
The Identity, Virulence, and Antifungal Effects of the Didymellacesous Fungi Associated with the Rapeseed Blackleg Pathogen Leptosphaeria biglobosa
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultural Media
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analysis
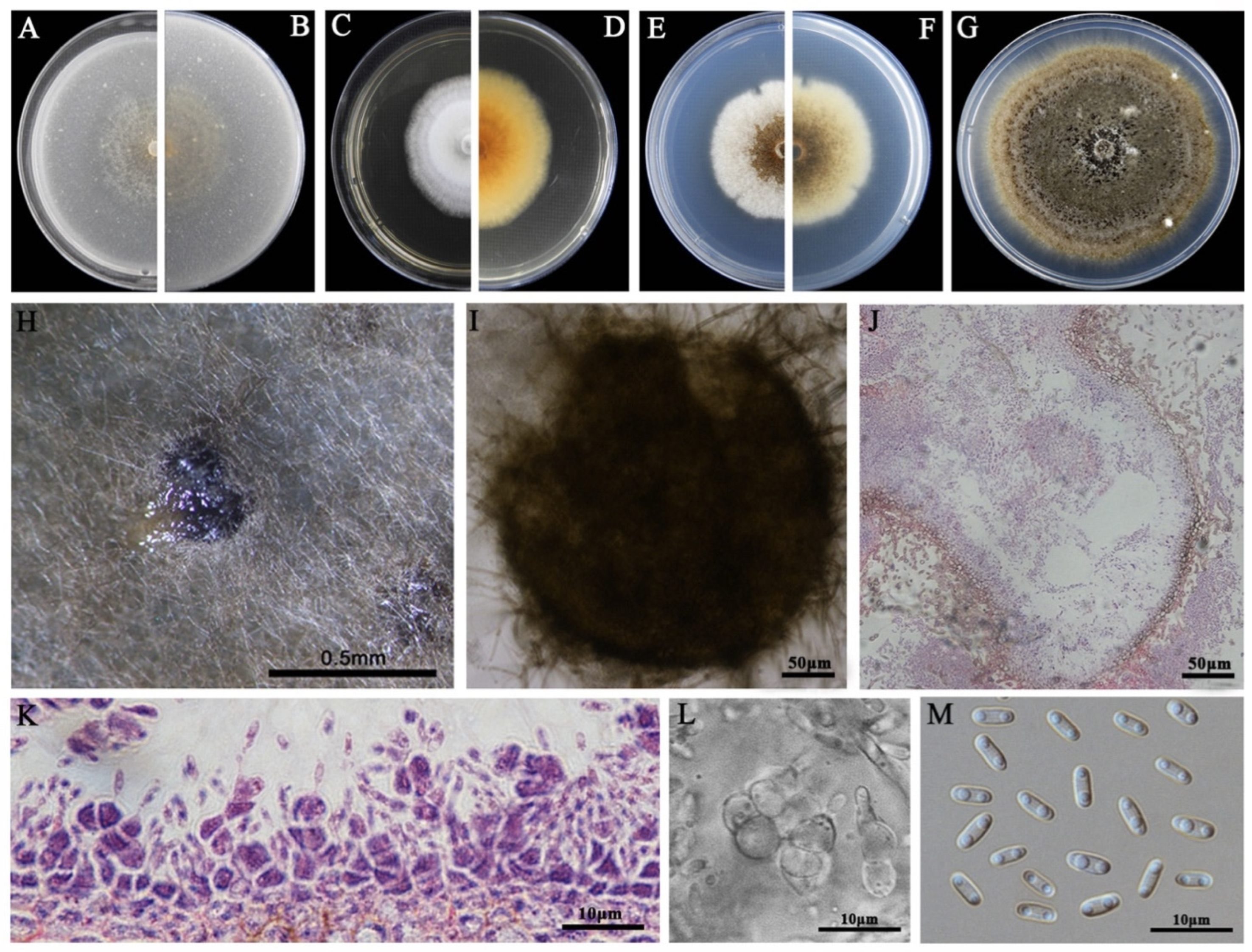
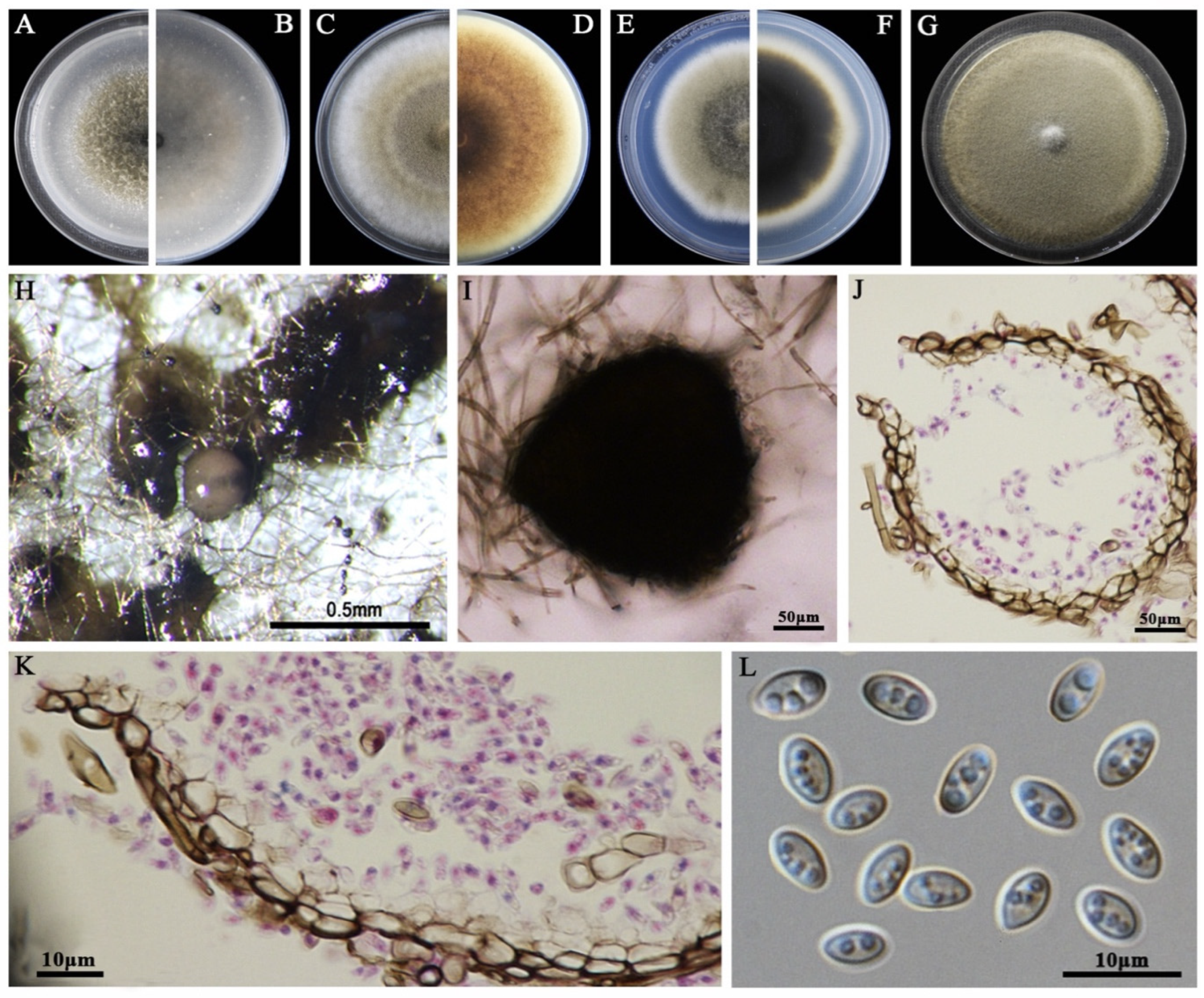
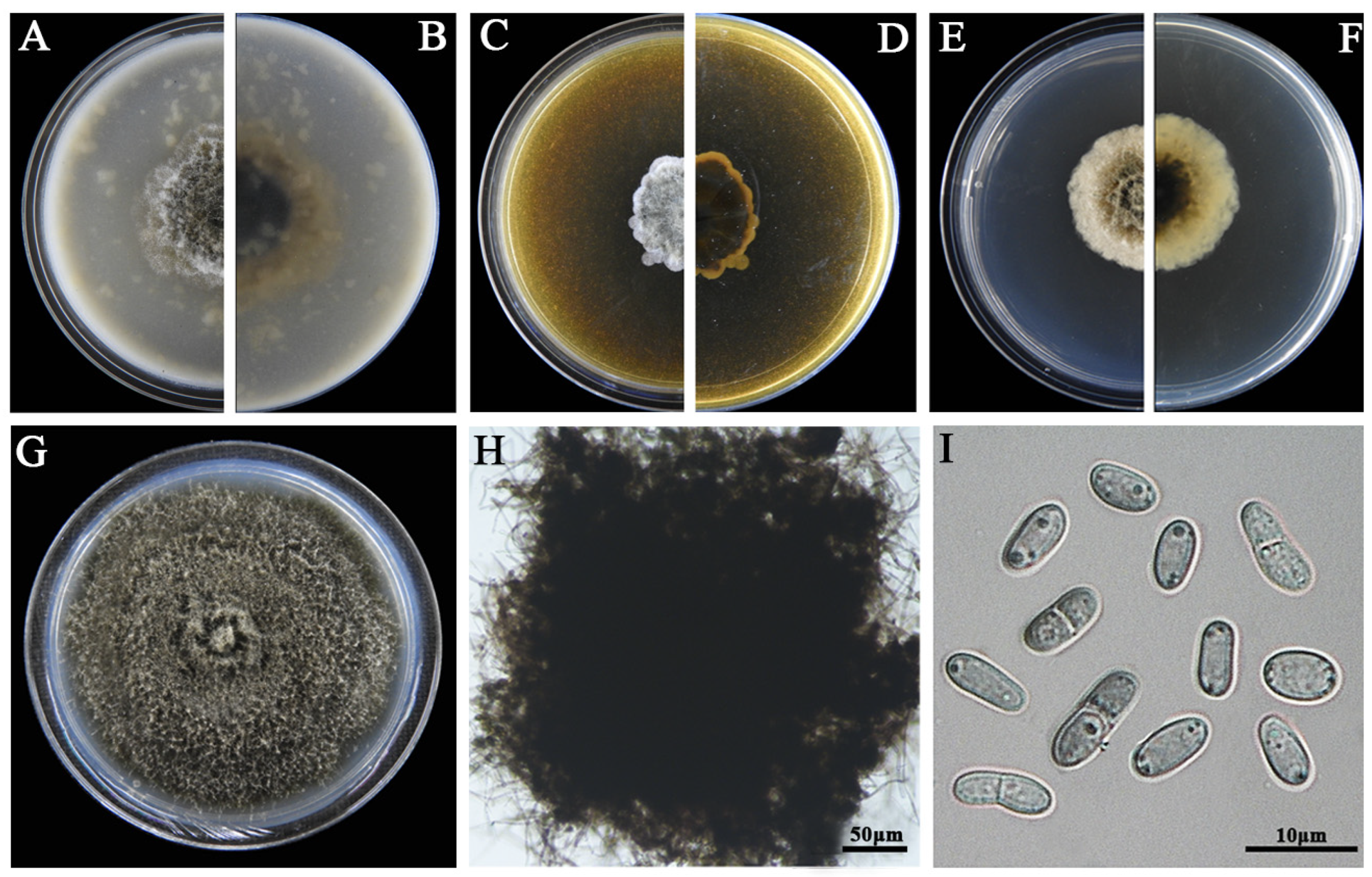
2.4. Morphological Characterization
2.5. Virulence Determination
2.6. Co-Inoculation Assays
2.7. Suppression of Fungal Infection by Cultural Filtrates of LbR Strains
2.8. Determination of the Antifungal Activity of P2 Cultural Filtrate
2.9. Determination of the Antifungal Activity of P2 Pycnidiospore Suspensions
2.10. The Extraction and Purification of Antifungal Metabolites
2.11. Identification of Penicillither
2.12. The Quantification of Penicillither
2.13. Data Analysis
3. Results
3.1. The Species Identity of the LbRs
3.2. Morphological Characteristics
3.3. Virulence of the LbRs on Rapeseed
3.4. The Effect of the LbRs on Infection by L. biglobosa
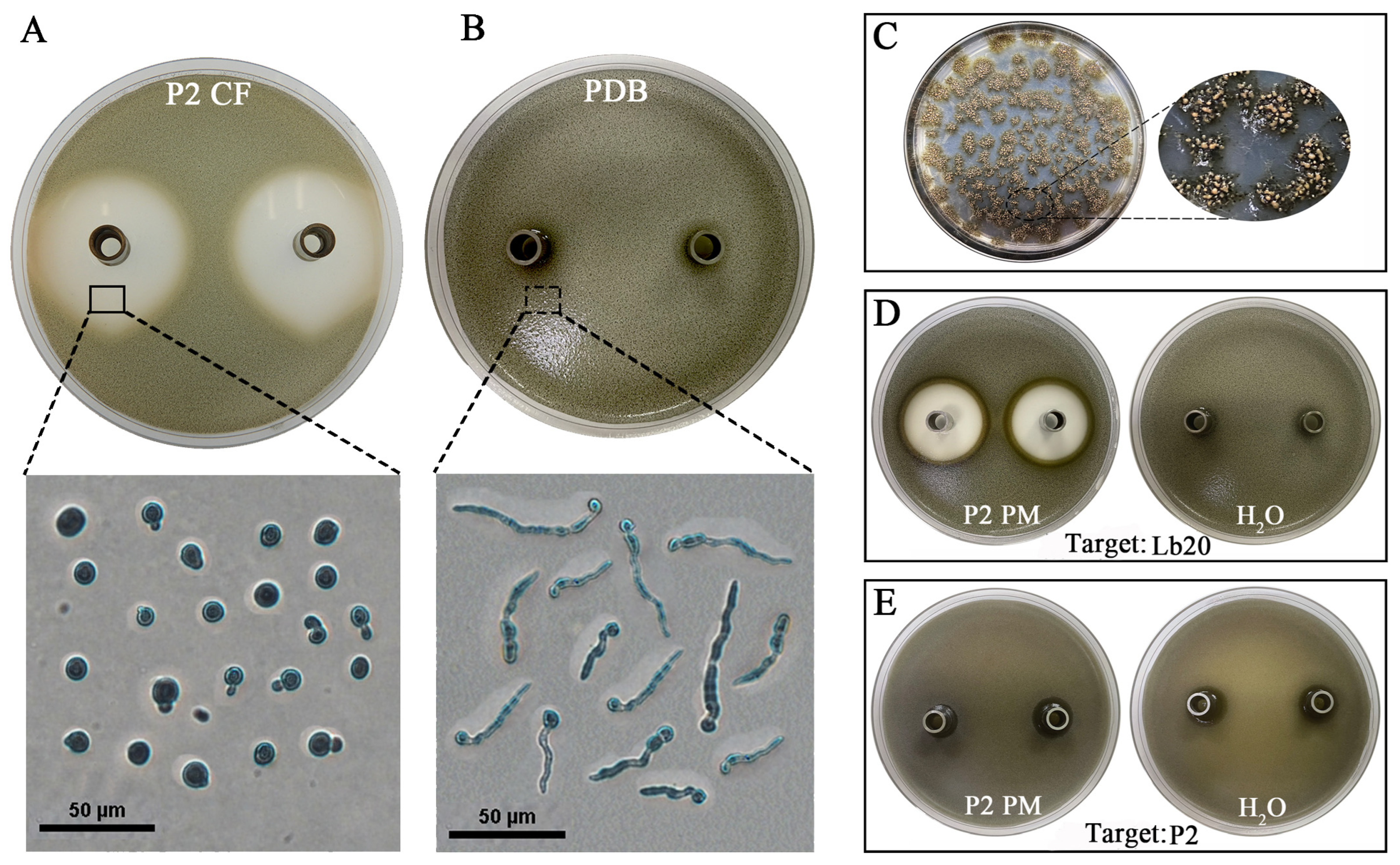
3.5. The Efficacy of Cultural Filtrates of the LbRs in the Suppression of L. biglobosa Infection
