Google Scholar]
- Mercado-Sierra, A.; Castañeda-Ruíz, R.F. Nueva especie de Triadelphia (Hyphomycetes, Deuteromycotina) de Cuba. Revista Jard. Bot. Nac. Univ. Habana 1983, 4, 65–79. [Google Scholar]
- Mercado-Sierra, A.; Castañeda-Ruíz, R.F. Nuevos hifomicetes tálicos de Cuba. Acta Bot. Cub. 1985, 32, 1–10. [Google Scholar]
- Castañeda-Ruíz, R.F.; Arnold, G.R.W. Deuteromycotina de Cuba. I. Hyphomycetes. Revista Jard. Bot. Nac. Univ. Habana 1985, 6, 47–67. [Google Scholar]
- Castañeda-Ruíz, R.F.; Arnold, G.R.W. Algunos hongos nuevos para Cuba. Revista Jard. Bot. Nac. Univ. Habana 1985, 6, 55–56. [Google Scholar]
- Mercado-Sierra, A.; Holubová-Jechová, V.; Mena-Portales, J.; Fraginals, G.G. Hongos imperfectos de Pinar del Río, Cuba: El ambiente y la taxonomía de hifomicetes demaciáceos hallados. Rep. Investig. Inst. Ecol. Sist. Acad. Ci. Cuba 1987, 2, 1–10. [Google Scholar]
- Mercado-Sierra, A.; Holubová-Jechová, V.; Mena-Portales, J. Estudios sobre la microflora de Cuba: Hifomicetes con tretoconidios. Rep. Investig. Inst. Ecol. Sist. Acad. Ci. Cuba 1989, 4, 1–8. [Google Scholar]
- Mercado-Sierra, A.; Mena-Portales, J. Hifomicetes de Topes de Collantes, Cuba I. Especies holoblásticas. Acta Bot. Hung. 1986, 32, 189–205. [Google Scholar]
- Mena-Portales, J.; Mercado-Sierra, A. Hifomicetes de Topes de Collantes, Cuba II. Especies enteroblásticas. Acta Bot. Hung. 1987, 33, 75–79. [Google Scholar]
- Mena-Portales, J.; Mercado-Sierra, A. Algunos hifomicetes de las provincias Ciudad de La Habana y La Habana, Cuba. Rep. Investig. Inst. Ecol. Sist. Acad. Ci. Cuba 1987, 17, 1–16. [Google Scholar]
- Mercado-Sierra, A. Hifomicetes demaciáceos de Cuba (1). Acta Bot. Cub. 1980, 1, 1–5. [Google Scholar]
- Mercado-Sierra, A. Hifomicetes demaciáceos de Cuba (2). Acta Bot. Cub. 1982, 14, 1–7. [Google Scholar]
- Holubová-Jechová, V. Studies on Hyphomycetes from Cuba I. Czech Mycol. 1983, 37, 12–18. [Google Scholar]
- Holubová-Jechová, V. Studies on hyphomycetes from Cuba V. Six new species of dematiaceous hyphomycetes from Havana Province. Czech Mycol. 1987, 41, 29–36. [Google Scholar]
- Holubová-Jechová, V. Studies on hyphomycetes from Cuba VI. New and rare species with tretic and phialidic conidiogenous cells. Czech Mycol. 1987, 41, 107–114. [Google Scholar]
- Holubová-Jechová, V. Studies on hyphomycetes from Cuba VII. Seven new taxa of dematiaceous hyphomycetes. Czech Mycol. 1988, 42, 23–30. [Google Scholar]
- Holubová-Jechová, V. Studies on hyphomycetes from Cuba VIII. A new genus Piricaudilium and some species new for the territory of Cuba. Czech Mycol. 1988, 42, 200–204. [Google Scholar]
- Holubová-Jechová, V.; Mercado-Sierra, A. Studies on Hyphomycetes from Cuba II. Hyphomycetes from the Isla de la Juventud. Czech Mycol. 1984, 38, 96–120. [Google Scholar]
- Holubová-Jechová, V.; Mercado-Sierra, A. Studies on hyphomycetes from Cuba IV. Dematiaceous hyphomycetes from the province Pinar del Río. Czech Mycol. 1986, 40, 142–164. [Google Scholar]
- Holubová-Jechová, V.; Castañeda-Ruíz, R.F. Studies on hyphomycetes from Cuba III. New and interesting dematiaceous taxa from leaf litter. Czech Mycol. 1986, 40, 74–85. [Google Scholar]
- Mena-Portales, J.; Mercado-Sierra, A. Nuevos o raros hifomicetes de Cuba III. Phragmospathulella. Un nuevo género trético. Revista Jard. Bot. Nac. Univ. Habana 1986, 7, 31–34. [Google Scholar]
- Mena-Portales, J.; Mercado-Sierra, A. Nuevos o raros hifomicetes de Cuba. IV. Un nuevo género lignícola con conidiogénesis trética. Acta Bot. Cub. 1988, 54, 1–6. [Google Scholar]
- Mercado-Sierra, A.; Castañeda-Ruíz, R.F. Nuevos o raros hifomicetes de Cuba. I. Especies de Cacumisporium, Guedea, Rhinocladium y Veronaea. Acta Bot. Cub. 1987, 50, 1–7. [Google Scholar]
- Mercado-Sierra, A.; Mena-Portales, J. Nuevos o raros hifomicetes de Cuba. II. Un nuevo género sobre Roystonea regia. Acta Bot. Cub. 1988, 53, 1–5. [Google Scholar]
- Mercado-Sierra, A.; Mena-Portales, J. Nuevos o raros hifomicetes de Cuba. V. Especies de Stachybotrys. Acta Bot. Cub. 1988, 55, 1–8. [Google Scholar]
- Mercado-Sierra, A.; Mena-Portales, J. Nuevos o raros hifomicetes de Cuba. VI. Neosporidesmium, nuevo género sinemático. Acta Bot. Cub. 1988, 59, 1–6. [Google Scholar]
- Mercado-Sierra, A.; Mena-Portales, J. Nuevos o raros hifomicetes de Cuba VII. Especies enteroblásticas. Acta Bot. Hung. 1992, 37, 63–73. [Google Scholar]
- Mercado-Sierra, A. Hifomicetes Demaciáceos de Sierra del Rosario, Cuba; Editorial Academia: La Habana, Cuba, 1984. [Google Scholar]
- Hyde, K.D.; Taylor, J.E.; Fröhlich, J. Genera of ascomycetes from palms. Fung. Divers. Res. Ser. 2000, 2, 1–247. [Google Scholar]
- Hyde, K.D. Fungi from palms. I. The genus Linocarpon, a revision. Sydowia 1992, 44, 32–54. [Google Scholar]
- Hyde, K.D. Fungi from palms. II. Kirschsteiniothelia aethiops from the date palm Phoenix dactylifera. Sydowia 1992, 45, 1–4. [Google Scholar]
- Hyde, K.D. Fungi from palms. III. The genus Pemphidium Montagne (Ascomycotina). Sydowia 1993, 45, 5–14. [Google Scholar]
- Hyde, K.D. Fungi from palms. IV. Palmicola archontophoenicis gen. et sp. nov. Sydowia 1993, 45, 15–20. [Google Scholar]
- Hyde, K.D. Fungi from palms. IX. Manokwaria notabilis gen. et sp. nov. (Ascomycetes) from Irian Jaya and Australia. Sydowia 1993, 45, 246–251. [Google Scholar]
- Hyde, K.D. Fungi from palms. V. Phomatospora nypae sp. nov. and notes on marine fungi from Nypa fruticans in Malaysia. Sydowia 1993, 45, 199–203. [Google Scholar]
- Hyde, K.D. Fungi from palms. VI. Reflections on Oxydothis and related genera. Sydowia 1993, 45, 204–225. [Google Scholar]
- Hyde, K.D. Fungi from palms. VII. The genus Oxydothis from rachides of palms in north Queensland, including five new species. Sydowia 1993, 45, 226–240. [Google Scholar]
- Hyde, K.D. Fungi from palms. VIII. The genus Myelosperma (Ascomycotina). Sydowia 1993, 45, 241–245. [Google Scholar]
- Hyde, K.D. Fungi from palms. X. Lockerbia palmicola, a new cleistothecial genus in the Sordariales. Sydowia 1993, 46, 23–28. [Google Scholar]
- Hyde, K.D. Fungi from palms. XI. Appendispora frondicola gen. et sp. nov. from Oncosperma horridum in Brunei. Sydowia 1994, 46, 29–34. [Google Scholar]
- Hyde, K.D. Fungi from palms. XII. Three new intertidal ascomycetes from palm fronds. Sydowia 1994, 46, 257–264. [Google Scholar]
- Hyde, K.D. Fungi from palms. XIII. The genus Oxydothis, a revision. Sydowia 1994, 46, 265–314. [Google Scholar]
- Hyde, K.D. Fungi from palms. XIV. Durispora elaeidicola gen. et sp. nov. Sydowia 1994, 46, 315–320. [Google Scholar]
- Fröhlich, J.; Hyde, K.D. Fungi from palms. XIX. Caudatispora palmicola gen. et sp. nov. in Ecuador. Sydowia 1995, 47, 38–43. [Google Scholar]
- Lu, B.-S.; Hyde, K.D. Fungi from palms. XLI. Fasciatispora sabalicola: Further collections from Florida, USA. Mycotaxon 1999, 71, 393–397. [Google Scholar]
- Fröhlich, J.; Lowen, R.; Hyde, K.D. Fungi from palms. XLV. Nectriella erythroclypea sp. nov. (Bionectriaceae, Hypocreales). Nova Hedwig. 2000, 70, 425–430. [Google Scholar] [CrossRef]
- Hyde, K.D. Fungi from palms. XV. Sabalicola gen. nov., and a new combination for Anthostomella sabalensioides. Nova Hedwig. 1995, 60, 595–598. [Google Scholar]
- Hyde, K.D. Fungi from palms. XVI. Cocoicola gen. nov. Nova Hedwig. 1995, 60, 599–604. [Google Scholar]
- Hyde, K.D. Fungi from palms. XVII. The genus Fasciatispora with notes on Amphisphaerella. Nova Hedwig. 1995, 61, 249–268. [Google Scholar]
- Hyde, K.D. Fungi from palms. XVIII. Appendicospora coryphae, a new name for Apiosporella coryphae. Sydowia 1995, 47, 31–37. [Google Scholar]
- Hyde, K.D. Fungi from palms. XX. The genus Guignardia. Sydowia 1995, 47, 180–198. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXI. The genus Seynesia. Sydowia 1995, 47, 199–212. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXII. A new species of Ascotaiwania. Sydowia 1995, 47, 213–216. [Google Scholar]
- Hyde, K.D.; Fröhlich, J. Fungi from palms. XXIII. Rachidicola gen. et sp. nov. Sydowia 1995, 47, 217–222. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXIV. The genus Bondiella. Mycotaxon 1996, 57, 347–352. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXIX. Arecophila gen. nov. (Amphisphaeriales, Ascomycota), with five new species and two new combinations. Nova Hedwig. 1996, 63, 81–100. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXV. Pestalosphaeria elaeidis. Mycotaxon 1996, 57, 353–357. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXVI. The genus Anthostomella, with ten new species. Nova Hedwig. 1996, 62, 273–340. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXVII. Capsulospora gen. nov., with three new species. Sydowia 1996, 48, 111–121. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXVIII. Two new species of Pemphidium from Australia and Indonesia. Sydowia 1996, 48, 122–130. [Google Scholar]
- Hyde, K.D.; Kang, J.C.; Kong, R.Y.C. Fungi from palms. XXX. Notes on Amphisphaeria species from palms and a description of Amphisphaeria umbrina. Nova Hedwig. 1996, 63, 101–108. [Google Scholar]
- Hyde, K.D.; Taylor, J.E. Fungi from palms. XXXI. The genus Nipicola (Ascomycetes, Xylariaceae) with one new species. Nova Hedwig. 1996, 63, 417–424. [Google Scholar]
- Hyde, K.D. Fungi from palms. XXXII. Arecomyces gen. nov., with seven new species. Sydowia 1996, 48, 224–240. [Google Scholar]
- Hyde, K.D.; Aptroot, A. Fungi from palms. XXXIII. The genus Massarina, with a new species. Nova Hedwig. 1997, 64, 491–504. [Google Scholar] [CrossRef]
- Hyde, K.D.; Taylor, J.E.; Fröhlich, J. Fungi from palms. XXXIV. The genus Neolinocarpon with five new species and one new combination. Fungal Divers. 1998, 1, 115–131. [Google Scholar]
- Fröhlich, J.; Hyde, K.D. Fungi from palms. XXXIX. Asymmetricospora gen. et sp. nov. (Melannomataceae). Sydowia 1998, 50, 182–186. [Google Scholar]
- Taylor, J.E.; Hyde, K.D.; Jones, E.B.G. Fungi from palms. XXXV. Thyridium chrysomallum associated with Archontophoenix alexandrae (Palmae) cultivated in Hong Kong. Sydowia 1997, 49, 94–100. [Google Scholar]
- Hyde, K.D.; Fröhlich, J.; Taylor, J.E. Fungi from palms. XXXVI. Reflections on unitunicate ascomycetes with apiospores. Sydowia 1998, 50, 21–80. [Google Scholar]
- Hyde, K.D.; Fröhlich, J. Fungi from palms. XXXVII. The genus Astrosphaeriella, including ten new species. Sydowia 1998, 50, 81–132. [Google Scholar]
- Fröhlich, J.; Hyde, K.D. Fungi from palms. XXXVIII. The genera Mycosphaerella and Sphaerella. Sydowia 1998, 50, 171–181. [Google Scholar]
- Taylor, J.E.; Hyde, K.D. Fungi from palms. XL. Iodosphaeria. Sydowia 1999, 51, 127–132. [Google Scholar]
- Hyde, K.D.; Aptroot, A.; Fröhlich, J.; Taylor, J.E. Fungi from palms. XLII. Didymosphaeria and similar ascomycetes from palms. Nova Hedwig. 1999, 69, 449–471. [Google Scholar] [CrossRef]
- Hyde, K.D.; Aptroot, A.; Fröhlich, J.; Taylor, J.E. Fungi from palms. XLIII. Lophiostoma and Astrosphaeriella species with slit–like ostioles. Nova Hedwig. 2000, 70, 143–160. [Google Scholar] [CrossRef]
- Aptroot, A.; Fröhlich, J.; Hyde, K.D. Fungi from palms. XLIV. Two new Massarina species with pigmented ostioles. Nova Hedwig. 2000, 70, 227–232. [Google Scholar] [CrossRef]
- Smith, G.J.D.; Hyde, K.D. Fungi from palms. XLIX. Astrocystis, Biscogniauxia, Cyanopulvis, Hypoxylon, Nemania, Guestia, Rosellinia and Stilbohypoxylon. Fungal Divers. 2001, 7, 89–127. [Google Scholar]
- Guo, L.D.; Hyde, K.D. Fungi from palms. XLVI. Seynesia livistonae sp nov (Xylariaceae) from Hong Kong. Nova Hedwig. 2001, 72, 461–465. [Google Scholar] [CrossRef]
- Hosagoudar, V.B.; Abraham, T.K.; Biju, C.K.; Hyde, K.D. Fungi from palms. XLVII. A new species of Asterina on palms from India. Fungal Divers. 2001, 6, 69–73. [Google Scholar]
- Sarma, V.V.; Hyde, K.D. Fungi from palms. XLVIII. Curvatispora singaporensis gen. et sp nov on Livistona spinosa from Singapore. Nova Hedwig. 2001, 72, 479–485. [Google Scholar] [CrossRef]
- Hyde, K.D. The genus Linocarpon from the mangrove palm Nypa fruticans. Trans. Mycol. Soc. Jpn. 1988, 29, 339–350. [Google Scholar]
- Hyde, K.D. Studies on the tropical marine fungi of Brunei. Bot. J. Linn. Soc. 1988, 98, 135–151. [Google Scholar] [CrossRef]
- Hyde, K.D.; Nakagiri, A. A new species of Oxydothis from the mangrove palm, Nypa fruticans. Trans. Mycol. Soc. Jpn. 1989, 30, 69–75. [Google Scholar]
- Hyde, K.D. Studies on the tropical marine fungi of Brunei. II. Notes on five interesting species. Trans. Mycol. Soc. Jpn. 1988, 29, 161–171. [Google Scholar]
- Hyde, K.D.; Jones, E.B.G. Marine mangrove fungi. Mar. Ecol. 1988, 9, 15–34. [Google Scholar] [CrossRef]
- Hyde, K.D. Intertidal mangrove fungi from north Sumatra. Can. J. Bot. 1989, 67, 3078–3082. [Google Scholar] [CrossRef]
- Hyde, K.D. Helicascus kanaloanus, Helicascus nypae sp. nov. and Salsuginea ramicola gen. et sp. nov. from intertidal mangrove wood. Bot. Mar. 1991, 34, 311–318. [Google Scholar] [CrossRef]
- Hyde, K.D. A new amphisphaeriaceous fungus from intertidal fronds of Nypa fruticans. Trans. Mycol. Soc. Jpn. 1991, 32, 265–271. [Google Scholar]
- Hyde, K.D. Fungi from decaying intertidal fronds of Nypa fruticans, including three new genera and four new species. Bot. J. Linn. Soc. 1992, 110, 95–110. [Google Scholar] [CrossRef]
- Hyde, K.D. Fungi from Nypa fruticans: Nipicola carbospora gen. et sp. nov. (Ascomycotina). Cryptogam. Bot. 1992, 2, 330–332. [Google Scholar]
- Hyde, K.D.; Sutton, B.C. Nypaella frondicola gen. et sp. nov., Plectophomella nypae sp. nov. and Pleurophomopsis nypae sp. nov. (Coelomycetes) from intertidal fronds of Nypa fruticans. Mycol. Res. 1992, 96, 210–214. [Google Scholar] [CrossRef]
- Hyde, K.D.; Alias, S.A. Linocarpon angustatum sp. nov., and Neolinocarpon nypicola sp. nov. from petioles of Nypa fruticans, and a list of fungi from aerial parts of this host. Mycoscience 1999, 40, 145–149. [Google Scholar] [CrossRef]
- Hyde, K.D.; Goh, T.-K.; Lu, B.-S.; Alias, S.A. Eleven new intertidal fungi from Nypa fruticans. Mycol. Res. 1999, 103, 1409–1422. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Hyde, K.D.; Read, S.J.; Moss, S.T.; Alias, S.A. Tirisporella gen nov, an ascomycete from the mangrove palm Nypa fruticans. Can. J. Bot. 1996, 74, 1487–1495. [Google Scholar] [CrossRef]
- Hyde, K.D.; Lee, S.Y. Ecology of mangrove fungi and their role in nutrient cycling: What gaps occur in our knowledge? Hydrobiologia 1995, 295, 107–118. [Google Scholar] [CrossRef]
- Hyde, K.D. The genus Saccardoella from intertidal mangrove wood. Mycologia 1992, 84, 803–810. [Google Scholar] [CrossRef]
- Hyde, K.D. Frondisphaeria palmicola gen. et sp. nov. from Brunei. Mycoscience 1996, 37, 169–171. [Google Scholar] [CrossRef]
- Hyde, K.D. The genus Roussoella, including two new species from palms in Cuyabeno, Ecuador. Mycol. Res. 1997, 101, 609–616. [Google Scholar] [CrossRef]
- Hyde, K.D. Additions to the genus Linocarpon (Ascomycetes: Hyponectriaceae). Bot. J. Linn. Soc. 1997, 123, 109–131. [Google Scholar] [CrossRef]
- Hyde, K.D.; Philemon, E. Capitorostrum cocoes sp. nov., causing leaf spot of Cocos nucifera. Mycotaxon 1991, 42, 95–97. [Google Scholar]
- Hyde, K.D. Aquatic fungi on rachides of Livistona in the Western Province of Papua New Guinea. Mycol. Res. 1994, 98, 719–725. [Google Scholar] [CrossRef]