3.6. The Antifungal Activity of P2 CF and P2 PS
3.7. Penicillither Yield and Antifungal Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, H.Z. New demand-oriented rapeseed industry developing strategy. Chin. J. Oil Crop Sci. 2018, 40, 613–617. [Google Scholar]
- Zheng, X.; Koopmann, B.; Ulber, B.; von Tiedemann, A. A global survey on diseases and pests in oilseed rape-current challenges and innovative strategies of control. Front. Agron. 2020, 2, 590908. [Google Scholar] [CrossRef]
- Liban, S.H.; Cross, D.J.; Kutcher, H.R.; Peng, G.; Fernando, W.G.D. Race structure and frequency of avirulence genes in the western Canadian Leptosphaeria maculans pathogen population, the causal agent of blackleg in brassica species. Plant Pathol. 2016, 65, 1161–1169. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Huang, Y.J.; Bosch, F.V.D.; West, J.S. Coexistence of related pathogen species on arable crops in space and time. Annu. Rev. Phytopathol. 2006, 44, 163–182. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynsk, B.; Tudzynski, P.; van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- McLaren, D.L.; Huang, H.C.; Rimmer, S.R. Control of apothecial production of Sclerotinia sclerotiorum by Coniothyrium minitans and Talaromyces flavus. Plant Dis. 1996, 80, 1373–1378. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Tewari, J.P.; Shinners, T.C.; Briggs, K.G. Production of calcium oxalate crystals by two species of Cyathus in culture and in infested plant debris. Verl. Z. Für Naturforschung 1997, 52, 421–425. [Google Scholar] [CrossRef]
- Danielsson, J.; Reva, O.; Meijer, J. Protection of rapeseed (Brassica napus) toward fungal pathogens by strains of plant-associated Bacillus amyloliquefaciens. Microbial Ecol. 2007, 54, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Abuamsha, R.; Salman, M.; Ehlers, R.U. Effect of seed priming with Serratia plymuthica and Pseudomonas chlororaphis to control Leptosphaeria maculans in different rapeseed cultivars. Eur. J. Plant Pathol. 2011, 130, 287–295. [Google Scholar] [CrossRef]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identifcation Manual; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- Aveskamp, M.M.; de Gruyter, J.; Crous, P.W. Biology and recent developments in the systematics of Phoma, a complex genus of major quarantine signifcance. Fungal Divers. 2008, 31, 1–18. [Google Scholar]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Xu, B.G.; Wang, J.Y.; Su, Z.Z.; Lin, F.C.; Zhang, C.L.; Kubicek, C.P. Bioactive metabolites from Phoma species, an endophytic fungus from the Chinese medicinal plant Arisaema erubescens. Appl. Microbiol. Biotechnol. 2011, 93, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Arie, T.; Hobayashi, Y.; Okada, G.; Kono, Y.; Yamaguchi, I. Control of soilborne clubroot disease of cruciferous plants by epoxydon from Phoma glomerata. Plant Pathol. 1998, 47, 743–748. [Google Scholar] [CrossRef]
- Arie, T.; Kobayashi, Y.; Kono, Y.; Okada, G.; Yamaguchi, I. Control of clubroot of crucifers by Phoma glomerata and its product. Pest Sci. 1999, 55, 566–614. [Google Scholar] [CrossRef]
- Graupner, P.R.; Carr, A.; Clancy, E.; Gilbert, J.; Bailey, K.L.; Derby, J.-A.; Gerwick, B.C. The macrocidins: Novel cyclic tetramic acids with herbicidal activity produced by Phoma macrostoma. J. Nat. Prod. 2003, 66, 1558–1561. [Google Scholar] [CrossRef]
- Sullivan, R.F.; White, J.F.J.R. Phoma glomerata as a mycoparasite of powdery mildew. Appl. Environ. Microb. 2000, 66, 425–427. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef]
- Strobel, G.; Singh, S.K.; Riyaz-UI-Hassan, S.; Mitchell, A.M.; Geary, B.; Sears, J. An endophytic/pathogenic Phoma sp. from creosote bush producing biologically active volatile compounds having fuel potential. FEMS Microbiol. Lett. 2011, 320, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, J.C.; Lyv, X.; Xu, J.W.; Wu, M.D.; Zhang, J.; Yang, L.; Li, G.Q. Large-scale surveys of blackleg of rapeseed (Leptosphaeria biglobosa) revealed new insights into epidemics of this disease in China. Plant Dis. 2023, 107, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species associated with anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Evol. Genet. Anal. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P.; et al. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModelTest v. 2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Swofford, D. PAUP 4.0 b10: Phylogenetic analysis using parsimony (*and other methods). In Computer Programme; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree, v. 1.4.2. Institute of Evolutionary Biology. University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 June 2020).