- Hyde, K.D. Fungi from rachides of Livistona in the western province of Papua New Guinea. Bot. J. Linn. Soc. 1994, 116, 315–324. [Google Scholar] [CrossRef]
- Hyde, K.D.; Eriksson, O.E.; Yue, J.Z. Roussoella, an ascomycete genus of uncertain relationships with a Cytoplea anamorph. Mycol. Res. 1996, 100, 1522–1528. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cannon, P.F.; Barr, M.E. Phaeochoraceae, a new ascomycete family from palms. Syst. Ascomycetum 1997, 15, 117–120. [Google Scholar]
- Hyde, K.D.; Wong, S.-W. Ultrastructural studies on the Myelospermaceae fam. nov., with a new species of Myelosperma. Mycol. Res. 1999, 103, 347–352. [Google Scholar] [CrossRef]
- Hyde, K.D.; Fröhlich, J. Nigramammilla calami gen. et sp nov and Arecomyces calami, A. licualae and Pseudohalonectria palmae spp. nov from palms. Cryptogam. Mycol. 2003, 24, 13–20. [Google Scholar]
- Hyde, K.D.; Fröhlich, J.; Taylor, J.E. Cocoicola livistonicola, sp. nov., and notes on Cocoicola cylindrospora from palms. Mycoscience 1997, 38, 255–258. [Google Scholar] [CrossRef]
- Hyde, K.D.; Goh, T.-K.; Taylor, J.E.; Fröhlich, J. Byssosphaeria, Chaetosphaeria, Niesslia and Ornatispora gen. nov., from palms. Mycol. Res. 1999, 103, 1423–1439. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. Melanographium palmicolum sp. nov. from Hong Kong, and a key to the genus. Mycol. Res. 1997, 101, 1097–1100. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. The generic distinction between Chaetopsina and Kionochaeta, with descriptions of two new species. Mycol. Res. 1997, 101, 1517–1523. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D.; Ho, W.H.; Yanna. A revision of the genus Dictyosporium, with descriptions of three new species. Fungal Divers. 1999, 2, 65–100. [Google Scholar]
- Taylor, J.E.; Fröhlich, J.; Hyde, K.D. Lasiosphaeria and a similar new genus from palms. Mycoscience 2001, 42, 369–377. [Google Scholar] [CrossRef]
- Lu, B.-S.; Hyde, K.D.; Ho, H.W.; Tsui, K.M.; Taylor, J.E.; Wong, K.M.; Yanna; Zhou, D. Checklist of Hong Kong fungi. Fung. Divers. Res. Ser. 2000, 5, 1–207. [Google Scholar]
- Fröhlich, J.; Hyde, K.D. Guignardia candeloflamma sp. nov. causing leaf spots of Pinanga spp. Mycol. Res. 1995, 99, 110–112. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. Spadicoides palmicola sp. nov. on Licuala sp. from Brunei, and a note on Spadicoides heterocolorata comb. nov. Can. J. Bot. 1998, 76, 1698–1702. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. Stratiphoromyces brunneisporus gen. et sp. nov., an undescribed dematiaceous hyphomycete on Licuala palms. Mycol. Res. 1998, 102, 1149–1152. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. Polybulbophiale palmicola gen. et sp. nov. (Hyphomycetes) from Brunei. Mycotaxon 1998, 69, 145–151. [Google Scholar]
- Tsui, C.K.M.; Leung, Y.M.; Hyde, K.D.; Hodgkiss, I.J. Three new Ophioceras species (Ascomycetes) from the tropics. Mycoscience 2001, 42, 321–326. [Google Scholar] [CrossRef]
- Phengsintham, P.; Hyde, K.D. Fungi of Laos I: Ascomycetes from Palms. In Proceedings of the Building Capacity in Biodiversity Information Sharing, 2003: Joint International Forum on Biodiversity Information, Building Capacity in Asia and Oceania, Tsukuba, Japan, 4–10 October 2003; National Institute for Environmental Studies: Tsukuba, Japan, 2003; pp. 174–183. [Google Scholar]
- Phengsintham, P.; Hyde, K.D. Check list of Lao fungi. In Proceedings of the Building Capacity in Biodiversity Information Sharing, 2003: Joint International Forum on Biodiversity Information, Building Capacity in Asia and Oceania, Tsukuba, Japan, 4–10 October 2003; National Institute for Environmental Studies: Tsukuba, Japan, 2003; pp. 184–190. [Google Scholar]
- Taylor, J.E.; Hyde, K.D.; Jones, E.B.G. Pulmosphaeria archontophoenicis gen. et sp. nov. associated with Archontophoenix alexandrae (Arecaceae) in Northern Queensland. Sydowia 1996, 48, 255–262. [Google Scholar]
- Frohlich, J.; Hyde, K.D. New Oxydothis species associated with palm leaf spots in North Queensland, Australia. Mycol. Res. 1994, 98, 213–218. [Google Scholar] [CrossRef]
- Frohlich, J.; Hyde, K.D. Astrosphaeriella fronsicola sp. nov. associated with leaf spots of Oraniopsis and other palms. Mycol. Res. 1995, 99, 453–456. [Google Scholar] [CrossRef]
- Frohlich, J.; Hyde, K.D. Maculatipalma fronsicola gen. et sp. nov. causing leaf spots on palm species in North Queensland with descriptions of related genera: Apioplagiostoma and Plagiostoma. Mycol. Res. 1995, 99, 727–734. [Google Scholar] [CrossRef]
- Hyde, K.D.; Frohlich, J. Mycosphaerella palmicola associated with leaf spots of Cocos nucifera in Australia, Irian Jaya and Papua New Guinea. Mycol. Res. 1995, 99, 704–706. [Google Scholar] [CrossRef]
- Frohlich, J.; Hyde, K.D.; Guest, D.I. Fungi associated with leaf spots of palms in North Queensland, Australia. Mycol. Res. 1997, 101, 721–732. [Google Scholar] [CrossRef]
- Hyde, K.D.; Taylor, J.E.; Fröhlich, J. Two new species of Pseudohalonectria from palms. Mycologia 1999, 91, 520–524. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. Delortia palmicola and two new species from wood submerged in a freshwater stream in Australia. Mycol. Res. 1997, 101, 42–46. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. A new species of Canalisporium from Australia. Mycologia 2000, 92, 589–592. [Google Scholar] [CrossRef]
- Hyde, K.D.; Goh, T.-K. Tropical Australian Freshwater Fungi XIII. A new species of Anthostomella and its sporodochial Geniculosporium anamorph. Nova Hedwig. 1998, 67, 225–233. [Google Scholar] [CrossRef]
- Taylor, J.E.; Hyde, K.D. Cannonia gen. nov., from palms in the Southern Hemisphere. Mycol. Res. 1999, 103, 1398–1402. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. Lepteutypa hexagonalis sp. nov. from Pinanga sp. in Ecuador. Mycol. Res. 1997, 101, 85–88. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. A new species of Palmicola from Ecuador. Mycol. Res. 1996, 100, 714–716. [Google Scholar] [CrossRef]
- Goh, T.-K.; Hyde, K.D. A new species of Nectria from Mauritia flexuosa (Arecaceae) in Ecuador and a key to Nectria and allied genera on palms. Mycoscience 1996, 37, 277–282. [Google Scholar] [CrossRef]
- Hyde, K.D.; Stanley, S.J.; Steinke, T.D. Fungi associated with leaf spots of palms. Maculatifrondis aequatoriensis gen. et sp. nov., with a Cyclodomus anamorph, and Myelosperma parasitica sp. nov. Mycol. Res. 1996, 100, 1509–1514. [Google Scholar] [CrossRef]
- Lu, B.-S.; Hyde, K.D.; Liew, E.C.Y. Eight new species of Anthostomella from South Africa. Mycol. Res. 2000, 104, 742–754. [Google Scholar] [CrossRef]
- Hyde, K.D.; Ho, W.-H.; Tsui, C.K.M. The genera Aniptodera, Halosarpheia, Nais and Phaeonectriella from freshwater habitats. Mycoscience 1999, 40, 165–183. [Google Scholar] [CrossRef]
- Wong, S.-W.; Hyde, K.D. Ultrastructural observations on Oxydothis alexandrarum. Fungal Divers. 1999, 2, 181–188. [Google Scholar]
- Hyde, K.D.; Cannon, P.F. Fungi causing tar spots on palms. Mycol. Pap. 1999, 175, 1–114. [Google Scholar]
- Barr, M.E.; Ohr, H.D.; Murphy, M.K. The genus Serenomyces on palms. Mycologia 1989, 81, 47–51. [Google Scholar] [CrossRef]
- Barr, M.E.; Ohr, H.D.; Ferrin, D.M.; Mundo-Ocampo, M. A new species of Serenomyces from date palm in California. Mycotaxon 1997, 61, 481–484. [Google Scholar]
- Taylor, J.E.; Hyde, K.D.; Jones, E.B.G. Endophytic fungi associated with the temperate palm, Trachycarpus fortunei, within and outside its natural geographic range. New Phytol. 1999, 142, 335–346. [Google Scholar] [CrossRef]
- Hyde, K.D.; Wong, S.-W. An ultrastructural study of the asci and banded ascospores of Fasciatispora petrakii. Fungal Divers. 1999, 2, 129–134. [Google Scholar]
- Wang, Y.-Z.; Hyde, K.D. Hyponectria buxi with notes on the Hyponectriaceae. Fungal Divers. 1999, 3, 159–172. [Google Scholar]
- Lu, B.-S.; Hyde, K.D.; Yuan, Z.Q. The genus Anthostomella in Australia. Fungal Divers. 1999, 3, 99–106. [Google Scholar]
- Lu, B.-S.; Hyde, K.D. A world monograph of Anthostomella. Fung. Divers. Res. Ser. 2000, 4, 1–376. [Google Scholar]
- Lu, B.-S.; Hyde, K.D. Anthostomella longa sp. nov. and note on other species on monocots from Hong Kong. Cryptogam. Mycol. 2000, 21, 207–214. [Google Scholar] [CrossRef]
- Lu, B.-S.; Hyde, K.D. Species of Anthostomella from Brunei, including A. oblongata sp. nov. Mycoscience 2000, 41, 223–226. [Google Scholar] [CrossRef]
- Aptroot, A. A monograph of Didymosphaeria. Stud. Mycol. 1995, 37, 1–160. [Google Scholar]
- Aptroot, A. Redisposition of some species excluded from Didymosphaeria (Ascomycotina). Nova Hedwig. 1995, 60, 325–379. [Google Scholar]
- Yanna; Hyde, K.D.; Goh, T.-K. Staurophoma calami, a new coelomycete from Hong Kong. Sydowia 1998, 50, 139–143. [Google Scholar]
- Yanna; Hyde, K.D.; Fröhlich, J. A new species of Appendicospora from Hong Kong. Mycoscience 1997, 38, 395–397. [Google Scholar] [CrossRef]
- Yanna; Hyde, K.D.; Goh, T.-K. Koorchaloma novojournalis sp. nov., a new sporodochial fungus from Hong Kong. Fungal Divers. 1998, 1, 193–197. [Google Scholar]
- Yanna; Hyde, K.D.; Goh, T.-K. Endomelanconium phoenicicola, a new coelomycete from Phoenix hanceana in Hong Kong. Fungal Divers. 1999, 2, 199–204. [Google Scholar]
- Yanna; Ho, W.H.; McKenzie, E.H.C.; Hyde, K.D. New saprobic fungi on palm fronds, including Brachysporiopsis gen. nov. Cryptogam. Mycol. 2004, 25, 129–135. [Google Scholar]
- Yanna; Ho, W.H.; Goh, T.-K.; Hyde, K.D. A new species of Everhartia associated with leaf spots of Phoenix hanceana from Hong Kong. Bot. J. Linn. Soc. 2000, 134, 465–470. [Google Scholar] [CrossRef]
- Yanna; Ho, W.H.; Hyde, K.D.; McKenzie, E.H.C. Sporidesmiella oraniopsis, a new species of dematiaceous hyphomycete from North Queensland, Australia and synopsis of the genus. Fungal Divers. 2001, 8, 183–190. [Google Scholar]
- Zhuang, W.-Y.; Hyde, K.D. New species of Lachnum and Perrotia from Hong Kong, China. Mycologia 2001, 93, 606–611. [Google Scholar] [CrossRef]
- Wong, M.K.M.; Yanna; Goh, T.-K.; McKenzie, E.H.C. Two new species of Constantinella from Hong Kong. Fungal Divers. 2001, 8, 173–181. [Google Scholar]
- Yu, Z.-H.; Zhuang, W.-Y. New taxa and new records of Lachnum and Arachnopeziza (Helotiales, Hyaloscyphaceae) from tropical China. Nova Hedwig. 2002, 74, 415–428. [Google Scholar] [CrossRef]
- Zhuang, W.-Y. A new species of Lachnum on leaves of Livistona and a key to the Chinese species of the genus. Mycotaxon 2003, 86, 375–382. [Google Scholar]
- Ho, W.H.; Yanna; Hyde, K.D.; Goh, T.-K. Endosporoideus gen. nov., a mitosporic fungus on Phoenix hanceana. Mycologia 2005, 97, 238–245. [Google Scholar] [CrossRef]
- Hyde, K.D.; Yanna; Pinnoi, A.; Jones, E.B.G. Goidanichiella fusiforma sp. nov. from palm fronds in Brunei and Thailand. Fungal Divers. 2002, 11, 119–122. [Google Scholar]
- Yanna; Ho, W.H.; Goh, T.-K.; Hyde, K.D. Craspedodidymum nigroseptatum sp. nov., a new hyphomycete on palms from Brunei Darussalam. Mycol. Res. 2000, 104, 1146–1151. [Google Scholar] [CrossRef]
- Ho, W.H.; Yanna; Hyde, K.D. Two new species of Spadicoides from Brunei and Hong Kong. Mycologia 2002, 94, 302–306. [Google Scholar] [CrossRef]
- Haines, J.H. Studies in the Hyaloscyphaceae VI: The genus Lachnum (ascomycetes) of the Guayana Highlands. Nova Hedwig. 1992, 54, 97–112. [Google Scholar]
- Cantrell, S.A.; Haines, J.H. New red species of Lachnum from the tropics. Mycol. Res. 1997, 101, 1081–1084. [Google Scholar] [CrossRef]
- McKenzie, E.H.C.; Pinnoi, A.; Wong, M.K.M.; Hyde, K.D.; Jones, E.B.G. Two new hyaline Chalara species, and a key to species described since 1975. Fungal Divers. 2002, 11, 129–139. [Google Scholar]
- Pinruan, U.; Jones, E.B.G.; Hyde, K.D. Aquatic fungi from peat swamp palms: Jahnula appendiculata sp. nov. Sydowia 2002, 54, 242–247. [Google Scholar]
- Pinruan, U.; Lumyong, S.; McKenzie, E.H.C.; Jones, E.B.G.; Hyde, K.D. Three new species of Craspedodidymum from palm in Thailand. Mycoscience 2004, 45, 177–180. [Google Scholar] [CrossRef]
- Pinruan, U.; McKenzie, E.H.C.; Jones, E.B.G.; Hyde, K.D. Two new species of Stachybotrys, and a key to the genus. Fungal Divers. 2004, 17, 145–157. [Google Scholar]
- Pinnoi, A.; Jones, E.B.G.; McKenzie, E.H.C.; Hyde, K.D. Aquatic fungi from peat swamp palms: Unisetosphaeria penguinoides gen. et sp. nov., and three new Dactylaria species. Mycoscience 2003, 44, 377–382. [Google Scholar] [CrossRef]
- Pinnoi, A.; McKenzie, E.C.; Jones, E.B.G.; Hyde, K.D. Palm fungi from Thailand: Custingophora undulatistipes sp. nov. and Vanakripa minutiellipsoidea sp. nov. Nova Hedwig. 2003, 77, 213–219. [Google Scholar] [CrossRef]
- Pinnoi, A.; Pinruan, U.; Hyde, K.D.; McKenzie, E.H.C.; Lumyong, S. Submersisphaeria palmae sp. nov. with a key to species, and notes on Helicoubisia. Sydowia 2004, 56, 72–78. [Google Scholar]
- Pinnoi, A.; Lumyong, S.; Hyde, K.D.; Jones, E.B.G. Biodiversity of fungi on the palm Eleiodoxa conferta in Sirindhorn peat swamp forest, Narathiwat, Thailand. Fungal Divers. 2006, 22, 205–218. [Google Scholar]
- Pinruan, U.; Hyde, K.D.; Lumyong, S.; McKenzie, E.H.C.; Jones, E.B.G. Occurrence of fungi on tissues of the peat swamp palm Licuala longicalycata. Fungal Divers. 2007, 25, 157–173. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. III. Mycol. Pap. 1961, 82, 1–55. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. V. Mycol. Pap. 1963, 93, 1–33. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. IV. Mycol. Pap. 1963, 87, 1–42. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. VI. Mycol. Pap. 1965, 103, 1–46. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. VII. Curvularia, Brachysporium, etc. Mycol. Pap. 1966, 106, 1–57. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes. IX. Spiropes and Pleurophragmium. Mycol. Pap. 1968, 114, 1–44. [Google Scholar]
- Ellis, M.B. Dematiaceous hyphomycetes. X. Mycol. Pap. 1971, 125, 1–30. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Kew, UK, 1971. [Google Scholar]
- Ellis, M.B. More Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: Kew, UK, 1976. [Google Scholar]
- Subramanian, C.V. A reassessment of Sporidesmium (Hyphomycetes) and some related taxa. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1992, 58, 179–190. [Google Scholar]
- Mena-Portales, J.; Hernández-Gutiérrez, A.; Mercado-Sierra, A. Acarocybiopsis, a new genus of synnematous hyphomycetes from Cuba. Mycol. Res. 1999, 103, 1032–1034. [Google Scholar] [CrossRef]
- Zhang, S.-N.; Hyde, K.D.; Jones, E.B.G.; Cheewangkoon, R.; Liu, J.-K. Acuminatispora palmarum gen. et sp. nov. from mangrove habitats. Mycol. Prog. 2018, 17, 1173–1188. [Google Scholar] [CrossRef]
- Subramanian, C.V. Agrabeeja kavakapriya gen. et sp. nov. and additions to Hemicorynespora. Kavaka 1995, 20, 1–9. [Google Scholar]
- Konta, S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; McKenzie, E.H.C.; Stadler, M.; Boonmee, S.; Phookamsak, R.; Jayawardena, R.S.; Senwanna, C.; Hyde, K.D.; et al. A new genus Allodiatrype, five new species and a new host record of diatrypaceous fungi from palms (Arecaceae). Mycosphere 2020, 11, 239–268. [Google Scholar] [CrossRef]
- do Carmo, L.T.; Monteiro, J.S.; Gusmao, L.F.P.; Sotao, H.M.P.; Gutiérrez, A.H.; Castañeda-Ruíz, R.F. Anabahusakala, a new genus from the Brazilian Amazon rainforest. Mycotaxon 2014, 127, 11–15. [Google Scholar] [CrossRef]
- Qiao, M.; Li, D.-W.; Yu, Z.-F.; Zhang, K.; Castañeda-Ruíz, R.F. Spadicoides matsushimae sp. nov., and Anisospadicoides gen. nov. for two atypical Spadicoides species. Mycotaxon 2019, 134, 161–167. [Google Scholar] [CrossRef]
- Subramanian, C.V. Basauxia and Ashtaangam of Hyphomycetes from Southeast Asia. Kor. J. Mycol. 1992, 20, 281–284. [Google Scholar]
- Phookamsak, R.; Norphanphoun, C.; Tanaka, K.; Dai, D.-Q.; Luo, Z.-L.; Liu, J.-K.; Su, H.-Y.; Bhat, D.J.; Bahkali, A.H.; Mortimer, P.E.; et al. Towards a natural classification of Astrosphaeriella-like species; introducing Astrosphaeriellaceae and Pseudoastrosphaeriellaceae fam. nov. and Astrosphaeriellopsis, gen. nov. Fungal Divers. 2015, 74, 143–197. [Google Scholar] [CrossRef]
- Wu, W.-P.; Diao, Y.-Z. Anamorphic chaetosphaeriaceous fungi from China. Fungal Divers. 2022, 116, 1–546. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Dayarathne, M.C.; Suetrong, S.; Guo, S.Y.; Alias, S.A.; Bahkali, A.H.; Nagahama, T.; Elgorban, A.M.; Abdel-Aziz, F.A.; Hodhod, M.S.; et al. New saprobic marine fungi and a new combination. Bot. Mar. 2017, 60, 469–488. [Google Scholar] [CrossRef]
- Pinruan, U.; Rungjindamai, N.; Sakayaroj, J.; Lumyong, S.; Hyde, K.D.; Jones, E.B.G. Baipadisphaeria gen. nov., a freshwater ascomycete (Hypocreales, Sordariomycetes) from decaying palm leaves in Thailand. Mycosphere 2010, 1, 53–63. [Google Scholar]
- Nagaraju, D.; Kunwar, I.K.; Sureshkumar, G.; Manoharachary, C. A new synnematous hyphomycetous fungus Bhadradriella gen. nov. from India. J. Mycol. Plant Pathol. 2011, 41, 238–240. [Google Scholar]
- Castañeda-Ruíz, R.F.; Iturriaga, T.; Decock, C. Bulbocatenospora, a new hyphomycete genus from Venezuela. Mycol. Res. 2000, 104, 107–109. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Miller, A.N.; Piepenbring, M. South Florida microfungi: Castanedospora, a new genus to accommodate Sporidesmium pachyanthicola (Capnodiales, Ascomycota). Cryptogam. Mycol. 2018, 39, 109–127. [Google Scholar] [CrossRef]
- Gutiérrez, A.H. New or rare fungi from eastern Amazonia. 1. Circinoconiopsis amazonica gen. and sp. nov. Mycotaxon 2013, 123, 107–111. [Google Scholar] [CrossRef]
- Sivanesan, A. Corynesporasca caryotae gen. et sp. nov. with a Corynespora anamorph, and the family Corynesporascaceae. Mycol. Res. 1996, 100, 783–788. [Google Scholar] [CrossRef]
- Boonmee, S.; Wanasinghe, D.N.; Calabon, M.S.; Huanraluek, N.; Chandrasiri, S.K.U.; Jones, G.E.B.; Rossi, W.; Leonardi, M.; Singh, S.K.; Rana, S.; et al. Fungal Diversity notes 1387–1511: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2021, 111, 1–335. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano–Lira, J.F.; Valenzuela–Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, F.A. Two new cheirosporous asexual taxa (Dictyosporiaceae, Pleosporales, Dothideomycetes) from freshwater habitats in Egypt. Mycosphere 2016, 7, 448–457. [Google Scholar] [CrossRef]
- Hongsanan, S.; Bahkali, A.H.; Chomnunti, P.; Liu, J.-K.; Yang, J.-B.; Hyde, K.D. Discopycnothyrium palmae gen. & sp. nov. (Asterinaceae). Mycotaxon 2016, 131, 859–869. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, A.K.; Kamal. New hyphopodiate hyphomycetes from North-Eastern Uttar Pradesh, India. Mycol. Res. 1995, 99, 395–396. [Google Scholar] [CrossRef]
- Liu, J.-K.; Phookamsak, R.; Jones, E.B.G.; Zhang, Y.; Ko–Ko, T.W.; Hu, H.L.; Boonmee, S.; Doilom, M.; Chukeatirote, E.; Bahkali, A.H.; et al. Astrosphaeriella is polyphyletic, with species in Fissuroma gen. nov., and Neoastrosphaeriella gen. nov. Fungal Divers. 2011, 51, 135–154. [Google Scholar] [CrossRef]
- Pinruan, U.; Sakayaroj, J.; Jones, E.B.G.; Hyde, K.D. Flammispora gen. nov., a new freshwater ascomycete from decaying palm leaves. Stud. Mycol. 2004, 50, 381–386. [Google Scholar]
- Hou, L.; Giraldo, A.; Groenewald, J.Z.; Raemae, T.; Summerbell, R.C.; Huang, G.; Cai, L.; Crous, P.W. Redisposition of acremonium-like fungi in Hypocreales. Stud. Mycol. 2023, 105, 23–203. [Google Scholar] [CrossRef]
- Konta, S.; Hyde, K.D.; Eungwanichayapant, P.D.; Karunarathna, S.C.; Samarakoon, M.C.; Xu, J.C.; Dauner, L.A.P.; Aluthwattha, S.T.; Lumyong, S.; Tibpromma, S. Multigene phylogeny reveals Haploanthostomella elaeidis gen. et sp. nov. and familial replacement of Endocalyx (Xylariales, Sordariomycetes, Ascomycota). Life 2021, 11, 486. [Google Scholar] [CrossRef] [PubMed]
- Konta, S.; Hyde, K.V.D.; Karunarathna, S.C.; Mapook, A.; Senwanna, C.; Dauner, L.A.P.; Nanayakkara, C.M.; Xu, J.C.; Tibpromma, S.; Lumyong, S. Multi-gene phylogeny and morphology reveal Haplohelminthosporium gen. nov. and Helminthosporiella gen. nov. associated with palms in Thailand and a checklist for Helminthosporium reported worldwide. Life 2021, 11, 454. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.F.; Heredia, G.; Arias, R.M. Digitella rigidophora and Redbia inflata, two new microfungi from Mexico. Mycotaxon 2013, 125, 227–233. [Google Scholar] [CrossRef]
- Minter, D.W. Nomenclatural novelties: David William Minter. Index Fungorum 2015, 226, 1. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Le Roux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, C.V. Hemisynnema malayasianum gen. et sp. nov. Nova Hedwig. 1994, 58, 223–228. [Google Scholar]
- Delgado-Rodríguez, G. South Florida microfungi: Kalamarospora multiflagellata gen. et sp. nov. (hyphomycetes), with additional new records from USA. Mycotaxon 2010, 114, 231–246. [Google Scholar] [CrossRef]
- Rodrigues, K.F.; Samuels, G.J. Letendraeopsis palrnarum, a new genus and species of loculoascomycetes. Mycologia 1994, 86, 254–258. [Google Scholar] [CrossRef]
- Crous, W.P.; Begoude, B.A.D.; Boers, J.; Braun, U.; Declercq, B.; Dijksterhuis, J.; Elliott, T.F.; Garay-Rodriguez, G.A.; Jurjević, Ž.; Kruse, J.; et al. New and interesting fungi. 5. Fungal. Syst. Evol. 2022, 10, 19–90. [Google Scholar] [CrossRef]
- Konta, S.; Hyde, K.D.; Phookamsak, R.; Xu, J.C.; Maharachchikumbura, S.S.N.; Daranagama, D.A.; McKenzie, E.H.C.; Boonmee, S.; Tibpromma, S.; Eungwanichayapant, P.D.; et al. Polyphyletic genera in Xylariaceae (Xylariales): Neoxylaria gen. nov. and Stilbohypoxylon. Mycosphere 2020, 11, 2629–2651. [Google Scholar] [CrossRef]
- Subramanian, C.V. Nusia gen. nov. for two interesting hyphomycetes. Cryptogam. Mycol. 1993, 14, 109–116. [Google Scholar]
- Pereira, D.S.; Phillips, A.J.L. A new leaf spot disease of Chamaerops humilis caused by Palmeiromyces chamaeropicola gen. et sp. nov. Phytopathol. Mediterr. 2020, 59, 353–363. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal Diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Crous, P.W.; Boers, J.; Holdom, D.; Osieck, E.R.; Steinrucken, T.V.; Tan, Y.P.; Vitelli, J.S.; Shivas, R.G.; Barrett, M.; Boxshall, A.G.; et al. Fungal Planet description sheets: 1383–1435. Persoonia 2022, 48, 261–371. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J.; et al. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs–Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet description sheets: 1182–1283. Persoonia 2021, 46, 313–528. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Ruíz, R.F.; Abarca, G.H.; Arias, R.M.; Saikawa, M.; Minter, D.W.; Stadler, M. Anamorphic fungi from submerged plant material: Phaeomonilia pleiomorpha, P. corticola and Cacumisporium pleuroconidiophorum. Mycotaxon 2007, 100, 327–336. [Google Scholar]
- Pinruan, U.; Sakayaroj, J.; Jones, E.B.G.; Hyde, K.D. Aquatic fungi from peat swamp palms: Phruensis brunneispora gen. et sp nov and its hyphomycete anamorph. Mycologia 2004, 96, 1163–1170. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Gene, J.; Guarro, J.; Baseia, I.G.; Garcia, D.; Gusmao, L.F.P.; Souza-Motta, C.M.; et al. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef]
- Mapook, A.; Macabeo, A.P.G.; Thongbai, B.; Hyde, K.D.; Stadler, M. Polyketide-derived secondary metabolites from a Dothideomycetes fungus, Pseudopalawania siamensis gen. et sp. nov., (Muyocopronales) with antimicrobial and cytotoxic activities. Biomolecules 2020, 10, 569. [Google Scholar] [CrossRef]
- Prabhugaonkar, A.; Bhat, D.J. Rattania setulifera, an undescribed endophytic hyphomycete on rattans from Western Ghats, India. Mycotaxon 2009, 108, 217–222. [Google Scholar] [CrossRef]
- Hernández-Gutiérrez, A.; Mena-Portales, J. A new helicosporous hyphomycete collected on Roystonea regia in Cuba. Mycol. Res. 1996, 100, 1483–1484. [Google Scholar] [CrossRef]
- Dubey, R.; Moonnambeth, N.A. Sawantomyces—A new hyphomycete genus from Western Ghats, India. J. New Biol. Rep. 2013, 2, 234–237. [Google Scholar]
- Pinruan, U.; Sakayaroj, J.; Hyde, K.D.; Jones, E.B.G. Thailandiomyces bisetulosus gen. et sp nov (Diaporthales, Sordariomycetidae, Sordariomycetes) and its anamorph Craspedodidymum, is described based on nuclear SSU and LSU rDNA sequences. Fungal Divers. 2008, 29, 89–98. [Google Scholar]
- Subramanian, C.V. Tretendophragmia palmivora gen. et sp. nov. an interesting hyphomycete from Singapore. Kavaka 1991, 19, 58–66. [Google Scholar]
- Subramanian, C.V. Tretocephala decidua gen. et sp. nov., an interesting new hyphomycete. Cryptogam. Mycol. 1992, 13, 65–68. [Google Scholar]
- Konta, S.; Tibpromma, S.; Karunarathna, S.C.; Samarakoon, M.C.; Steven, L.S.; Mapook, A.; Boonmee, S.; Senwanna, C.; Balasuriya, A.; Eungwanichayapant, P.D.; et al. Morphology and multigene phylogeny reveal ten novel taxa in Ascomycota from terrestrial palm substrates (Arecaceae) in Thailand. Mycosphere 2023, 14, 107–152. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.; Iturriaga, T. Venustusporium, a new genus of hyphomycetes from Venezuela. Mycotaxon 1999, 72, 455–459. [Google Scholar]
- Delgado-Rodríguez, G. South Florida microfungi: Veramycella bispora, a new palmicolous anamorphic genus and species, with some new records for the continental USA. Mycotaxon 2009, 107, 357–373. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.-K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.-Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Voglmayr, H.; Jaklitsch, W.M. Corynespora, Exosporium and Helminthosporium revisited—New species and generic reclassification. Stud. Mycol. 2017, 87, 43–76. [Google Scholar] [CrossRef]
- Wu, W.-P.; Zhuang, W.-Y. Sporidesmium, Endophragmiella and related genera from China. Fung. Divers. Res. Ser. 2005, 15, 1–531. [Google Scholar]
- Wang, Y.; Hyde, K.D.; McKenzie, E.H.C.; Jiang, Y.-L.; Li, D.-W.; Zhao, D.-G. Overview of Stachybotrys (Memnoniella) and current species status. Fungal Divers. 2015, 71, 17–83. [Google Scholar] [CrossRef]
- Goos, R.D. On the anamorph genus Cirrenalia. Proc. Indian Acad. Sci. Plant Sci. 1985, 94, 245–252. [Google Scholar] [CrossRef]
- Goos, R.D. A review of the anamorph genus Helicomyces. Mycologia 1985, 77, 606–618. [Google Scholar] [CrossRef]
- Goos, R.D. The anamorph genus Zalerion. Mycotaxon 1985, 23, 445–449. [Google Scholar]
- Goos, R.D. A review of the anamorph genus Helicoma. Mycologia 1986, 78, 744–761. [Google Scholar] [CrossRef]
- Goos, R.D. Fungi with a twist: The helicosporous hyphomycetes. Mycologia 1987, 79, 1–22. [Google Scholar] [CrossRef]
- Goos, R.D. On the anamorph genera Helicosporium and Drepanospora. Mycologia 1989, 81, 356–374. [Google Scholar] [CrossRef]
- Goos, R.D. Review of the anamorph genus Xenosporium. Mycologia 1990, 82, 742–752. [Google Scholar] [CrossRef]
- Goos, R.D. Fungi of Barro Colorado Island, adjacent Panama, and the Cali region of Colombia. Mycotaxon 1997, 64, 375–383. [Google Scholar]
- Goos, R.D.; Abdullah, S.K.; Fisher, P.J.; Webster, J. The anamorph genus Helicodendron. Trans. Br. Mycol. Soc. 1985, 84, 423–435. [Google Scholar] [CrossRef]
- Subramanian, C.V. Validation of names of some taxa (hyphomycetes). Kavaka 1995, 20, 57–58. [Google Scholar]
- Subramanian, C.V. Hyphomycetes from South East Asia—Novelties from Singapore and Malaysia. Kavaka 1994, 22, 52–76. [Google Scholar]
- Mercado-Sierra, A.; Gonzalez-Fraginals, G.; Mena-Portales, J.; Rodríguez-Morejón, K. Las Palmas y su relación como sustratos de hongos microscópicos (Hifomicetos) en Cuba. Bol. Soc. Micol. Madrid 1997, 22, 34–44. [Google Scholar]
- Castañeda-Ruíz, R.F.; Kendrick, B. Conidial fungi from Cuba: I. Univ. Waterloo Biol. Ser. 1990, 32, 1–53. [Google Scholar]
- Castañeda-Ruíz, R.F.; Kendrick, B. Conidial fungi from Cuba: II. Univ. Waterloo Biol. Ser. 1990, 33, 1–62. [Google Scholar]
- Castañeda-Ruíz, R.F.; Kendrick, B. Ninety-nine conidial fungi from Cuba and three from Canada. Univ. Waterloo Biol. Ser. 1991, 35, 1–132. [Google Scholar]
- Hernández-Gutiérrez, A.; Mena-Portales, J. Sporidesmium coccothrinacis Hernández & Mena, sp. nov. Bol. Soc. Micol. Madrid 1994, 19, 313–314. [Google Scholar]
- Hernández-Gutiérrez, A.; Mena-Portales, J. Nuevos registros de hifomicetos sobre Roystonea regia en Cuba. Bol. Soc. Micol. Madrid 1995, 20, 15–23. [Google Scholar]
- Hernández-Gutiérrez, A.; Mena-Portales, J. Hifomicetos asociados a Coccothrinax (Palmae) en diferentes localidades de la Provincia de Camagüey (Cuba). Bol. Soc. Micol. Madrid 1995, 20, 25–33. [Google Scholar]
- Hernández-Gutiérrez, A.; Mena-Portales, J. Dictyochaeta minutissima sp. nov. on Coccothrinax miraguama from Cuba. Mycol. Res. 1996, 100, 687–688. [Google Scholar] [CrossRef]
- Mercado-Sierra, A.; Mena-Portales, J. Hifomicetes dematiáceos de tres provincias orientales de Cuba. Rev. Iberoam. Micol. 1995, 12, 101–107. [Google Scholar]
- Castañeda-Ruíz, R.F.; Guarro, J.; Mayayo, E.; Decock, C. Notes on conidial fungi. XVI. A new species of Dendryphiosphaera and some new records from Cuba. Mycotaxon 1998, 67, 9–19. [Google Scholar]
- Mercado-Sierra, A.; Holubová-Jechová, V.; Mena-Portales, J. Hifomicetes Demaciáceos de Cuba. Enteroblásticos. Monografie XXIII; Museo Regionale di Scienze Naturali: Turin, Italy, 1997. [Google Scholar]
- Mercado-Sierra, A.; Mena-Portales, J.; Figueras, M.J. Revision of the genus Phragmospathula. Mycologia 1997, 89, 304–308. [Google Scholar] [CrossRef]
- Mercado-Sierra, A.; Figueiras, M.J.; Gené, J. New or rare hyphomycetes from Cuba. VIII. Species of Lylea, Phaeoisaria, Arxiella, Graphium, Periconia and Ramichloridium. Mycotaxon 1997, 63, 369–375. [Google Scholar]
- Mercado-Sierra, A.; Gené, J.; Figueras, M.J.; Rodríguez-Morejón, K.; Guarro, J. New or rare hyphomycetes from Cuba. IX. Some species from Pinar del Rio Province. Mycotaxon 1998, 67, 417–426. [Google Scholar]
- Mena-Portales, J.; Delgado-Rodríguez, G.; Mercado-Sierra, A.; Gené, J.; Guarro, J.; Iacona, V. New or interesting hyphomycetes from the Biosphere Reserve of Sierra del Rosario, Cuba. Mycologia 2001, 93, 751–757. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Mena-Portales, J. Virgariella ellipsospora sp. nov. (Hyphomycetes, Anamorphic fungi) from Cuba. Cryptogam. Mycol. 2003, 24, 153–157. [Google Scholar]
- Mena-Portales, J.; Herrera-Figueroa, S.; Mercado-Sierra, A.; Brito, H.I.; Blanco-Hernández, N.; Medina, J.L.O.; González, S.M.; Herrera, G.R.; Rodríguez-Hernández, M.; Vilaró, M.C.; et al. Estrategia Para la Conservación de la Diversidad Fúngica en Cuba. Estado de Conocimiento, Estrategia y Plan de Acción; Mena-Portales, J., Herrera-Figueroa, S., Mercado-Sierra, A., Minter, D.W., Eds.; Instituto de Ecología y Sistemática: La Habana, Cuba, 2000; Available online: https://repositorio.geotech.cu/xmlui/handle/1234/2728 (accessed on 15 September 2023).