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.H.; Hwang, B.P.; Kim, H.S.; Lee, J.J. Fungal metabolites, asterric acid derivatives inhibit vascular endothelial growth factor (VEGF)-induced tube formation of HUVECs. J. Antibiot. 2002, 55, 552–556. [Google Scholar] [CrossRef]
- Chen, W.J. Studies on Antibacterial Ingredients from the Marine Fungus Aspergillus flavipes. Master’s Thesis, Harbin University of Commerce, Harbin, China, 2020. [Google Scholar]
- Baird, R.E. First report of Phoma macrostoma causing leaf spot on fevertree. Plant Dis. 1993, 77, 1168. [Google Scholar] [CrossRef]
- Evans, H.C.; Seier, M.K.; Derby, J.-A.; Falk, S.; Bailey, K.L. Tracing the origins of white tip disease of Cirsium arvense and its causal agent, Phoma macrostoma. Weed Res. 2013, 53, 42–52. [Google Scholar] [CrossRef]
- Caesar, A.J.; Lartey, R.T.; Caesar-Ton-That, T.C. First report of a root and crown disease of the invasive weed Lepidium draba caused by Phoma macrostoma. Plant Dis. 2012, 96, 145. [Google Scholar] [CrossRef] [PubMed]
- Chohan, J.S.; Chand, T. Phoma glomerata, a new pathogen om pears (Pyrus communis). Trans. Br. Mycol. Society 1980, 75, 509. [Google Scholar] [CrossRef]
- Moral, J.; Lichtemberg, P.S.F.; Papagelis, A.; Sherman, J.; Michailides, T.J. Didymella glomerata causing leaf blight on pistachio. Eur. J. Plant Pathol. 2018, 151, 1095–1099. [Google Scholar] [CrossRef]
- Boerema, G.H.; Loerakker, W.M.; Wittern, I. Zum Auftreten von Phoma nigrificans (P. Karst.) comb. nov. (teleomorph Didymella macropodii Petrak) an Winterraps (Brassica napus L. var. oleifera Metzger). J. Phytopathol. 1986, 115, 167–273. [Google Scholar] [CrossRef]
- Schaffrath, U.; Vogel, L.; Loehrer, M. First report of Boeremia exigua var. exigua causing black spot-like symptoms on commercially grown soybean in Germany. Plant Dis. 2021, 105, 219. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Hay, F.S.; Gorny, A. Spatiotemperal attributes and crop loss associated with tan spot epidemics in baby lima bean in New York. Plant Dis. 2018, 102, 405–412. [Google Scholar] [CrossRef]
- Li, Y.P.; You, M.P.; Finnegan, P.M.; Khan, T.N.; Lanoiselet, V.; Eyres, N.; Barbetti, M.J. First report of black spot caused by Boeremia exigua var. exigua on field pea in Australia. Plant Dis. 2012, 96, 148. [Google Scholar]
- Thomidis, T.; Zambounis, A.; Prodromou, I. Occurrence of a postharvest fruit rot disease of apples caused by Boeremia exigua var. exigua in Ptolemaida Kozani, Greece. Plant Dis. 2016, 100, 2333. [Google Scholar]
- Cai, F.F.; Yang, C.D.; Ma, T.; Jin, M.J.; Cui, L.X. First report of Boeremia exigua var. exigua causing branch blight on walnut in China. Plant Dis. 2021, 105, 3291. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ma, Y.P.; Wang, J.W.; Wei, X.Y. Antibacterial substances of endophytic fungus isolated from Pueraria lbata. J. Shaanxi Norm. Univ. 2014, 42, 71–74. [Google Scholar]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Lee, I.-J. Endophytic fungi produce gibberellins and indoleacetic acidand promotes host-plant growth during stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef] [PubMed]