- Delgado-Rodríguez, G.; Mena-Portales, J.; Calduch, M.; Decock, C. Hyphomycetes (Hongos Mitospóricos) del Área Protegida Mil Cumbres, Cuba Occidental. Cryptogam. Mycol. 2002, 23, 277–293. [Google Scholar]
- Delgado-Rodríguez, G.; Mena-Portales, J.; Mercado-Sierra, A. Nuevos registros de hifomicetos en Alturas de Trinidad (Cuba). Bol. Soc. Micol. Madrid 2003, 27, 49–54. [Google Scholar]
- Mercado-Sierra, A.; Delgado-Rodríguez, G.; Mena-Portales, J.; Guarro, J. Some Hyphomycetes (mitosporic fungi) from “Ciénega de Zapata” Biosphere Reserve (Cuba). Bol. Soc. Micol. Madrid 2002, 26, 183–187. [Google Scholar]
- Mena-Portales, J.; Delgado-Rodríguez, G.; Hernández-Gutiérrez, A.; González-Fraginals, G.; Mercado-Sierra, A. Hifomicetes de Sierra de Cubitas, Cuba. Acta Bot. Cub. 2017, 216, 17–30. [Google Scholar]
- Mena-Portales, J.; Cantillo-Pérez, T.; Jimenez-Gómez, I. Hifomicetes de la Reserva de la Biosfera “Ciénaga de Zapata”. Acta Bot. Cub. 2018, 217, 96–108. [Google Scholar]
- Mena-Portales, J.; González-Fraginals, G.; Mercado-Sierra, A.; Cantillo-Pérez, T. Hifomicetes del Parque Nacional “Caguanes”, Reserva de la Biosfera “Buena Vista”, Cuba. Acta Bot. Cub. 2020, 219, 1–9. [Google Scholar]
- Mena-Portales, J.; Cantillo-Pérez, T.; Jiménez-Gómez, I. Hongos microscópicos interesantes hallados sobre palmas en Cuba. Acta Bot. Cub. 2021, 220, 1–5. [Google Scholar]
- Mercado-Sierra, A.; Gene, J.; Guarro, J. Some Costa Rican Hyphomycetes. I. Nova Hedwigia 1997, 64, 455–465. [Google Scholar] [CrossRef]
- Mercado-Sierra, A.; Gene, J.; Guarro, J. Some Costa Rican hyphomycetes. II. Mycotaxon 1997, 64, 7–15. [Google Scholar]
- Granados-Montero, M.; Minter, D.W.; Castañeda-Ruíz, R.F. A checklist of asexual fungi from Costa Rica. Mycotaxon 2018, 133, 365. [Google Scholar] [CrossRef]
- Stevenson, J.A. The Fungi of Puerto Rico and the American Virgin Islands; Contribution of Reed Herbarium No. XXIII; Reed Herbarium: Baltimore, MD, USA, 1975. [Google Scholar]
- Huhndorf, S.M.; Fernández, F. Neotropical ascomycetes 7: Caudatispora biapiculata sp. nov. from Puerto Rico. Sydowia 1998, 50, 200–204. [Google Scholar]
- Guatimosim, E.; Pinto, H.J.; Barreto, R.W. Passalora acrocomiae sp. nov. and Exosporium acrocomiae from the palm Acrocomia aculeata in Puerto Rico. Mycotaxon 2012, 122, 61–67. [Google Scholar] [CrossRef]
- Piepenbring, M. Checklist of fungi in Panama. Preliminary version. Puente Biológico 2006, 1, 1–190. [Google Scholar]
- Delgado-Rodríguez, G. Nicaraguan fungi: A checklist of hyphomycetes. Mycotaxon 2011, 115, 534. [Google Scholar]
- Mercado-Sierra, A.; Heredia-Abarca, G. Hyphomycetes asociados a restos vegetales en el estado de Veracruz, México. Revista Mex. Micol. 1994, 10, 33–48. [Google Scholar] [CrossRef]
- Mercado-Sierra, A.; Heredia-Abarca, G.; Mena-Portales, J. New species of dematiaceous hyphomycetes from Veracruz, Mexico. Mycotaxon 1995, 55, 491–499. [Google Scholar]
- Mercado-Sierra, A.; Heredia-Abarca, G.; Mena-Portales, J. Tropical hyphomycetes of Mexico I. New species of Hemicorynespora, Piricauda and Rhinocladium. Mycotaxon 1997, 63, 155–168. [Google Scholar]
- Heredia-Abarca, G.; Mena-Portales, J.; Mercado-Sierra, A. Hyphomycetes saprobios tropicales. Nuevos registros de dematiáceos para México. Revista Mex. Micol. 1997, 13, 41–51. [Google Scholar] [CrossRef]
- Heredia-Abarca, G.; Mena-Portales, J.; Mercado-Sierra, A.; Estebanez, M.R. Tropical hyphomycetes of Mexico. II. Some species from the tropical biology station “Los Tuxtlas”, Veracruz, Mexico. Mycotaxon 1997, 64, 203–223. [Google Scholar]
- Heredia-Abarca, G.H.; Mercado-Sierra, A. Tropical Hyphomycetes of Mexico. III. Some species from the Calakmul Biosphere Reserve, Campeche. Mycotaxon 1998, 68, 137–143. [Google Scholar]
- Heredia-Abarca, G.; Arias-Mota, R.M.; Estebanez, M.R. Contribución al conocimiento de los hongos Hyphomycetes de México. Acta Bot. Mex. 2000, 51, 39–51. [Google Scholar] [CrossRef]
- Mercado-Sierra, A.; Guarro, J.; Heredia-Abarca, G.H. The hyphomycete genus Piricauda, with the description of a new species. Mycol. Res. 2005, 109, 723–728. [Google Scholar] [CrossRef]
- San Martín, F.E.; Lavin, P.A. Cuatro especies y una variedad del género Astrosphaeriella (Dothideales, Melanommataceae) de México. Acta Bot. Mex. 1999, 46, 19–27. [Google Scholar] [CrossRef]
- Mena-Portales, J.; Delgado-Rodríguez, G.; Heredia-Abarca, G. Nuevas combinaciones para especies de Sporidesmium S.L. (hongos mitospóricos). Bol. Soc. Micol. Madrid 2000, 25, 265–270. [Google Scholar]
- Mena-Portales, J.; Heredia-Abarca, G.; Mercado-Sierra, A.; Becerra-Hernández; Arias-Mota, R.M.; Gómez-Cornelio, S.A. Especies de Stachybotrys Corda (hongos anamorfos) de regiones tropicales y subtropicales de México. Bol. Soc. Micol. Madrid 2009, 33, 7–23. [Google Scholar]
- Heredia-Abarca, G.; Estebanez, M.R.; Arias-Mota, R.M.; Mena-Portales, J.; Mercado-Sierra, A. Adiciones al conocimiento de la diversidad de los hongos conidiales del bosque mesófilo de montaña del Estado de Veracruz. Acta Bot. Mex. 2004, 66, 1–22. [Google Scholar] [CrossRef]
- Heredia-Abarca, G.; Arias-Mota, R.M.; Mena-Portales, J.; Mercado-Sierra, A. Adiciones al conocimiento de la diversidad de los hongos conidiales del bosque mesófilo de montaña del Estado de Veracruz. II. Acta Bot. Mex. 2006, 77, 15–30. [Google Scholar] [CrossRef]
- Heredia-Abarca, G.; Castañeda-Ruíz, R.F.; Becerra-Hernández, C.I.; Arias-Mota, R.M. Contribución al conocimiento de los hongos anamorfos saprobios del Estado de Tabasco. I. Revista Mex. Micol. 2006, 23, 53–62. [Google Scholar] [CrossRef]
- Arias-Mota, R.M.; Heredia-Abarca, G.; Mena-Portales, J. Adiciones al conocimiento de la diversidad de los hongos anamorfos del bosque mesófilo de montaña del estado de Veracruz III. Acta Bot. Mex. 2010, 90, 19–42. [Google Scholar] [CrossRef]
- Arias-Mota, R.M.; Heredia-Abarca, G.; Castañeda-Ruíz, R.F. Adiciones al conocimiento de la diversidad de los hongos conidiales saprobios del bosque mesófilo de montaña del estado de Veracruz IV. Acta Bot. Mex. 2015, 113, 87–101. [Google Scholar] [CrossRef]
- Arias-Mota, R.M.; Heredia-Abarca, G.; Castañeda-Ruíz, R.F. Checklist of saprobic asexual microfungi from the tropical montane cloud forest of Veracruz, México. Mycotaxon 2018, 132, 986. [Google Scholar] [CrossRef]
- Becerra-Hernández, C.I.; Heredia-Abarca, G.; Arias-Mota, R.M. Contribución al conocimiento de los hongos anamorfos saprobios del Estado de Tabasco. II. Revista Mex. Micol. 2007, 24, 39–53. [Google Scholar] [CrossRef]
- Becerra-Hernández, C.I.; Heredia-Abarca, G.; Arias-Mota, R.M.; Castañeda-Ruíz, R.F.; Mena-Portales, J. Especies raras de hongos anamorfos saprobios en el Estado de Tabasco. Acta Bot. Mex. 2011, 96, 15–31. [Google Scholar] [CrossRef]
- Becerra-Hernández, C.I.; Heredia-Abarca, G.; Arias-Mota, R.M.; Mena-Portales, J.; Castañeda-Ruíz, R.F. Los hongos anamorfos saprobios del Estado de Tabasco. III. Rev. Mex. Micol. 2008, 28, 25–39. [Google Scholar] [CrossRef]
- Mercado-Sierra, A.; Basilico, J.C.; Iacona, V.; Luz-Zapata, M. Some interesting mitosporic fungi (hyphomycetes) from Argentina. Bol. Soc. Micol. Madrid 2000, 25, 243–250. [Google Scholar]
- Gómez-Zapata, P.A.; Salazar-Yepes, M. Camarotella colombiana sp. nov. (Phyllachoraceae) sobre Ceroxylon quindiuense (Arecaceae) en Colombia. Revista Mex. Biodivers. 2017, 88, 275–279. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.F.; Iturriaga, T.; Heredia-Abarca, G.; Minter, D.W.; Gené, J.; Stadler, M.; Saikawa, M.; Silvera-Simón, C. Notes on Heteroconium and a new species from Venezuela. Mycotaxon 2008, 105, 175–184. [Google Scholar]
- Castañeda-Ruíz, R.F.; Iturriaga, T.; Minter, D.W.; Heredia-Abarca, G.; Stadler, M.; Saikawa, M.; Fernández, R. Two new anamorphic fungi and some microfungi recorded from ‘El Avila’, Venezuela. Mycotaxon 2009, 107, 225–237. [Google Scholar] [CrossRef]
- Castañeda-Ruíz, R.F.; Iturriaga, T.; Minter, D.W.; Saikawa, M.; Vidal, G.; Velázquez-Noa, S. Microfungi from Venezuela: A new species of Brachydesmiella, a new combination, and new records. Mycotaxon 2003, 85, 211–229. [Google Scholar]
- Castañeda-Ruíz, R.F.; Iturriaga, T.; Saikawa, M.; Cano, J.; Guarro, J. The genus Menisporopsis in Venezuela with the addition of M. anisospora anam. sp nov from a palm tree. Cryptogam. Mycol. 2001, 22, 259–263. [Google Scholar] [CrossRef]
- Monteiro, J.S.; Gusmao, L.F.P.; Castañeda-Ruíz, R.F. Pleurothecium bicoloratum & Sporidesmiopsis pluriseptata spp. nov from Brazil. Mycotaxon 2016, 131, 145–152. [Google Scholar] [CrossRef]
- Capdet, M.; Pereira, S.; Romero, A.I. Coccostromopsis palmicola on Butia yatay from Argentina. Mycotaxon 2010, 114, 91–97. [Google Scholar] [CrossRef]
- Capdet, M.; Romero, A.I. Fungi from palms in Argentina. 1. Mycotaxon 2010, 112, 339–355. [Google Scholar] [CrossRef]
- Capdet, M.; Romero, A.I. Ascomicetes sobre palmeras nativas de la Argentina. II. Anamorfos. Bol. Soc. Argent. Bot. 2012, 47, 303–310. [Google Scholar]
- Subramanian, C.V. Fungi imperfecti from Madras—III. Beltraniella gen. nov. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1952, 36, 223–228. [Google Scholar] [CrossRef]
- Subramanian, C.V. Fungi imperfecti from Madras—II. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1952, 36, 160–169. [Google Scholar] [CrossRef]
- Subramanian, C.V. Fungi imperfecti from Madras—I. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1952, 36, 43–53. [Google Scholar] [CrossRef]
- Subramanian, C.V. Fungi imperfecti from Madras—V. Curvularia. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1953, 38, 27–39. [Google Scholar] [CrossRef]
- Subramanian, C.V. Fungi imperfecti from Madras—IV. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1953, 37, 96–105. [Google Scholar] [CrossRef]
- Subramanian, C.V. Fungi imperfecti from Madras—VI. J. Indian Bot. Soc. 1954, 33, 36–42. [Google Scholar]
- Subramanian, C.V. Fungi imperfecti from Madras—VII. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1955, 42, 283–292. [Google Scholar] [CrossRef]
- Subramanian, C.V. Hyphomycetes. An account of Indian species, except Cercosporae; Indian Council of Agricultural Research: New Delhi, India, 1971. [Google Scholar]
- Subramanian, C.V. Three new hyphomycetes. J. Indian Bot. Soc. 1954, 33, 28–35. [Google Scholar]
- Subramanian, C.V. Some species of Periconia from India. J. Indian Bot. Soc. 1955, 34, 339–361. [Google Scholar]
- Subramanian, C.V. Two new species of Petrakia. Beih. Zur Sydowia 1957, 1, 14–15. [Google Scholar]
- Subramanian, C.V.; Nair, N.G. Panchanania and Phragmospathula, two new genera of the Hyphomycetes. Antonie Leeuwenhoek 1966, 32, 381–386. [Google Scholar] [CrossRef]
- Subramanian, C.V.; Natarajan, K. Two new hyphomycetes from India. Mycologia 1975, 67, 1211–1217. [Google Scholar] [CrossRef]
- Subramanian, C.V.; Ramakrishnan, K. List of Indian Fungi 1952–1956. J. Madras Univ. Sec. B Sci. 1956, 26, 327–421. [Google Scholar]
- Subramanian, C.V.; Bhat, D.J. Developmental morphology of Ascomycetes XI. Nectria kera. Cryptogam. Mycol. 1984, 5, 135–145. [Google Scholar]
- Subramanian, C.V.; Bhat, D.J. Hyphomycetes from south India I. Some new taxa. Kavaka 1987, 15, 41–74. [Google Scholar]
- Subramanian, C.V. Hyphomycetes—II. J. Indian Bot. Soc. 1956, 35, 446–494. [Google Scholar]
- Subramanian, C.V. Hyphomycetes—I. J. Indian Bot. Soc. 1956, 35, 53–91. [Google Scholar]
- Subramanian, C.V. Hyphomycetes—IV. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 1957, 46, 324–335. [Google Scholar] [CrossRef]
- Subramanian, C.V. Hyphomycetes—III. Two new genera, Dwayaloma and Sadasivania. J. Indian Bot. Soc. 1957, 36, 61–67. [Google Scholar]
- Subramanian, C.V. Hyphomycetes—VI. Two new genera, Edmundmasonia and Iyengarina. J. Indian Bot. Soc. 1958, 37, 401–407. [Google Scholar]
- Subramanian, C.V. Hyphomycetes—V. J. Indian Bot. Soc. 1958, 37, 47–64. [Google Scholar]
- Rao, D.; Rao, P.R. ‘Vrikshopama’, a new genus of Stilbaceae. Mycopathol. Mycol. Appl. 1964, 23, 287–290. [Google Scholar] [CrossRef]
- Rao, P.R.; Rao, D. Some Helicosporae from Hyderabad—II. Mycopathol. Mycol. Appl. 1964, 24, 27–34. [Google Scholar] [CrossRef]
- Rao, P.R.; Rao, D. Some species of Camposporium Harkn from India. Antonie Leeuwenhoek 1964, 30, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.R.; Rao, D. Some allied Dematiaceae-Dictyosporae from India. Mycopathol. Mycol. Appl. 1964, 23, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.R.; Rao, D. The genus Periconia from India. Mycopathol. Mycol. Appl. 1964, 22, 285–310. [Google Scholar] [CrossRef]
- Rao, V.; Rao, D. A new Haplobasidion from Hyderabad. Curr. Sci. 1970, 39, 18–19. [Google Scholar]
- Rao, P.R.; Rao, S. A new Hansfordia Hughes from India. Curr. Sci. 1980, 49, 447. [Google Scholar]
- Rao, D.; Rao, V. Studies on Barnettella. Indian Phytopathol. 1973, 26, 233–236. [Google Scholar]
- Chaudhury, R.; Rao, P.N. Ascomycetes from Hyderabad, India I. Mycopathol. Mycol. Appl. 1964, 22, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.N.; Chaudhury, R. A new species of Cochliobolus from Hyderabad—India. Mycopathol. Mycol. Appl. 1964, 23, 36–38. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. Some additions to fungi of India. I. Mycopathol. Mycol. Appl. 1966, 28, 45–48. [Google Scholar] [CrossRef]
- Rao, R. Two new species of Patellaria from India. Mycopathol. Mycol. Appl. 1967, 31, 29–32. [Google Scholar] [CrossRef]
- Rao, R. Some new and noteworthy fungi from India—VII. Sydowia 1971, 24, 322–325. [Google Scholar]
- Varghese, K.I.M.; Rao, V.G. Phaeoisaria Hohnel, a new generic record to Indian Hyphomycetes. Curr. Sci. 1977, 46, 156. [Google Scholar]
- Varghese, K.I.M.; Rao, V.G. Two undescribed species of dematiaceous hyphomycetes. Norw. J. Bot. 1980, 27, 55–57. [Google Scholar]
- Pande, A.; Rao, V.G. The genus Rosellinia (Sphaeriales) from peninsular India. Czech Mycol. 1995, 48, 177–182. [Google Scholar] [CrossRef]
- Bhat, D.J.; Kendrick, B. Twenty-five new conidial fungi from the Western Ghats and the Andaman Islands (India). Mycotaxon 1993, 49, 19–90. [Google Scholar]
- Bhat, D.J. Two undescribed species of conidial fungi from forests of western Ghats in southern India. J. For. Res. 1994, 17, 129–133. [Google Scholar]
- D’Souza, M.; Bhat, D.J. Two new hyphomycetes from India. In Microbes and Plants; Aruna, S., Ed.; Campus Books International: New Delhi, India, 2001; pp. 1–6. [Google Scholar]
- D’Souza, M.; Singh, S.K.; Bhat, D.J. New conidial fungi from Andaman Islands, India. Mycotaxon 2002, 82, 133–143. [Google Scholar]
- D’Souza, M.; Bhat, D.J. Diversity and abundance of endophytic fungi in four plant species of Western Ghat forest of Goa, southern India. Kavaka 2007, 35, 11–20. [Google Scholar]
- D’Souza, M.A.; Bhat, D.J. Occurrence of microfungi as litter colonizers and endophytes in varied plant species from the Western Ghats forests, Goa, India. Mycosphere 2013, 4, 567–582. [Google Scholar] [CrossRef]
- Manoharachary, C.; Sridhar, K.; Singh, R.; Adholeya, A.; Suryanarayanan, T.S.; Rawat, S.; Johri, B.N. Fungal biodiversity: Distribution, conservation and prospecting of fungi from India. Curr. Sci. 2005, 89, 58–71. [Google Scholar]
- Hosagoudar, V.B.; Mathew, S.P. A preliminary report on the mycoflora of the Andaman and Nicobar Islands, India. J. Econ. Taxon. Bot. 2000, 24, 631–640. [Google Scholar]
- Bhat, D.J. The forests of Western Ghats, an abode of novel and interesting microfungi. Kavaka 2008, 36, 1–11. [Google Scholar]
- Bhat, D.J.; Pratibha, J.; Gawas, P.; Sarita, K.Y.; Swapnaja, D. Diversity of microfungi in the forests of Western Ghats in Goa and surrounding regions. In Plant and Fungal Biodiversity and Bioprospecting; Krishnan, S., Bhat, D.J., Eds.; Broadway Book Centre: Goa, India, 2009; pp. 117–133. [Google Scholar]
- Bhat, D.J. Fascinating Microfungi (Hyphomycetes) of Western Ghats, India; Broadway Book Centre: Goa, India, 2010. [Google Scholar]
- Prabhugaonkar, A.; Bhat, D.J. New record of Megacapitula villosa and Paradictyoarthrinium diffractum from India. Mycosphere 2011, 2, 463–467. [Google Scholar]
- Hosagoudar, V.B.; Mathew, S.P.; Babu, D. Foliicolous fungi of Andaman Islands, India. J. Threat. Taxa 2014, 6, 5447–5463. [Google Scholar] [CrossRef]
- Pratibha, J.; Raghukumar, S.; Bhat, D.J. New species of Dendryphiopsis and Stauriella from Goa, India. Mycotaxon 2010, 113, 297–303. [Google Scholar] [CrossRef]
- Pratibha, S.J.; Raghukumar, S.; Bhat, D.J. Diversity of litter degrading microfungi from the forests of Western Ghats, India. In Biodiversity and Taxonomy; Kumar, A.B., Nayar, M.P., Varma, R.V., Peethambaran, C.K., Eds.; Narendra Publishing House: New Delhi, India, 2012; pp. 195–210. [Google Scholar]
- Dubey, R.; Moonnambeth, N.A. Acarocybellina arengae—A new generic and species record from India. Indian Phytopathol. 2013, 66, 326–327. [Google Scholar]
- Dubey, R.; Moonnambeth, N.A. Tharoopama livistonae sp. nov.—A new synematous hyphomycetes from India. Indian J. For. 2013, 36, 383–386. [Google Scholar]
- Dubey, R.; Moonnambeth, N.A. Some new host records for dematiaceous fungi from Western Ghats, India. Nelumbo 2013, 55, 219–221. [Google Scholar] [CrossRef]
- Dubey, R.; Moonnambeth, N.A. Solicorynespora matheranensis sp. nov.—A new species of Solicornespora from Indian subcontinent. NeBIO Int. J. Environ. Biodiversity 2014, 5, 15–18. [Google Scholar]
- Dubey, R.; Moonnambeth, N.A. Phragmospathula brachyspathula Mercado—A record of anamorphic fungi from Western Ghats, India. NeBIO Int. J. Environ. Biodiversity 2014, 5, 25–27. [Google Scholar]
- Dubey, R. Two new species of Zygosporium Mont. from Indian subcontinent. Indian J. Forest. 2014, 37, 165–168. [Google Scholar] [CrossRef]
- Dubey, R.; Neelima, A.M. Some new records of conidial fungi for India. J. New Biol. Rep. 2014, 3, 200–203. [Google Scholar]
- Niranjan, M.; Sarma, V.V. A check-list of fungi from Andaman and Nicobar Islands, India. Phytotaxa 2018, 347, 101–126. [Google Scholar] [CrossRef]
- Ram, T.; Sinha, G.P. A world key to Cryptothecia and Myriostigma (Arthoniaceae), with new species and new records from the Andaman and Nicobar Islands, India. Phytotaxa 2016, 266, 103–114. [Google Scholar] [CrossRef]
- Niranjan, M.; Sarma, V.V. Twelve new species of Ascomycetous fungi from Andaman Islands. Kavaka 2018, 50, 84–97. [Google Scholar]
- Niranjan, M.; Sarma, V.V. New Ascomycetous fungi in the family Aigialaceae from Andaman Islands, India. Curr. Res. Environ. Appl. Mycol. 2018, 8, 351–359. [Google Scholar] [CrossRef]
- Niranjan, M.; Sarma, V.V. New species and new records of Astrosphaeriellaceae from Andaman Islands, India. Kavaka 2020, 54, 38–42. [Google Scholar] [CrossRef]
- Niranjan, M.; Sarma, V.V. Diatrype: New records from Andaman Islands and a checklist from India. MycoAsia J. Mod. Mycol. 2020, 2020/02, 1–17. [Google Scholar] [CrossRef]
- Niranjan, M.; Sarma, V.V. Four novel species of Sordariomycetes from Andaman Islands, India. Kavaka 2021, 56, 105–111. [Google Scholar]
- Subramanian, C.V.; Tyagi, P.D. List of Indian Fungi 1956–1960. J. Madras Univ. Sec. B Sci. 1964, 34, 1–134. [Google Scholar]
- Jamaluddin; Goswami, M.G.; Ojha, B.M. Fungi of India 1989–2001; Scientific Publishers: Jodhpur, India, 2004. [Google Scholar]
- Pande, A. Ascomycetes of Peninsular India; Scientific Publishers: Jodhpur, India, 2008. [Google Scholar]
- Manoharachary, C.; Atri, N.S.; Devi, T.P.; Kamil, D.; Singh, S.K.; Singh, P.A. Bilgrami’s Fungi of India List and References (1988–2020); Today & Tomorrow’s Printers and Publishers: New Delhi, India, 2022. [Google Scholar]
- Pande, A.; Waingahkar, D.; Rao, V.G. Fungal records on palms from India. J. Econ. Taxon. Bot. 2001, 25, 663–686. [Google Scholar]
- Moses, T. Palms of Brazil. Principes 1962, 6, 26–37. [Google Scholar]
- Hennen, J.F.; Ono, Y. Cerradoa palmaea: The first rust fungus on Palmae. Mycologia 1978, 70, 569–576. [Google Scholar] [CrossRef]
- Farr, M.L. A new species of Cryptophiale from Amazonas. Mycotaxon 1980, 11, 177–181. [Google Scholar]
- Farr, Μ.L. Amazonian Foliicolous Fungi III. A preliminary list of Ascomycotina, mostly Dothideales, sensu lato. Acta Amaz. 1985, 15, 29–34. [Google Scholar] [CrossRef]
- Farr, M.L. Amazonian foliicolous fungi. II. Deuteromycotina. Mycologia 1986, 78, 269–286. [Google Scholar] [CrossRef]
- Rodrigues, K.F. The foliar fungal endophytes of the Amazonian palm Euterpe oleracea. Mycologia 1994, 86, 376–385. [Google Scholar] [CrossRef]
- Rodrigues, K.F.; Samuels, G.J. Preliminary study of endophytic fungi in a tropical palm. Mycol. Res. 1990, 94, 827–830. [Google Scholar] [CrossRef]
- Rodrigues, K.F.; Samuels, G.J. Idriella species endophytic in palms. Mycotaxon 1992, 43, 271–276. [Google Scholar]
- Rodrigues, K.F.; Leuchtmann, A.; Petrini, O. Endophytic species of Xylaria: Cultural and isozymic studies. Sydowia 1993, 45, 116–138. [Google Scholar]
- Rodrigues, K.F. Fungal endophytes of palms. In Endophytic Fungi in Grasses and Woody Plants. Systematics, Ecology and Evolution; Redlin, S.C., Carris, L.M., Eds.; APS Press: Saint Paul, MN, USA, 1996; pp. 121–132. [Google Scholar]
- Rodrigues, K.F.; Petrini, O. Biodiversity of endophytic fungi in tropical regions. In Biodiversity of Tropical Microfungi; Hyde, K.D., Ed.; Hong Kong University Press: Hong Kong, China, 1997; pp. 57–69. [Google Scholar]
- Silva, M.S.; Minter, D.W. Fungi from Brazil recorded by Batista and co-workers. Mycol. Pap. 1995, 169, 1–585. [Google Scholar]
- Mendes, M.A.S.; da Silva, V.L.; Dianese, J.C.; Ferreira, M.A.S.V.; dos Santos, C.E.N.; Neto, E.G.; Urben, A.F.; Carlos, C. Fungos em Plantas no Brasil; Embrapa-SPI/Embrapa-Cenargen: Brasília, Brazil, 1998. [Google Scholar]
- Mendes, M.A.S.; Urben, A.F.; Dianese, J.C.; Lobo, V.L.S.; Ferreira, M.A.S.V.; Simon, M.F.; Sanchez, M.; dos Santos, L.T.P.; dos Santos, C.E.N. Fungos em Plantas no Brasil. Edição Ampliada e Revisada, 2nd ed.; Mendes, M.A.S., Urben, A.F., Dianese, J.C., Eds.; Escola Nacional de Gestão Agropecuária: Brasília, Brazil, 2019. Available online: https://enagro.agricultura.gov.br/gestao-do-conhecimento/fungos-em-plantas-no-brasil (accessed on 15 September 2023).
- da Cruz, A.C.R.; Hernández-Gutiérrez, A.; Gusmão, L.F.P. O gênero Exserticlava (Fungo Anamorfo—Hyphomycetes) no Brasil. Braz. J. Bot. 2008, 31, 357–361. [Google Scholar] [CrossRef]
- Monteiro, J.S.; Hernández-Gutiérrez, A.; Sotão, H.M.P. Fungos anamorfos (hyphomycetes) da Floresta Nacional de Caxiuanã, Pará, Brasil: Novos registros para o Neotrópico. Acta Bot. Bras. 2010, 24, 868–870. [Google Scholar] [CrossRef]
- Monteiro, J.S.; do Carmo, L.T.; Sotão, H.M.P. A new species of Bhatia (asexual ascomycetes) and new records from Brazil. Phytotaxa 2017, 331, 263–272. [Google Scholar] [CrossRef]
- Monteiro, J.S.; Sotão, H.M.O.; Cáceres, M.E.S.; Lücking, R.; Hernández-Gutiérrez, A. Checklist dos fungos da Floresta Nacional de Caxiuanã, Pará, Brasil. I. Fungos conidiais e liquenizados. Bol. Mus. Para. Emílio Goeldi Sér. Ciências Naturais 2018, 13, 221–245. [Google Scholar] [CrossRef]
- Monteiro, J.S.; Sarmento, P.S.M.; Sotão, H.M.P. Saprobic conidial fungi associated with palm leaf litter in eastern Amazon, Brazil. An. Acad. Bras. Ciênc. 2019, 91, e20180545. [Google Scholar] [CrossRef]
- de Castro, C.C.; Hernández-Gutiérrez, A.; Sotão, H.M.P. Novos registros de fungos anamorfos (hifomicetos) para o Neotrópico e América do Sul. Braz. J. Bot. 2011, 34, 515–521. [Google Scholar] [CrossRef]
- de Castro, C.C.; Hernández-Gutiérrez, A.; Sotão, H.M.P. Fungos conidiais em Euterpe oleracea Mart. (açaizeiro) na Ilha do Combu, Pará-Brasil. Acta Bot. Bras. 2012, 26, 761–771. [Google Scholar] [CrossRef]
- Dianese, J.C.; Medeiros, R.B.; Santos, L.T.P. Cerradoa palmaea Hennen & Ono found in two new hosts, Syagrus commosa (Mart.) Mart. and S. flexuosa Becc. Fitopatol. Bras. 1992, 17, 198. [Google Scholar]
- Dianese, J.C.; Medeiros, R.B.; Santos, L.T.P. Biodiversity of microfungi found on native plants of the Brazilian Cerrado. In Biodiversity of Tropical Microfungi; Hyde, K.D., Ed.; Hong Kong University Press: Hong Kong, China, 1997; pp. 367–417. [Google Scholar]
- Dianese, J.C.; Inácio, C.A.; Carvalho-Junior, A.A.; dos Santos, M.D.M.; Cantillo-Pérez, T.; Pinho, D.B. Exploring the overlooked diversity of plant-associated cerrado microfungi. Anu. Patol. Plantas 2022, 28, 69–101. [Google Scholar] [CrossRef]
- Medeiros, R.B.; Dianese, J.C. Passalora eitenii sp. nov. on Syagrus comosa in Brazil and a key to Passalora species. Mycotaxon 1994, 51, 509–513. [Google Scholar]
- de Souza, C.A.P.; Victória, N.S.; Bezerra, J.L.; Luz, E.D.M.N.; Inácio, C.A.; Dianese, J.C. Camarotella brasiliensis sp. nov. (Phyllachoraceae) on Syagrus schizophylla (Arecaceae) from Brazil. Mycotaxon 2008, 103, 313–317. [Google Scholar]
- Souza, E.S.C.; Pereira-Carvalho, R.C.; Sanchez, M.; Dianese, J.C. Echidnodella species (Asterinaceae, Ascomycota) on Mauritia flexuosa (Arecaceae) from the Brazilian Cerrado. Phytopathology 2013, 103, 137. [Google Scholar]
- Vitória, N.S.; Bezerra, J.L.; Gramacho, K.P.; Luz, E.D.M.N. Camaroetella torrendiella comb. nov. e C. acrocomiae: Agentes etiológicos das lixas do coqueiro. Trop. Plant Pathol. 2008, 33, 295–301. [Google Scholar] [CrossRef]
- Vitoria, N.S.; Cavalcanti, M.A.Q.; Hyde, K.D.; Bezerra, J.L. Arecomyces new to Brazil, including A. attaleae sp. nov. Cryptogam. Mycol. 2011, 32, 103–108. [Google Scholar] [CrossRef]
- Vitoria, N.S.; Cavalcanti, M.A.Q.; Luz, E.D.M.N.; Bezerra, J.L. Endocalyx melanoxanthus var. melanoxanthus (Ascomycota): New to Brazil and three new hosts. Mycotaxon 2011, 117, 109–113. [Google Scholar] [CrossRef]
- Vitoria, N.S.; Cavalcanti, M.A.Q.; Hyde, K.D.; Bezerra, J.L. Brunneiapiospora brasiliensis sp. nov. (Clypeosphaeriaceae) on palms from Brazil. Nova Hedwig. 2012, 94, 245–250. [Google Scholar] [CrossRef]
- Vitória, N.S.; Cavalcanti, M.A.Q.; Bezerra, J.L. Lasiodiplodia theobromae: A new host and a revision of plant hosts reported in Brazil. Agrotrópica 2012, 24, 63–66. [Google Scholar] [CrossRef]
- Vitoria, N.S.; Cavalcanti, M.A.Q.; dos Santos, C.D.; Pereira, J.; Bezerra, J.L. Neolinocarpon attaleae sp. nov. on Attalea funifera (Arecaceae) from Brazil. Mycotaxon 2013, 123, 141–145. [Google Scholar] [CrossRef]
- Vitória, N.S.; Cavalcanti, M.A.Q.; dos Santos, M.V.O.; Silvério, M.L.; Bezerra, J.L. Ascomycota em palmeiras: Novos registros e novos hospedeiros para o Nordeste Brasileiro. Agrotrópica 2014, 26, 35–42. [Google Scholar] [CrossRef]
- Vitória, N.S.; dos Santos, M.A.L.; Fortes, N.G.S. Comunidade fúngica de Syagrus coronata (Mart.) Becc: Ascomycota anamórficos e teleomórficos. In Ecologia e Biodiversidade do Semiárido Nordestino; de Andrade, M.J.G., Nogueira, E.M.S., dos Santos, C.A.B., Eds.; Sociedade Brasileira de Ecologia Humana: Bahia, Brazil, 2016; pp. 35–45. [Google Scholar]
- Vitória, N.S.; Cavalcanti, M.A.D.; Bezerra, J.L. Species of Astrosphaeriella and Fissuroma from palms: New records for South America and Brazil. Nova Hedwig. 2016, 102, 129–140. [Google Scholar] [CrossRef]
- Vitória, N.S.; dos Santos, M.A.L.; Bezerra, J.L. Contribuições para o conhecimento de fungos (Ascomycota) em Mauritia flexuosa L.f. e Acrocomia intumescens Drude, Brasil. Rev. Bras. Geogr. Fís. 2019, 12, 1252–1258. [Google Scholar] [CrossRef]
- Vitória, N.S.; dos Santos, M.A.L.; Bezerra, J.L. Euterpe oleracea Mart. e Elaeis guineensis Jacq. (Arecaceae): Novos hospedeiros para o registro de ocorrência de microfungos (Ascomycota) no Brasil. In Pindorama; dos Santos, F.A.R., Carneiro, C.H., Eds.; Editora da Universidade Federal do Piauí: Piauí, Brazil, 2019; pp. 119–133. [Google Scholar]
- Vitória, N.S.; Fortes, N.G.S.; dos Santos, M.A.L.; Barbosa, R.L. Mycota (Ascomycota) of Syagrus coronata (Mart.) Becc., Raso da Catarina Ecological Station, Brazil: New records. Acta Bras. 2020, 4, 110–120. [Google Scholar] [CrossRef]
- Cruz, A.C.R.; Izabel, T.S.S.; Leão-Ferreira, S.M.; Gusmão, L.F.P. Conidial fungi from the semi-arid Caatinga biome of Brazil. New species and new records of Helicosporium. Mycotaxon 2009, 110, 53–64. [Google Scholar] [CrossRef]
- Cruz, A.C.R.; Gusmão, L.F.P. Fungos conidiais na Caatinga: Espécies associadas ao folhedo. Acta Bot. Bras. 2009, 23, 999–1012. [Google Scholar] [CrossRef]
- Cruz, A.C.R.; Gusmão, L.F.P. Fungos conidiais na Caatinga: Espécies lignícolas. Acta Bot. Bras. 2009, 23, 1133–1144. [Google Scholar] [CrossRef]
- Adamcík, S.; Cai, L.; Chakraborty, D.; Chen, X.-H.; Cotter, H.V.; Dai, D.-Q.; Dai, Y.-C.; Das, K.; Deng, C.-Y.; Ghobad-Nejhad, M.; et al. Fungal Biodiversity Profiles 1–10. Cryptogam. Mycol. 2015, 36, 121–166. [Google Scholar] [CrossRef]
- de Oliveira, R.J.V.; Bezerra, J.L.; Lima, T.E.F.; da Silva, G.A.; Cavalcantil, M.A.D. Phaeosphaeria nodulispora, a new endophytic coelomycete isolated from tropical palm (Cocos nucifera) in Brazil. Nova Hedwig. 2016, 103, 185–192. [Google Scholar] [CrossRef]
- dos Santos, M.A.L.; Vitória, N.S.; Bezerra, J.L. Fungos colonizando palmeiras em áreas de Caatinga do Sertão da Bahia. Agrotrópica 2016, 28, 37–46. [Google Scholar] [CrossRef]
- dos Santos, E.C.S.; Vitória, N.S. Espécies de Ascomycota em Syagrus coronata (Mart.) Becc., Água Branca, Alagoas, Brasil. Rev. Ouricuri 2017, 7, 80–97. [Google Scholar]
- dos Santos, M.A.L.; Fortes, N.G.S.; Silva, T.E.F.; Vitória, N.S. Ascomycota (lichenized and non-lichenized) on Syagrus coronata in the Caatinga biome: New and interesting records for Brazil and South America. Mycotaxon 2019, 134, 737. [Google Scholar] [CrossRef]
- dos Santos, M.A.L.; Bezerra, J.L.; Vitória, N.S. Phaeoseptum aquaticum (Halotthiaceae): New record for American continent in a new host for science. Rodriguésia 2019, 70, e00282018. [Google Scholar] [CrossRef]
- dos Santos, M.A.L.; Vitória, N.S.; de Oliveira, R.J.V.; Bezerra, J.L. Diatrypella heveae Senwanna, Phookamsak & K.D. Hyde (Diatrypaceae, Xylariales): A new record for the Neotropics. Check List 2020, 16, 1703–1708. [Google Scholar] [CrossRef]
- Rocha, P.Q.; Vitória, N.S. New occurrences of ascomycetes for South America and the Neotropics. Agrotrópica 2020, 32, 31–36. [Google Scholar] [CrossRef]
- Fortes, N.G.S.; dos Santos, M.A.L.; Vitória, N.S. Apiosordaria nigeriensis (Ascomycota): A new record for the Americas. Rodriguésia 2020, 71, e00852018. [Google Scholar] [CrossRef]
- da Silva, M.S.R.; Vitória, N.S. Fungos endofíticos em frutos de Syagrus coronata (Mart.) Becc. Encicl. Biosf. 2023, 20, 188–203. [Google Scholar] [CrossRef]
- Vitória, N.S.; dos Santos, M.A.L.; Souza, V.M.F.; da Silva, T.B.M.; Bezerra, J.L. Sexual morph of Stachybotrys frondicola (Ascomycota): First record in Brazil. Encicl. Biosf. 2022, 19, 35–45. [Google Scholar] [CrossRef]
- Rocha, P.Q.; Barbosa, R.L.; Vitória, N.S. Ascomycetes in Syagrus coronata (Mart.) Becc. in the Raso da Catarina Ecological Station, with new distribution records. Rev. Ouricuri 2023, 13, 222–247. [Google Scholar] [CrossRef]
- McKenzie, E.H.C. A new species of Lylea (hyphomycetes) on Rhopalostylis (Arecaceae) in New Zealand. Mycotaxon 2009, 109, 39–42. [Google Scholar] [CrossRef]
- McKenzie, E.H.C. Two new dictyosporous hyphomycetes on Rhopalostylis sapida (Arecaceae) in New Zealand. Mycotaxon 2010, 111, 155–160. [Google Scholar] [CrossRef]
- Petrini, L.E. Rosellinia and related genera in New Zealand. N. Z. J. Bot. 2003, 41, 71–138. [Google Scholar] [CrossRef]
- Braun, U.; Hill, C.F.; Schubert, K. New species and new records of biotrophic micromycetes from Australia, Fiji, New Zealand and Thailand. Fungal Divers. 2006, 22, 13–35. [Google Scholar]
- Johnston, P.R.; Whitton, S.R.; Buchanan, P.K.; Park, D.; Pennycook, S.R.; Johnson, J.E.; Moncalvo, J.M. The basidiomycete genus Favolaschia in New Zealand. N. Z. J. Bot. 2006, 44, 65–87. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hawksworth, D.L.; Hyde, K.D.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Manamgoda, D.S.; Thambugala, K.M.; Udayanga, D.; Camporesi, E.; Daranagama, A.; et al. Epitypification and neotypification: Guidelines with appropriate and inappropriate examples. Fungal Divers. 2014, 69, 57–91. [Google Scholar] [CrossRef]
- Pinruan, U.; Rungjindamai, N.; Choeyklin, R.; Lumyong, S.; Hyde, K.D.; Jones, E.B.G. Occurrence and diversity of basidiomycetous endophytes from the oil palm, Elaeis guineensis in Thailand. Fungal Divers. 2010, 41, 71–88. [Google Scholar] [CrossRef]
- Pinnoi, A.; Jeewon, R.; Sakayaroj, J.; Hyde, K.D.; Jones, E.B.G. Berkleasmium crunisia sp. nov. and its phylogenetic affinities to the Pleosporales based on 18S and 28S rDNA sequence analyses. Mycologia 2007, 99, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Pinnoi, A.; Phongpaichit, P.; Jeewon, R.; Tang, A.M.C.; Hyde, K.D.; Jones, E.B.G. Phylogenetic relationships of Astrocystis eleiodoxae sp. nov. (Xylariaceae). Mycosphere 2010, 1, 1–9. [Google Scholar]
- Bahl, J.; Jeewon, R.; Hyde, K.D. Phylogeny of Rosellinia capetribulensis sp nov and its allies (Xylariaceae). Mycologia 2005, 97, 1102–1110. [Google Scholar] [CrossRef]
- Hidayat, I.; Jeewon, R.; To-Anun, C.; Hyde, K.D. The genus Oxydothis: New palmicolous taxa and phylogenetic relationships within the Xylariales. Fungal Diver. 2006, 23, 159–179. [Google Scholar]
- Konta, S.; Hongsanan, S.; Tibpromma, S.; Thongbai, B.; Maharachchikumbura, S.S.N.; Bahkali, A.H.; Hyde, K.D.; Boonmee, S. An advance in the endophyte story: Oxydothidaceae fam. nov. with six new species of Oxydothis. Mycosphere 2016, 7, 1425–1446. [Google Scholar] [CrossRef]
- Hu, H.M.; Liu, L.L.; Zhang, X.; Lin, Y.; Shen, X.C.; Long, S.H.; Kang, J.C.; Wijayawardene, N.N.; Li, Q.R.; Long, Q.D. New species and records of Neomassaria, Oxydothis and Roussoella (Pezizomycotina, Ascomycota) associated with palm and bamboo from China. Mycokeys 2022, 93, 165–191. [Google Scholar] [CrossRef]
- Konta, S.; Hongsanan, S.; Liu, J.K.; Eungwanichayapant, P.D.; Jeewon, R.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Boonmee, S. Leptosporella (Leptosporellaceae fam. nov.) and Linocarpon and Neolinocarpon (Linocarpaceae fam. nov.) are accommodated in Chaetosphaeriales. Mycosphere 2017, 8, 1943–1974. [Google Scholar] [CrossRef]
- Zhang, S.N.; Hyde, K.D.; Jones, E.B.G.; Cheewangkoon, R.; Liu, J.K. Additions to Fissuroma and Neoastrosphaeriella (Aigialaceae, Pleosporales) from palms. Mycosphere 2020, 11, 269–284. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Jeewon, R.; Jones, E.B.G.; Boonmee, S.; Kaewchai, S.; Manawasinghe, I.S.; Lumyong, S.; Hyde, K.D. Novel palmicolous taxa within Pleosporales: Multigene phylogeny and taxonomic circumscription. Mycol. Prog. 2018, 17, 571–590. [Google Scholar] [CrossRef]
- Konta, S.; Hyde, K.D.; Eungwanichayapant, P.D.; Doilom, M.; Tennakoon, D.S.; Senwanna, C.; Boonmee, S. Fissuroma (Aigialaceae: Pleosporales) appears to be hyperdiverse on Arecaceae: Evidence from two new species from southern Thailand. Acta Bot. Bras. 2020, 34, 384–393. [Google Scholar] [CrossRef]
- Liu, J.K.; Phookamsak, R.; Dai, D.Q.; Tanaka, K.; Jones, E.B.G.; Xu, J.C.; Chukeatirote, E.; Hyde, K.D. Roussoellaceae, a new pleosporalean family to accommodate the genera Neoroussoella gen. nov., Roussoella and Roussoellopsis. Phytotaxa 2014, 181, 1–33. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; Hongsanan, S.; Phukhamsakda, C.; Li, J.F.; Boonmee, S. Palawaniaceae fam. nov., a new family (Dothideomycetes, Ascomycota) to accommodate Palawania species and their evolutionary time estimates. Mycosphere 2016, 7, 1732–1745. [Google Scholar] [CrossRef]
- Yu, X.D.; Zhang, S.N.; Cheewangkoon, R.; Liu, J.K. Additions to Occultibambusaceae (Pleosporales, Dothideomycetes): Unrevealing Palmicolous Fungi in China. Diversity 2021, 13, 516. [Google Scholar] [CrossRef]
- Hawksworth, D.L. ‘Misidentifications’ in fungal DNA sequence databanks. New Phytol. 2004, 161, 13–15. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Liu, X.Z.; Chamyuang, S.; Stadler, M.; Bahkali, A.H.; Hyde, K.D. Towards a natural classification of Sordariomycetes: The genera Frondisphaeria, Immersisphaeria, Lasiobertia, Pulmosphaeria and Yuea (Sordariomycetes incertae sedis). Phytotaxa 2016, 258, 153–163. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Stadler, M.; Jones, E.B.G.; Promputtha, I.; Suwannarach, N.; Camporesi, E.; Bulgakov, T.S.; Liu, J.K. Taxonomy, phylogeny, molecular dating and ancestral state reconstruction of Xylariomycetidae (Sordariomycetes). Fungal Divers. 2022, 112, 1–88. [Google Scholar] [CrossRef]
- Hyde, K.V.D.; Dong, Y.; Phookamsak, R.T.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal Diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Hidayat, I.; Meeboon, J.; To–Anun, C. Anthostomella and Fasciatispora species (Xylariaceae) from palms including F. ujungkulonensis sp. nov. Mycotaxon 2007, 102, 347–354. [Google Scholar]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal Diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.Z.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Jeewon, R.; Liu, X.Z.; Stadler, M.; Lumyong, S.; Hyde, K.D. Taxonomic rearrangement of Anthostomella (Xylariaceae) based on a multigene phylogeny and morphology. Cryptogam. Mycol. 2016, 37, 509–538. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Miller, A.N.; Hashimoto, A.; Iida, T.; Ohkuma, M.; Okada, G. A phylogenetic assessment of Endocalyx (Cainiaceae, Xylariales) with E. grossus comb. et stat. nov. Mycol. Prog. 2022, 21, 221–242. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.R.; Wijesinghe, S.N.; Raza, M.; Bao, D.F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Pennycook, S.R.; Johnston, P.R.; Ramaley, A.; Akulov, A.; Crous, P.W. Resolving the phylogenetic and taxonomic status of dark-spored teleomorph genera in the Botryosphaeriaceae. Persoonia 2008, 21, 29–55. [Google Scholar] [CrossRef]
- Konta, S.; Phillips, A.J.L.; Bahkali, A.H.; Jones, E.B.G.; Eungwanichayapant, D.P.; Hyde, K.D.; Boonmee, S. Botryosphaeriaceae from palms in Thailand—Barriopsis archontophoenicis sp. nov, from Archontophoenix alexandrae. Mycosphere 2016, 7, 921–932. [Google Scholar] [CrossRef]
- Konta, S.; Hongsanan, S.; Phillips, A.J.L.; Jones, E.B.G.; Boonmee, S.; Hyde, K.D. Botryosphaeriaceae from palms in Thailand II—Two new species of Neodeightonia, N. rattanica and N. rattanicola from Calamus (rattan palm). Mycosphere 2016, 7, 950–961. [Google Scholar] [CrossRef]
- Douanla–Meli, C.; Scharnhorst, A. Palm foliage as pathways of pathogenic Botryosphaeriaceae fungi and host of new Lasiodiplodia species from Mexico. Pathogens 2021, 10, 1297. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.R.; Manawasinghe, I.S.; Liao, C.F.; Hyde, K.D.; Dong, Z.Y. Neodeightonia arengae sp. nov., Botryosphaeriaceous taxa on Arenga tremula (Arecaceae) from Guangdong, China. Phytotaxa 2022, 530, 130–140. [Google Scholar] [CrossRef]
- Wu, N.; Dissanayake, A.J.; Chethana, K.W.T.; Liu, J.-K. Neodeightonia septata sp. nov. and N. subglobosa (Botryosphaeriaceae) from Northern Thailand. Phytotaxa 2022, 575, 129–139. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef]
- Liu, A.R.; Chen, S.C.; Lin, X.M.; Wu, S.Y.; Xu, T.; Cai, F.M.; Jeewon, R. Endophytic Pestalotiopsis species associated with plants of Palmae, Rhizophoraceae, Planchonellae and Podocarpaceae in Hainan, China. Afr. J. Microbiol. Res. 2010, 4, 2661–2669. [Google Scholar]
- Zhang, Y.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Hyde, K.D. A novel species of Pestalotiopsis causing leaf spots of Trachycarpus fortunei. Cryptogam. Mycol. 2012, 33, 311–318. [Google Scholar] [CrossRef]
- Geng, K.; Zhang, B.; Song, Y.; Hyde, K.D.; Kang, J.C.; Wang, Y. A new species of Pestalotiopsis from leaf spots of Licuala grandis from Hainan, China. Phytotaxa 2013, 88, 49–54. [Google Scholar] [CrossRef]
- Xiong, Y.R.; Manawasinghe, I.S.; Maharachchikumbura, S.S.N.; Lu, L.; Dong, Z.Y.; Xiang, M.M.; Xu, B. Pestalotioid species associated with palm species from Sothern China. Curr. Res. Environ. Appl. Mycol. 2022, 12, 285–321. [Google Scholar] [CrossRef]
- Guterres, D.C.; Silva, M.A.; Martins, M.D.; Azevedo, D.M.Q.; Lisboa, D.O.; Pinho, D.B.; Furtado, G.Q. Leaf spot caused by Neopestalotiopsis species on Arecaceae in Brazil. Australas. Plant Pathol. 2023, 52, 47–62. [Google Scholar] [CrossRef]
- Jiang, N.; Liang, Y.M.; Tian, C.M. Morphological and phylogenic evidences reveal a new Seiridium species in China. Phytotaxa 2019, 418, 287–293. [Google Scholar] [CrossRef]
- Pereira, D.S.; Phillips, A.J.L. Two new Morinia species from palms (Arecaceae) in Portugal. Mycol. Prog. 2021, 20, 83–94. [Google Scholar] [CrossRef]
- Li, J.F.; Phookamsak, R.; Jeewon, R.; Tibpromma, S.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Chukeatirote, E.; Lumyong, S.; Hyde, K.D.; McKenzie, E.H.C. Establishment of Zygosporiaceae fam. nov. (Xylariales, Sordariomycetes) based on rDNA sequence data to accommodate Zygosporium. Mycosphere 2017, 8, 1855–1868. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G. South Florida microfungi: New records of saprophytic hyphomycetes on plant debris. Fla. Sci. 2008, 71, 76–89. [Google Scholar]
- Delgado-Rodríguez, G. South Florida microfungi: A new species of Stanjehughesia (hyphomycetes) from Sabal palm. Mycotaxon 2008, 103, 229–234. [Google Scholar]
- Delgado-Rodríguez, G. South Florida microfungi: A new species of Ellisembia (hyphomycetes) with new records from the USA. Mycotaxon 2013, 123, 445–450. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G. South Florida microfungi: Linkosia longirostrata, a new hyphomycete on paurotis palm. Mycotaxon 2014, 129, 41–46. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Miller, A.N. South Florida microfungi: A new species of Taeniolella (anamorphic Sordariomycetes) isolated from cabbage palm. Nova Hedwig. 2017, 105, 1–14. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Koukol, O.; Cáceres, O.; Piepenbring, M. The phylogenetic placement of Ernakulamia cochinensis within Pleosporales (Dothideomycetes, Ascomycota). Cryptogam. Mycol. 2017, 38, 435–451. [Google Scholar] [CrossRef]
- Koukol, O.; Delgado-Rodríguez, G.; Hofmann, T.A.; Piepenbring, M. Panama, a hot spot for Hermatomyces (Hermatomycetaceae, Pleosporales) with five new species, and a critical synopsis of the genus. IMA Fungus 2018, 9, 107–141. [Google Scholar] [CrossRef] [PubMed]
- Nuankaew, S.; Suetrong, S.; Wutikhun, T.; Pinruan, U. Hermatomyces trangensis sp. nov., a new dematiaceous hyphomycete (Hermatomycetaceae, Pleosporales) on sugar palm in Thailand. Phytotaxa 2019, 391, 277–288. [Google Scholar] [CrossRef]
- Delgado-Rodríguez, G.; Koukol, O.; Heredia-Abarca, G.; Piepenbring, M. Texas microfungi: Hermatomyces amphisporus (Pleosporales, Dothideomycetes) revisited. Czech Mycol. 2020, 72, 95–107. [Google Scholar] [CrossRef]
- Prasher, I.B.; Sushma. Hermatomyces indicus sp. nov. (Hyphomycetes) from India. Nova Hedwig. 2014, 99, 551–556. [Google Scholar] [CrossRef]
- Koukol, O.; Delgado-Rodríguez, G. Do not forget Africa—Revision of fungarium collections at Kew revealed a new species of Hermatomyces (Hermatomycetaceae, Pleosporales). Nova Hedwig. 2019, 109, 413–423. [Google Scholar] [CrossRef]
- Chen, Y.P.; Tian, W.H.; Guo, Y.B.; Madrid, H.; Maharachchikumbura, S.S.N. Synhelminthosporium gen. et sp. nov. and two new species of Helminthosporium (Massarinaceae, Pleosporales) from Sichuan Province, China. J. Fungi 2022, 8, 712. [Google Scholar] [CrossRef] [PubMed]
- Kularathnage, N.D.; Wanasinghe, D.N.; Senanayake, I.C.; Yang, Y.H.; Manawasinghe, I.S.; Phillips, A.J.L.; Hyde, K.D.; Dong, W.; Song, J.G. Microfungi associated with ornamental palms: Byssosphaeria phoenicis sp. nov. (Melanommataceae) and Pseudocoleophoma rhapidis sp. nov. (Dictyosporiaceae) from south China. Phytotaxa 2022, 568, 149–169. [Google Scholar] [CrossRef]
- Tian, X.G.; Tibpromma, S.; Karunarathna, S.C.; Dai, D.Q.; Lu, Y.Z.; Mapook, A.; Jayawardena, R.S. A new species and a new host record of Pseudoberkleasmium (Pseudoberkleasmiaceae, Dothideomycetes) from Cocos nucifera and Zea mays in northern Thailand. Phytotaxa 2022, 547, 232–242. [Google Scholar] [CrossRef]
- Kularathnage, N.D.; Senanayake, I.C.; Wanasinghe, D.N.; Doilom, M.; Stephenson, S.L.; Song, J.G.; Dong, W.; Xu, B. Plant-associated novel didymellaceous taxa in the South China Botanical Garden (Guangzhou, China). J. Fungi 2023, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Manawasinghe, I.S.; Wanasinghe, D.N.; Hongsanan, S.; Hyde, K.D.; Biao, X.; Dong, Z. Two new species and a new host record of Pleosporales (Dothideomycetes) from palm (Arecaceae) in Guangdong Province, China. N. Z. J. Bot. 2023, 1–27. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J. Two new species of Chaetopsina (Nectriaceae) from Saül (French Guiana). Ascomycete.org 2019, 11, 127–134. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J. Clonostachys spinulosispora (Hypocreales, Bionectriaceae), a new species on palm from French Guiana. Ascomycete.org 2018, 10, 127–130. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J.; Chaduli, D.; Lesage-Meessen, L.; Favel, A. Clonostachys saulensis (Bionectriaceae, Hypocreales), a new species from French Guiana. Ascomycete.org 2019, 11, 65–68. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J.; Chaduli, D.; Favel, A. Hydropisphaera palmicola (Bionectriaceae), a new species from Saül (French Guiana). Ascomycete.org 2022, 14, 81–84. [Google Scholar] [CrossRef]
- Lechat, C.; Lesage-Meessen, L.; Favel, A. A new species of Ijuhya, I. fournieri, from French Guiana. Ascomycete.org 2015, 7, 101–104. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J. Four new species of Ijuhya (Bionectriaceae) from Belgium, metropolitan France and French Guiana. Ascomycete.org 2017, 9, 11–18. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J.; Chaduli, D.; Favel, A. Lasionectria saulensis (Bionectriaceae, Hypocreales), a new species from French Guiana. Ascomycete.org 2022, 14, 85–88. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J.; Chaduli, D.; Favel, A. Three new holomorphic species of Volutella (Nectriaceae, Hypocreales) from Saül (French Guiana). Ascomycete.org 2022, 14, 89–95. [Google Scholar] [CrossRef]
- Crane, J.L.; Miller, A.N. Studies in genera similar to Torula: Bahusaganda, Bahusandhika, Pseudotorula, and Simmonsiella gen. nov. IMA Fungus 2016, 7, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Aoki, T.; Ariyawansa, H.A.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Fungal taxonomy and sequence-based nomenclature. Nat. Microbiol. 2021, 6, 540–548. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Phillips, A.J.L.; Pereira, D.S.; Dai, D.Q.; Aptroot, A.; Monteiro, J.S.; Druzhinina, I.S.; Cai, F.; Fan, X.L.; Selbmann, L.; et al. Forecasting the number of species of asexually reproducing fungi (Ascomycota and Basidiomycota). Fungal Divers. 2022, 114, 463–490. [Google Scholar] [CrossRef]
- Wulandari, N.F.; To-Anun, C.; McKenzie, E.H.C.; Hyde, K.D. Guignardia bispora and G. ellipsoidea spp. nov. and other Guignardia species from palms (Arecaceae). Mycosphere 2011, 2, 115–128. [Google Scholar]
- Lechat, C.; Fournier, J. Two new species of Lasionectria (Bionectriaceae, Hypocreales) from Guadeloupe and Martinique (French West Indies). Mycotaxon 2012, 121, 275–280. [Google Scholar] [CrossRef]
- Prasher, I.B.; Verma, R.K. Two new species of Dictyosporium from India. Phytotaxa 2015, 204, 193–202. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Gao, J.M.; Zhang, X.G.; Li, Z. Two new species of Endophragmiella from southern China. Nova Hedwig. 2016, 103, 349–357. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Phillips, A.J.L.; Tibpromma, S.; Dai, D.-Q.; Selbmann, L.; Monteiro, J.S.; Aptroot, A.; Flakus, A.; Rajeshkumar, K.C.; Coleine, C.; et al. Looking for the undiscovered asexual taxa: Case studies from lesser studied life modes and habitats. Mycosphere 2021, 12, 1186–1229. [Google Scholar] [CrossRef]
- Suetrong, S.; Schoch, C.L.; Spatafora, J.W.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; Sakayaroj, J.; Phongpaichit, S.; Tanaka, K.; Hirayama, K.; Jones, E.B.G. Molecular systematics of the marine Dothideomycetes. Stud. Mycol. 2009, 64, 155–173. [Google Scholar] [CrossRef]
- Suetrong, S.; Sakayaroj, J.; Phongpaichit, S.; Jones, E.B.G. Morphological and molecular characteristics of a poorly known marine ascomycete, Manglicola guatemalensis (Jahnulales: Pezizomycotina; Dothideomycetes, Incertae sedis): New lineage of marine ascomycetes. Mycologia 2010, 102, 83–92. [Google Scholar] [CrossRef]
- Suetrong, S.; Hyde, K.D.; Zhang, Y.; Bahkali, A.H.; Jones, E.B.G. Trematosphaeriaceae fam. nov. (Dothideomycetes, Ascomycota). Cryptogam. Mycol. 2011, 32, 343–358. [Google Scholar] [CrossRef]
- Suetrong, S.; Klaysuban, A.; Sakayaroj, J.; Preedanon, S.; Ruang-Areerate, P.; Phongpaichit, S.; Pang, K.L.; Jonese, E.B.G. Tirisporellaceae, a new family in the order Diaporthales (Sordariomycetes, Ascomycota). Cryptogam. Mycol. 2015, 36, 319–330. [Google Scholar] [CrossRef]
- Liu, J.K.; Jones, E.B.G.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Lignincola conchicola from palms with a key to the species of Lignincola. Mycotaxon 2011, 117, 343–349. [Google Scholar] [CrossRef]
- Abdel-Aziz, F.A. The genus Lolia from freshwater habitats in Egypt with one new species. Phytotaxa 2016, 267, 279–288. [Google Scholar] [CrossRef]
- Boonmee, S.; D’Souza, M.J.; Luo, Z.L.; Pinruan, U.; Tanaka, K.; Su, H.Y.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Taylor, J.E.; et al. Dictyosporiaceae fam. nov. Fungal Divers. 2016, 80, 457–482. [Google Scholar] [CrossRef]
- Yang, J.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Liu, Z.Y. New species in Dictyosporium, new combinations in Dictyocheirospora and an updated backbone tree for Dictyosporiaceae. Mycokeys 2018, 36, 83–105. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Devadatha, B.; Abdel-Wahab, M.A.; Dayarathne, M.C.; Zhang, S.N.; Hyde, K.D.; Liu, J.K.; Bahkali, A.H.; Sarma, V.V.; Tibell, S.; et al. Phylogeny of new marine Dothideomycetes and Sordariomycetes from mangroves and deep-sea sediments. Bot. Mar. 2020, 63, 155–181. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.-Q.; Dai, Y.-C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal Diversity notes 111–252: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal Diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal Diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Wang, S.; Sun, Y.R.; Suwannarach, N.; Sysouphanthong, P.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Abeywickrama, P.D.; Abreu, V.P.; et al. Fungal Diversity notes 1512–1610: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2022, 117, 1–272. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Romberg, M.; Mel’nik, V.A.; Verkley, G.J.M.; Groenewald, J.Z. Fungal Planet description sheets: 92–106. Persoonia 2011, 27, 130–162. [Google Scholar] [CrossRef]
- Crous, P.W.; Shivas, R.G.; Wingfield, M.J.; Summerell, B.A.; Rossman, A.Y.; Alves, J.L.; Adams, G.C.; Barreto, R.W.; Bell, A.; Coutinho, M.L.; et al. Fungal Planet description sheets: 128–153. Persoonia 2012, 29, 146–201. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Summerell, B.A.; Giraldo, A.; Gené, J.; Guarro, J.; Wanasinghe, D.N.; Hyde, K.D.; Camporesi, E.; et al. Fungal Planet description sheets: 281–319. Persoonia 2014, 33, 212–292. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Hernandez-Restrepo, M.; Sutton, D.A.; Acharya, K.; Barber, P.A.; Boekhout, T.; Dimitrov, R.A.; Dueñas, M.; et al. Fungal Planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Lombard, L.; Roets, F.; Swart, W.J.; Alvarado, P.; Carnegie, A.J.; Moreno, G.; Luangsa-Ard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Osieck, E.R.; Shivas, R.G.; Tan, Y.P.; Bishop-Hurley, S.L.; Esteve-Raventós, F.; Larsson, E.; Luangsa-Ard, J.J.; Pancorbo, F.; Balashov, S.; et al. Fungal Planet description sheets: 1478–1549. Persoonia 2023, 50, 158–310. [Google Scholar] [CrossRef]
- Boonmee, S.; Phookamsak, R.; Hongsanan, S.; Doilom, M.; Mapook, A.; McKenzie, E.H.C.; Bhat, D.J.; Hyde, K.D. Mycosphere notes 51–101. Revision of genera in Perisporiopsidaceae and Pseudoperisporiaceae and other Ascomycota genera incertae sedis. Mycosphere 2017, 8, 1695–1801. [Google Scholar] [CrossRef]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; de Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Calabon, M.S.; Jones, E.B.G.; Zhang, Y.X.; Liao, C.F.; Xiong, Y.R.; Chaiwan, N.; Kularathnage, N.D.; Liu, N.G.; Tang, S.M.; et al. Mycosphere notes 345–386. Mycosphere 2022, 13, 454–557. [Google Scholar] [CrossRef]
- Pem, D.; Jeewon, R.; Bhat, D.J.; Doilom, M.; Boonmee, S.; Hongsanan, S.; Promputtha, I.; Xu, J.C.; Hyde, K.D. Mycosphere notes 275–324: A morpho-taxonomic revision and typification of obscure Dothideomycetes genera (incertae sedis). Mycosphere 2019, 10, 1115–1246. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Wanasinghe, D.N.; Papizadeh, M.; Goonasekara, I.D.; Camporesi, E.; Bhat, D.J.; McKenzie, E.H.C.; Phillips, A.J.L.; Diederich, P.; et al. Taxonomy and phylogeny of dematiaceous coelomycetes. Fungal Divers. 2016, 77, 1–316. [Google Scholar] [CrossRef]
- Crous, W.P.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. 2. Fungal. Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Crous, W.P.; Wingfield, M.J.; Schumacher, R.K.; Akulov, A.; Bulgakov, T.S.; Carnegie, A.J.; Jurjević, Ž.; Decock, C.; Denman, S.; Lombard, L.; et al. New and interesting fungi. 3. Fungal. Syst. Evol. 2020, 6, 157–231. [Google Scholar] [CrossRef] [PubMed]
- Crous, W.P.; Hernández-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Kölling, G.; Marais, E.; Wingfield, M.J.; Yilmaz, N.; Adan, O.C.G.; Akulov, A.; et al. New and interesting fungi. 4. Fungal. Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; de Silva, N.I.; Jeewon, R.; Bhat, D.J.; Phookamsak, R.; Doilom, M.; Boonmee, S.; Jayawardena, R.S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Li, W.J.; McKenzie, E.H.C.; Liu, J.K.; Bhat, D.J.; Dai, D.Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.N.; Luo, Z.L.; Shang, Q.J.; et al. Taxonomy and phylogeny of hyaline–spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Niranjan, M.; Dong, W.; Samarakoon, M.C.; Bao, D.F.; Calabon, M.S.; Chaiwan, N.; Chuankid, B.; Dayarathne, M.C.; de Silva, N.I.; et al. AJOM new records and collections of fungi: 101–150. Asian J. Mycol. 2021, 4, 113–260. [Google Scholar]
- Crous, P.W.; Carris, L.M.; Giraldo, A.; Groenewald, J.Z.; Hawksworth, D.L.; Hernández-Restrepo, M.; Jaklitsch, W.M.; Lebrun, M.H.; Schumacher, R.K.; Stielow, J.B.; et al. The Genera of Fungi—Fixing the application of the type species of generic names—G 2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 2015, 6, 163–198. [Google Scholar] [CrossRef] [PubMed]
- To-Anun, C.; Nguenhom, J.; Meeboon, J.; Hidayat, I. Two fungi associated with necrotic leaflets of areca palms (Areca catechu). Mycol. Prog. 2009, 8, 115–121. [Google Scholar] [CrossRef]
- Kinge, T.R.; Mih, A.M. Ganoderma ryvardense sp. nov. associated with basal stem rot (BSR) disease of oil palm in Cameroon. Mycosphere 2011, 2, 179–188. [Google Scholar]
- Mbenoun, M.; de Beer, Z.W.; Wingfield, M.J.; Wingfield, B.D.; Roux, J. Reconsidering species boundaries in the Ceratocystis paradoxa complex, including a new species from oil palm and cacao in Cameroon. Mycologia 2014, 106, 757–784. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Guarnaccia, V.; Vitale, A.; Cirvilleri, G.; Granata, G.; Epifani, F.; Perrone, G.; Polizzi, G.; Groenewald, J.Z.; Crous, P.W. Ilyonectria palmarum sp. nov. causing dry basal stem rot of Arecaceae. Eur. J. Plant Pathol. 2014, 138, 347–359. [Google Scholar] [CrossRef]
- Yanna; Ho, W.H.; Hyde, K.D. Fungal succession on fronds of Phoenix hanceana in Hong Kong. Fungal Divers. 2002, 10, 185–211. [Google Scholar]
- Southcott, K.A.; Johnson, J.A. Isolation of endophytes from two species of palm, from Bermuda. Can. J. Microbiol. 1997, 43, 789–792. [Google Scholar] [CrossRef]
- Girivasan, K.P.; Suryanarayanan, T.S. Intact leaves as substrate for fungi: Distribution of endophytes and phylloplane fungi in rattan palms. Czech Mycol. 2004, 56, 33–43. [Google Scholar] [CrossRef]
- Song, J.J.; Pongnak, W.; Soytong, K. Isolation and identification of endophytic fungi from 10 species palm trees. Int. J. Agric. Technol. 2016, 12, 349–363. [Google Scholar]
- Jiaojiao, S.; Wattanachai, P.; Kasem, S. Biological activity of endophytic fungi from palm trees against chili anthracnose caused by Colletotrichum capsica. Int. J. Agric. Technol. 2015, 11, 1927–1940. [Google Scholar]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. A method to promote sporulation in palm endophytic fungi. Fungal Divers. 1998, 1, 109–113. [Google Scholar]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Detection and taxonomic placement of endophytic fungi within frond tissues of Livistona chinensis based on rDNA sequences. Mol. Phylogenet. Evol. 2001, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Vidal, S.; Lopez-Llorca, L.V.; Jansson, H.B.; Salinas, J. Endophytic colonization of date palm (Phoenix dactylifera L.) leaves by entomopathogenic fungi. Micron 2006, 37, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Rungjindamai, N.; Pinruan, U.; Choeyklin, R.; Hattori, T.; Jones, E.B.G. Molecular characterization of basidiomycetous endophytes isolated from leaves, rachis and petioles of the oil palm, Elaeis guineensis, in Thailand. Fungal Divers. 2008, 33, 139–161. [Google Scholar]
- Mahmoud, F.M.; Krimi, Z.; Macia-Vicente, J.G.; Errahmani, M.B.; Lopez-Llorca, L.V. Endophytic fungi associated with roots of date palm (Phoenix dactylifera) in coastal dunes. Rev. Iberoam. Micol. 2017, 34, 116–120. [Google Scholar] [CrossRef]
- Saengket, M.; Hyde, K.D.; Kumar, V.; Doilom, M.; Brooks, S. Endophytic fungi from Oncosperma sp. with promising in vitro plant growth promotion and antagonistic activities. Chiang Mai J. Sci. 2021, 48, 837–852. [Google Scholar]
- Azuddin, N.F.; Mohd, M.H.; Rosely, N.F.N.; Mansor, A.; Zakaria, L. Molecular phylogeny of endophytic fungi from rattan (Calamus castaneus Griff.) spines and their antagonistic activities against plant pathogenic fungi. J. Fungi 2021, 7, 301. [Google Scholar] [CrossRef] [PubMed]
- Ben Chobba, I.; Elleuch, A.; Ayadi, I.; Khannous, L.; Namsi, A.; Cerqueira, F.; Drira, N.; Gharsallah, N.; Vallaeys, T. Fungal diversity in adult date palm (Phoenix dactylifera L.) revealed by culture-dependent and culture-independent approaches. J. Zhejiang Univ. Sci. B 2013, 14, 1084–1099. [Google Scholar] [CrossRef] [PubMed]
- Viana Diniz, F.; Doi, S.M.S.R.; Fittipaldy, M.C.P.M.; Lopes, R.F.; Margarido, S.S.O.R.; Pontes, S.M.A.; Ramos, D.P.; Araújo, A.V.; Carvalho, C.M. Isolation and identification of endophytic fungi from the amazonian palm Oenocarpus bataua Mart. S. Am. J. Basic Educ. Tech. Technol. 2021, 8, 139–153. [Google Scholar]
- Chase, A.R.; Broschat, T.K. Diseases and Disorders of Ornamental Palms; American Phytopathological Society Press: Saint Paul, MN, USA, 1991. [Google Scholar]
- Elliott, M.L. Diseases caused by “fungi”. In Compendium of Ornamental Palm Diseases and Disorders; Elliott, M.L., Broschat, T.K., Uchida, J.Y., Simone, G.W., Eds.; American Phytopathological Society Press: Saint Paul, MN, USA, 2004; pp. 8–37. [Google Scholar]
- Downer, A.J.; Uchida, J.Y.; Hodel, D.R.; Elliott, M.L. Lethal palm diseases common in the United States. HortTechnology 2009, 19, 710–716. [Google Scholar] [CrossRef]
- Mohammadi, H. Phaeoacremonium spp. and Botryosphaeriaceae spp. associated with date palm (Phoenix dactylifera L.) decline in Iran. J. Phytopathol. 2014, 162, 575–581. [Google Scholar] [CrossRef]
- Al-Hammadi, M.S.; Al-Shariqi, R.; Maharachchikumbura, S.S.N.; Al-Sadi, A.M. Molecular identification of fungal pathogens associated with date palm root diseases in the United Arab Emirates. J. Plant Pathol. 2019, 101, 141–147. [Google Scholar] [CrossRef]
- Alwahshi, K.J.; Saeed, E.E.; Sham, A.; Alblooshi, A.A.; Alblooshi, M.M.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular identification and disease management of date palm sudden decline syndrome in the United Arab Emirates. Int. J. Mol. Sci. 2019, 20, 923. [Google Scholar] [CrossRef]
- Al-Nadabi, H.; Maharachchikumbura, S.S.N.; Al-Gahaffi, Z.S.; Al-Hasani, A.S.; Velazhahan, R.; Al-Sadi, A.M. Molecular identification of fungal pathogens associated with leaf spot disease of date palms (Phoenix dactylifera). All Life 2020, 13, 587–597. [Google Scholar] [CrossRef]
- Nishad, R.; Ahmed, T.A. Survey and identification of date palm pathogens and indigenous biocontrol agents. Plant Dis. 2020, 104, 2498–2508. [Google Scholar] [CrossRef]
- Asensio, L.; López-Jiménez, J.A.; López-Llorca, L.V. Mycobiota of the date palm phylloplane: Description and interactions. Rev. Iberoam. Micol. 2007, 24, 299–304. [Google Scholar] [CrossRef]
- Kirkman, E.R.; Hilton, S.; Sethuraman, G.; Elias, D.M.O.; Taylor, A.; Clarkson, J.; Soh, A.C.; Bass, D.; Ooi, G.T.; McNamara, N.P.; et al. Diversity and ecological guild analysis of the oil palm fungal microbiome across root, rhizosphere, and soil compartments. Front. Microbiol. 2022, 13, 15. [Google Scholar] [CrossRef]
- Seephueak, P.; Preecha, C.; Seephueak, W. Diversity of macrofungi in oil palm (Elaeis guineensis jacq.) plantation in Southern Thailand. Walailak J. Sci. Technol. 2018, 15, 201–211. [Google Scholar] [CrossRef]
- Nobre, C.P.; da Costa, M.G.; Goto, B.T.; Gehring, C. Arbuscular mycorrhizal fungi associated with the babassu palm (Attalea speciosa) in the eastern periphery of Amazonia, Brazil. Acta Amaz. 2018, 48, 321–329. [Google Scholar] [CrossRef]
- Fisher, J.B.; Jayachandran, K. Beneficial role of arbuscular mycorrhizal fungi on florida native palms. Palms 2008, 52, 113–123. [Google Scholar]
- Polanco, G.; Carrillo, L.; Espadas, C.; Reyes-García, C.; Guadarrama, P.; Orellana, R. Asociación micorrízica arbuscular en Coccothrinax readii Quero. Trop. Subtrop. Agroecosystems 2013, 16, 223–233. [Google Scholar]
- Furrazola, E.; Sánchez-Rendon, J.A.; Guadarrama, P.; Pernús, M.; Torres-Arias, Y. Mycorrhizal status of Coccothrinax crinita (Arecaceae), an endangered endemic species from western Cuba. Rev. Mex. Biodivers. 2020, 91, 10. [Google Scholar] [CrossRef]
- Ambili, K.; Thomas, G.V.; Indu, P.; Gopal, M.; Gupta, A. Distribution of arbuscular mycorrhizae associated with coconut and arecanut based cropping systems. Agric. Res. 2012, 1, 338–345. [Google Scholar] [CrossRef]
- Ramos-Zapata, J.A.; Orellana, R.; Allen, E.B. Mycorrhizal dynamics and dependence of Desmoncus orthacanthos Martius (Arecaceae), a native palm of the Yucatan Peninsula, Mexico. Interciencia 2006, 31, 364–370. [Google Scholar]
- Asano, K.; Kagong, W.V.A.; Mohammad, S.M.B.; Sakazaki, K.; Abu Talip, M.S.; Sahmat, S.S.; Chan, M.K.Y.; Isoi, T.; Kano-Nakata, M.; Ehara, H. Arbuscular mycorrhizal communities in the roots of sago palm in mineral and shallow peat soils. Agriculture 2021, 11, 1161. [Google Scholar] [CrossRef]
- Bouamri, R.; Dalpé, Y.; Serrhini, M.N. Effect of seasonal variation on arbuscular mycorrhizal fungi associated with date palm. Emir. J. Food Agric. 2014, 26, 977–986. [Google Scholar] [CrossRef]
- Zougari-Elwedi, B.; Islami, W.; Msetra, A.; Sanaa, M.; Yolande, D.; Sahraoui, A.L. Monitoring the evolution of the arbuscular mycorrhizal fungi associated with date palm. J. New Sci. 2016, 31, 1822–1831. [Google Scholar]
- Pinruan, U.; Pinnoi, A.; Hyde, K.D.; Jones, E.B.G. Tropical peat swamp fungi with special reference to palms. In Freshwater Fungi and Fungal-Like Organisms; Jones, E.B.G., Hyde, K.D., Pang, K.L., Eds.; de Gruyter: Berlin, Germany, 2014; pp. 371–388. [Google Scholar] [CrossRef]
- Lateef, A.; Muid, S.; Bolhassan, M.H. Microfungi on leaves of Licuala bidentata (Arecaceae) from Sarawak, Malaysia. Makara J. Sci. 2015, 19, 161–166. [Google Scholar] [CrossRef]
- Pilantanapak, A.; Jones, E.B.G.; Eaton, R.A. Marine fungi on Nypa fruticans in Thailand. Bot. Mar. 2005, 48, 365–373. [Google Scholar] [CrossRef]
- Hyde, K.D.; Sarma, V.V. Biodiversity and ecological observations on filamentous fungi of mangrove palm Nypa fruticans Wurumb (Liliopsida, Arecales) along the Tutong River, Brunei. Indian J. Mar. Sci. 2006, 35, 297–307. [Google Scholar]
- Besitulo, A.; Moslem, M.A.; Hyde, K.D. Occurrence and distribution of fungi in a mangrove forest on Siargao Island, Philippines. Bot. Mar. 2010, 53, 535–543. [Google Scholar] [CrossRef]
- Loilong, A.; Sakayaroj, J.; Rungjindamai, N.; Choeyklin, R.; Jones, E.B.G. Biodiversity of fungi on the palm Nypa fruticans. In Marine Fungi and Fungal-Like Organisms; Jones, E.B.G., Pang, K.L., Eds.; de Gruyter: Berlin, Germany, 2012; pp. 273–290. [Google Scholar] [CrossRef]
- Sarma, V.V.; Hyde, K.D. Fungal species consortia on Nypa fruticans at Brunei. Stud. Fungi 2018, 3, 19–26. [Google Scholar] [CrossRef]
- Hyde, K.D.; Taylor, J.E. The palm fungi. In Proceedings of the Asia-Pacific Mycological Conference on Biodiversity and Biotechnology, Hua Hin, Thailand, 6–9 July 1998; pp. 34–38. [Google Scholar]
- Hyde, K.D.; Fröhlich, J. Ascomycetes associated with palms. In Ecology of Fungi; Bhat, D.J., Raghukumar, S., Eds.; Goa University: Goa, India, 2000; pp. 109–114. [Google Scholar]
- Pinnoi, A.; Pinruan, U.; Hyde, K.D.; Lumyong, S.; Jones, E.B.G. Palm fungi. In Thai Fungal Diversity; Jones, E.B.G., Tanticharoen, M., Hyde, K.D., Eds.; National Center for Genetic Engineering and Biotechnology, BIOTEC: Bangkok, Thailand, 2004; pp. 181–187. [Google Scholar]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales . Fungal Divers. 2012, 53, 1–221. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Huang, S.K.; Abdel–Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’Souza, M.J.; Goonasekara, I.D.; et al. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, J.D.; Dayarathne, M.C.; Huang, S.K.; Norphanphoun, C.; Senanayake, I.C.; Perera, R.H.; et al. Families of Sordariomycetes. Fungal Divers. 2016, 79, 317. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.N.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee, M.P.A.; Wingfield, M.J.; Crous, P.W. Evolution of lifestyles in Capnodiales. Stud. Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel–Wahab, M.A.; Abdel–Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Hongsanan, S.A.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Lücking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.G.; et al. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Ekanayaka, A.H.; Hyde, K.D.; Gentekaki, E.; McKenzie, E.H.C.; Zhao, Q.; Bulgakov, T.S.; Camporesi, E. Preliminary classification of Leotiomycetes. Mycosphere 2019, 10, 310–489. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez–Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Divakar, P.K.; Rajeshkumar, K.C.; Weerahewa, D.; Delgado, G.; Wang, Y.; Fu, L. Notes for genera update—Ascomycota: 6616–6821. Mycosphere 2018, 9, 115–140. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; McKenzie, E.H.C.; Wang, Y. Notes for genera update—Ascomycota: 6822–6917. Mycosphere 2018, 9, 1222–1234. [Google Scholar] [CrossRef]
- Læssøe, T.; Spooner, B.M. Rosellinia & Astrocystis (Xylariaceae): New species and generic concepts. Kew Bull. 1993, 49, 1–70. [Google Scholar] [CrossRef]
- Li, Q.R.; Zhang, X.; Lin, Y.; Samarakoon, M.C.; Hyde, K.D.; Shen, X.C.; Liao, W.Q.; Karunarathna, A.; Long, S.H.; Kang, Y.Q.; et al. Morpho-molecular characterisation of Arecophila, with A. australis and A. clypeata sp. nov. and A. miscanthi comb. nov. Mycokeys 2022, 88, 123–149. [Google Scholar] [CrossRef] [PubMed]
- Fernández, F.A.; Huhndorf, S.M. New species of Chaetosphaeria, Melanopsammella and Tainosphaeria gen. nov from the Americas. Fungal Divers. 2005, 18, 15–57. [Google Scholar]
- Elliott, M.L.; Des Jardin, E.A. Serenomyces associated with palms in southeastern USA: Isolation, culture storage and genetic variation. Mycologia 2014, 106, 698–707. [Google Scholar] [CrossRef]
- Mardones, M.; Trampe-Jaschik, T.; Oster, S.; Elliott, M.; Urbina, H.; Schmitt, I.; Piepenbring, M. Phylogeny of the order Phyllachorales (Ascomycota, Sordariomycetes): Among and within order relationships based on five molecular loci. Persoonia 2017, 39, 74–90. [Google Scholar] [CrossRef]
- Boonyuen, N.; Chuaseeharonnachai, C.; Suetrong, S.; Sri-Indrasutdhi, V.; Sivichai, S.; Jones, E.B.G.; Pang, K.L. Savoryellales (Hypocreomycetidae, Sordariomycetes): A novel lineage of aquatic ascomycetes inferred from multiple-gene phylogenies of the genera Ascotaiwania, Ascothailandia, and Savoryella. Mycologia 2011, 103, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Dayarathne, M.C.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; Dong, W.; Devadatha, B.; Yang, J.; Ekanayaka, A.H.; De Silva, W.; Sarma, V.V.; Al-Sadi, A.M.; et al. Phylogenetic revision of Savoryellaceae and evidence for its ranking as a subclass. Front. Microbiol. 2019, 10, 26. [Google Scholar] [CrossRef]
- Zhang, S.N.; Abdel-Wahab, M.A.; Jones, E.B.G.; Hyde, K.D.; Liu, J.K. Additions to the genus Savoryella (Savoryellaceae), with the asexual morphs Savoryella nypae comb. nov. and S. sarushimana sp. nov. Phytotaxa 2019, 408, 195–207. [Google Scholar] [CrossRef]
- Pratibha, J.; Prabhugaonkar, A. Multi-gene phylogeny of Pithomyces with the sexual morph of P. flavus Berk. & Broome. Phytotaxa 2015, 218, 84–90. [Google Scholar] [CrossRef]
- Elliott, M.L.; Morales, A.D.; Des Jardin, E.A. New records of Botryosphaeriaceae genera associated with palms in Florida, USA. Sydowia 2018, 70, 169–178. [Google Scholar] [CrossRef]
- Rathnayaka, A.R.; Chethana, K.W.T.; Phillips, A.J.L.; Jones, E.B.G. Two new species of Botryosphaeriaceae (Botryosphaeriales) and new host/geographical records. Phytotaxa 2022, 564, 8–38. [Google Scholar] [CrossRef]
- Tian, Q.; Liu, J.K.; Hyde, K.D.; Wanasinghe, D.N.; Boonmee, S.; Jayasiri, S.C.; Luo, Z.L.; Taylor, J.E.; Phillips, A.J.L.; Bhat, D.J.; et al. Phylogenetic relationships and morphological reappraisal of Melanommataceae (Pleosporales). Fungal Divers. 2015, 74, 267–324. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, R. Taxonomy of asexual microfungus Periconia on Phoenix in India. Mycol. Iran. 2015, 2, 65–68. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal Diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Reblova, M.; Kolarik, M.; Nekvindova, J.; Reblova, K.; Sklenar, F.; Miller, A.N.; Hernandez-Restrepo, M. Phylogenetic Reassessment, taxonomy, and biogeography of Codinaea and similar fungi. J. Fungi 2021, 7, 1097. [Google Scholar] [CrossRef]
- Elliott, M.L.; Des Jardin, E.A.; O’Donnell, K.; Geiser, D.M.; Harrison, N.A.; Broschat, T.K. Fusarium oxysporum f. sp. palmarum, a novel forma specialis causing a lethal disease of Syagrus romanzoffiana and Washingtonia robusta in Florida. Plant Dis. 2010, 94, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Jayawardena, R.S.; Bhunjun, C.S.; Harishchandra, D.L.; Hyde, K.D. Pseudocercospora dypsidis sp. nov. (Mycosphaerellaceae) on Dypsis lutescens leaves in Thailand. Phytotaxa 2020, 474, 218–234. [Google Scholar] [CrossRef]
- Braun, U.; Crous, P.; Kamal. New species of Pseudocercospora, Pseudocercosporella, Ramularia and Stenella (cercosporoid hyphomycetes). Mycol. Prog. 2003, 2, 197–208. [Google Scholar] [CrossRef]
- Braun, U.; Crous, P.W.; Nakashima, C. Cercosporoid fungi (Mycosphaerellaceae) 2. Species on monocots (Acoraceae to Xyridaceae, excluding Poaceae). IMA Fungus 2014, 5, 203–390. [Google Scholar] [CrossRef]
- Ma, L.G.; Ma, J.; Zhang, Y.D.; Zhang, X.G. Spadicoides camelliae and Diplococcium livistonae, two new hyphomycetes on dead branches from Fujian Province, China. Mycoscience 2012, 53, 25–30. [Google Scholar] [CrossRef]
- Wu, W.P.; Diao, Y.Z. The chalara-like anamorphs of Leotiomycetes. Fungal Divers. 2023, 119, 213–490. [Google Scholar] [CrossRef]
- Vinjusha, N.; Kumar, T.K.A. Revision of Ganoderma species associated with stem rot of coconut palm. Mycologia 2022, 114, 157–174. [Google Scholar] [CrossRef]
- Piepenbring, M.; Nold, F.; Trampe, T.; Kirschner, R. Revision of the genus Graphiola (Exobasidiales, Basidiomycota). Nova Hedwig. 2012, 94, 67–96. [Google Scholar] [CrossRef]
- Zak, J.C.; Willig, M.R. Fungal biodiversity patterns. In Biodiversity of Fungi: Inventory and Monitoring Methods, 1st ed.; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Academic Press: San Diego, CA, USA, 2004; pp. 59–75. [Google Scholar]
- DeLong, D.C. Defining biodiversity. Wildl. Soc. Bull. 1996, 24, 738–749. [Google Scholar]
- Hyde, K.D.; Bussaban, B.; Paulus, B.; Crous, P.W.; Lee, S.; McKenzie, E.H.C.; Photita, W.; Lumyong, S. Diversity of saprobic microfungi. Biodivers. Conserv. 2007, 16, 7–35. [Google Scholar] [CrossRef]
- Mueller, G.M.; Schmit, J.P. Fungal biodiversity: What do we know? What can we predict? Biodivers. Conserv. 2007, 16, 1–5. [Google Scholar] [CrossRef]
- Schmit, J.P.; Mueller, G.M. An estimate of the lower limit of global fungal diversity. Biodivers. Conserv. 2007, 16, 99–111. [Google Scholar] [CrossRef]
- Wu, B.; Hussain, M.; Zhang, W.W.; Stadler, M.; Liu, X.Z.; Xiang, M.C. Current insights into fungal species diversity and perspective on naming the environmental DNA sequences of fungi. Mycology 2019, 10, 127–140. [Google Scholar] [CrossRef]
- Fries, E.M. Systema orbis vegetabilis. Primas Lineas Novae Constructionis Periclitatur Elias Fries. Pars I. Plantae Homonemeae; Lundae e Typographia Academia: Lund, Sweden, 1825. [Google Scholar]
- Bisby, G.R.; Ainsworth, G.C. The numbers of fungi. Trans. Br. Mycol. Soc. 1943, 26, 16–19. [Google Scholar] [CrossRef]
- Martin, G.W. The numbers of fungi. Proc. Iowa Acad. Sci. 1951, 58, 175–178. [Google Scholar]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed]
- Cannon, P.F. Diversity of the Phyllachoraceae with special reference to the tropics. In Biodiversity of Tropical Microfungi; Hyde, K.D., Ed.; Hong Kong University Press: Hong Kong, China, 1997; pp. 255–278. [Google Scholar]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Fungal diversity and its implications for genetic resource collections. Stud. Mycol. 2004, 50, 9–17. [Google Scholar]
- Hawksworth, D.L. Global species numbers of fungi: Are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers. Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Bass, D.; Richards, T.A. Three reasons to re-evaluate fungal diversity “on Earth and in the ocean”. Fungal Biol. Rev. 2011, 25, 159–164. [Google Scholar] [CrossRef]
- Blackwell, M. The fungi: 1, 2, 3… 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 1–32. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjoller, R.; Koljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers—A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Jayawardena, R.S.; Hyde, K.D. Hurdles in fungal taxonomy: Effectiveness of recent methods in discriminating taxa. Megataxa 2020, 1, 114–122. [Google Scholar] [CrossRef]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Koljalg, U.; Abarenkov, K. Identifying the ‘unidentified’ fungi: A global-scale long–read third–generation sequencing approach. Fungal Divers. 2020, 103, 273–293. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zinger, L.; Nilsson, R.H.; Kennedy, P.G.; Yang, T.; Anslan, S.; Mikryukov, V. Best practices in metabarcoding of fungi: From experimental design to results. Mol. Ecol. 2022, 31, 2769–2795. [Google Scholar] [CrossRef] [PubMed]
- Gautam, A.K.; Verma, R.K.; Avasthi, S.; Sushma; Bohra, Y.; Devadatha, B.; Niranjan, M.; Suwannarach, N. Current insight into traditional and modern methods in fungal diversity estimates. J. Fungi 2022, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Vetrovsky, T.; Lepinay, C.; Kohout, P. High-throughput sequencing view on the magnitude of global fungal diversity. Fungal Divers. 2022, 114, 539–547. [Google Scholar] [CrossRef]
- Baldrian, P.; Kohout, P.; Větrovský, T. Global fungal diversity estimated from high-throughput sequencing. In Evolution of Fungi and Fungal-Like Organisms; Pöggeler, S., James, T., Eds.; Springer Cham: Aargau, Switzerland, 2023; pp. 227–238. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Ohman, A.; Glotzer, D.; Nuhn, M.; Kirk, P.; Nilsson, R.H. Progress in molecular and morphological taxon discovery in Fungi and options for formal classification of environmental sequences. Fungal Biol. Rev. 2011, 25, 38–47. [Google Scholar] [CrossRef]
- Ryberg, M.; Nilsson, R.H. New light on names and naming of dark taxa. Mycokeys 2018, 30, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Ryberg, M.; Wurzbacher, C.; Tedersoo, L.; Anslan, S.; Polme, S.; Spirin, V.; Mikryukov, V.; Svantesson, S.; Hartmann, M.; et al. How, not if, is the question mycologists should be asking about DNA-based typification. Mycokeys 2023, 96, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 Million Species. Microbiol. Spectr. 2017, 5, 17. [Google Scholar] [CrossRef]
- Lücking, R.; Hawksworth, D.L. Formal description of sequence-based voucherless Fungi: Promises and pitfalls, and how to resolve them. IMA Fungus 2018, 9, 143–165. [Google Scholar] [CrossRef] [PubMed]
- Cheek, M.; Lughadha, E.N.; Kirk, P.; Lindon, H.; Carretero, J.; Looney, B.; Douglas, B.; Haelewaters, D.; Gaya, E.; Llewellyn, T.; et al. New scientific discoveries: Plants and fungi. Plants People Planet 2020, 2, 371–388. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Rossman, A.Y. Where are all the undescribed fungi? Phytopathology 1997, 87, 888–891. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D. Where are the missing fungi? Does Hong Kong have any answers? Mycol. Res. 2001, 105, 1514–1518. [Google Scholar] [CrossRef]
- Tang, A.M.C.; Shenoy, B.D.; Hyde, K.D. Fungal diversity. In Reconstructing the Tree of Life: Taxonomy and Systematics of Species Rich Taxa; Hodkinson, T.R., Parnell, J.A.N., Eds.; CRC Press: Boca Raton, FL, USA, 2007; Volume 72, pp. 221–249. [Google Scholar]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.; Phillips, A.J.; Jones, E.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Edwards, J.; Taylor, P.W.J.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef] [PubMed]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal Diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, E.B.G.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal Diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef]
- Cannon, P.F.; Hawksworth, D.L. The diversity of fungi associated with vascular plants: The known, the unknown and the need to bridge the knowledge gap. In Advances in Plant Pathology; Andrews, J.H., Tommerup, I.C., Eds.; Academic Press: Cambridge, MA, USA, 1995; Volume 11, pp. 277–302. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, US National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 15 April 2022).
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 15 April 2022).
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal Diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Hyde, K.D. Measuring biodiversity of microfungi in the wet tropics of North Queensland. In Proceedings of the Measuring and Monitoring Biodiversity of Tropical and Temperate Forests, Chiang Mai, Thailand, 27 August–2 September 1994; Boyle, T.J.B., Boontawee, B., Eds.; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 1995; pp. 271–286. [Google Scholar] [CrossRef]
- Hyde, K.D. Biodiversity of microfungi in North Queensland. Aust. Syst. Bot. 1996, 9, 261–271. [Google Scholar] [CrossRef]
- Zhou, D.Q.; Hyde, K.D. Host-specificity, host-exclusivity, and host-recurrence in saprobic fungi. Mycol. Res. 2001, 105, 1449–1457. [Google Scholar] [CrossRef]
- Shivas, R.G.; Hyde, K.D. Biodiversity of plant pathogenic fungi in the tropics. In Biodiversity of Tropical Microfungi; Hyde, K.D., Ed.; Hong Kong University Press: Hong Kong, China, 1997; pp. 47–56. [Google Scholar]
- Holliday, P. A Dictionary of Plant Pathology, 2nd ed.; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Lucas, J.A. Plant Pathology and Plant Pathogens, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 1998. [Google Scholar]
- Li, J.M.; Cornelissen, B.; Rep, M. Host-specificity factors in plant pathogenic fungi. Fungal Genet. Biol. 2020, 144, 11. [Google Scholar] [CrossRef]
- Hyde, K.D. Non-lichenised Australian ascomycetes. Aust. Syst. Bot. 2001, 14, 357–375. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Ariyawansa, H.A.; Phillips, A.J.L.; Wanasinghe, D.N.; Bhat, D.J.; McKenzie, E.H.C.; Singtripop, C.; Camporesi, E.; Hyde, K.D. Additions o Sporormiaceae: Introducing two novel genera, Sparticola and Forliomyces, from Spartium. Cryptogam. Mycol. 2016, 37, 75–97. [Google Scholar] [CrossRef]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Besitulo, A.; Sarma, V.V.; Hyde, K.D. Mangrove fungi from Siargao Island, Philippines. In Fungi in Marine Environments; Hyde, K.D., Ed.; Fungal Diversity Press: Hong Kong, China, 2002; pp. 267–283. [Google Scholar]
- Tomlinson, P.B. The Structural Biology of Palms; Oxford University Press: Clarendon, NY, USA, 1990. [Google Scholar]
- Pirozynski, K.A.; Weresub, L.K. A biogeographic view of the history of ascomycetes and the development of their pleomorphism. In The Whole Fungus: The Sexual and Asexual Synthesis, Alberta, Canada; Kendrick, B., Ed.; National Museum of Natural Sciences: Ottawa, ON, Canada, 1979; Volume I, pp. 93–123. [Google Scholar] [CrossRef]
- May, R.M. A fondness for fungi. Nature 1991, 352, 475–476. [Google Scholar] [CrossRef]
- Dowe, J. Extra-tropical palms: A statistical overview. Palm Enthus. 1992, 9, 4–8. [Google Scholar]
- Couvreur, T.L.P.; Forest, F.; Baker, W.J. Origin and global diversification patterns of tropical rain forests: Inferences from a complete genus-level phylogeny of palms. BMC Biol. 2011, 9, 12. [Google Scholar] [CrossRef]
- Dhillon, B.; Graham, F.; Laughinghouse, D.; Chakrabarti, S. Assessment of endophytic fungal community in palm leaves and their biocontrol potential. PhytoFront 2023. [Google Scholar] [CrossRef]
Figure 1.
World sampling regions of palm trees for associated fungi based on the available literature. Studies prior to 1950 were not considered when constructing the map, as most of the information is difficult to access. The regions/countries where palm trees were collected are named and delimited with coloured blocks and referred to in the chart legend. Extensively sampled regions are those where most studies have been carried out to specifically analyse palm fungi. Moderately and occasionally sampled regions are those where the reports of palm fungi were a consequence of occasional taxonomic or broader studies. Many occasionally sampled regions have not been mapped, as they are island countries that are barely visible due to the scale of the map. These include several Antilles islands of the Caribbean (Barbados, Dominica, Grenada, Guadeloupe, French West Indies, and Virgin Islands), Bermuda, African island countries (Comoro Islands, Mauritius, São Tomé and Príncipe, Réunion, and Seychelles), the Andaman and Nicobar Islands, and several islands in the Pacific Ocean (Cook Islands, French Polynesia, Niue, Samoa, and Tonga in Polynesia, Fiji, Vanuatu, and Solomon Islands in Melanesia, and Guam and Kiribati in Micronesia, Oceania). Figure source: created with a template available in the webserver for MapChart (https://www.mapchart.net/, accessed on 15 September 2023).
Figure 1.
World sampling regions of palm trees for associated fungi based on the available literature. Studies prior to 1950 were not considered when constructing the map, as most of the information is difficult to access. The regions/countries where palm trees were collected are named and delimited with coloured blocks and referred to in the chart legend. Extensively sampled regions are those where most studies have been carried out to specifically analyse palm fungi. Moderately and occasionally sampled regions are those where the reports of palm fungi were a consequence of occasional taxonomic or broader studies. Many occasionally sampled regions have not been mapped, as they are island countries that are barely visible due to the scale of the map. These include several Antilles islands of the Caribbean (Barbados, Dominica, Grenada, Guadeloupe, French West Indies, and Virgin Islands), Bermuda, African island countries (Comoro Islands, Mauritius, São Tomé and Príncipe, Réunion, and Seychelles), the Andaman and Nicobar Islands, and several islands in the Pacific Ocean (Cook Islands, French Polynesia, Niue, Samoa, and Tonga in Polynesia, Fiji, Vanuatu, and Solomon Islands in Melanesia, and Guam and Kiribati in Micronesia, Oceania). Figure source: created with a template available in the webserver for MapChart (https://www.mapchart.net/, accessed on 15 September 2023).

Figure 2.
Percentage of fungal overlapping between the four peat swamp palm species in ecological studies conducted in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand (adapted from [669]).
Figure 2.
Percentage of fungal overlapping between the four peat swamp palm species in ecological studies conducted in the Sirindhom Peat Swamp Forest, Narathiwat, Thailand (adapted from [669]).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pereira, D.S.; Phillips, A.J.L. Palm Fungi and Their Key Role in Biodiversity Surveys: A Review. J. Fungi 2023, 9, 1121. https://doi.org/10.3390/jof9111121
AMA Style
Pereira DS, Phillips AJL. Palm Fungi and Their Key Role in Biodiversity Surveys: A Review. Journal of Fungi. 2023; 9(11):1121. https://doi.org/10.3390/jof9111121
Chicago/Turabian StylePereira, Diana S., and Alan J. L. Phillips. 2023. "Palm Fungi and Their Key Role in Biodiversity Surveys: A Review" Journal of Fungi 9, no. 11: 1121. https://doi.org/10.3390/jof9111121
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.