- de Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Redisposition of Phoma-like anamorphs in Pleosporales. Stud. Mycol. 2012, 75, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Latunde-Dada, A.O.; Hall, A.M.; Fitt, B.D.L. Phoma stem canker disease on oilseed rape (Brassica napus) in China is caused by Leptosphaeria biglobosa ‘brassicae’. Eur. J. Plant Pathol. 2014, 140, 841–857. [Google Scholar] [CrossRef]
- Cai, X.; Huang, Y.J.; Jiang, D.H.; Fitt, B.D.L.; Li, G.Q.; Yang, L. Evaluation of oilseed rape seed yield losses caused by Leptosphaeria biglobosa in central China. Eur. J. Plant Pathol. 2018, 150, 179–190. [Google Scholar] [CrossRef]
- Luo, T.; Li, G.Q.; Yang, L. First report of Leptosphaeria biglobosa ‘canadensis’ causing blackleg on oilseed rape (Brassica napus) in China. Plant Dis. 2021, 105, 3760. [Google Scholar] [CrossRef]
- Nathaniels, N.Q.R.; and Taylor, G.S. Latent infection of winter oilseed rape by Leptosphaeria maculans. Plant Pathol. 1983, 32, 23–31. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Zhang, J.; Yang, L.; Zhang, L.; Jiang, D.H.; Chen, W.D.; Li, G.Q. Diversity and biocontrol potential of endophytic fungi in Brassica napus. Biol. Control 2014, 72, 98–108. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [PubMed]
- De Lamo, F.J.; Takken, F.L.W. Biocontrol by Fusarium oxysporum using endophyte mediated resistance. Front. Plant Sci. 2020, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Pitt, W.M.; Bailey, K.L.; Fu, Y.B.; Peterson, G.W. Biological and genetic characterisation of Phoma macrostoma isolates with bioherbicidal activity. Biocontrol Sci. Technol. 2012, 22, 813–835. [Google Scholar] [CrossRef]
- Kemkuignou, B.M.; Treiber, L.; Zeng, H.X.; Schrey, H.; Schobert, R.; Stadler, M. Macrooxazoles A-D, new 2,5-disubstituted oxazole-4-carboxylic acid derivatives from the plant pathogenic fungus Phoma macrostoma. Molecules 2020, 25, 5497. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Satpradit, S.; Klaiklay, S.; Phongapaichit, S.; Borwornwiriyapan, K.; Sakayaroj, J. Polyketide anthraquinone, diphenyl ether, and xanthone derivatives from the soil fungus Penicillium sp. PSU-RSPG99. Tetrahedron 2014, 70, 5148–5152. [Google Scholar] [CrossRef]
- Qin, J.; Lyu, A.; Zhang, Q.H.; Yang, L.; Zhang, J.; Wu, M.D.; Li, G.Q. Strain identification and metabolites isolation of Aspergillus capensis CanS-34A from Brassica napus. Mol. Biol. Rep. 2019, 46, 3451–3460. [Google Scholar] [CrossRef]
- Curtis, R.F.; Hassalld, C.H.; Jones, D.W.; Williams, T.W. The biosynthesis of phenols. Part II. Asterric acid, a metabolic product of Aspergillus terreus Thorn. J. Chem. Soc. 1960, 940, 4838–4842. [Google Scholar] [CrossRef]
- Jayasuriya, H.; Ball, R.G.; Zink, D.L.; Smith, J.L.; Goetz, M.A.; Jenkins, R.G.; Nallin-Omstead, M.; Silverman, K.C.; Bills, G.F.; Lingham, R.B.; et al. Barceloneic acid A, a new farnesyl-protein transferase inhibitor from a Phoma species. J. Nat. Prod. 1995, 58, 986–991. [Google Scholar] [CrossRef]
- de Hoog, G.S.; Gerrits van den Ende, A.H. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.; Aveskamp, M.M.; de Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia 2009, 22, 56–62. [Google Scholar] [CrossRef]
- White, T.J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Wiley: Hoboken, NJ, USA, 1990; pp. 315–322. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Strain | Collection Place | GenBank Accession Number | |||
|---|---|---|---|---|---|
| ITS1 1 | LSU | tub2 | rpb2 | ||
| L. biglobosa Lb20 | Wuhan, Hubei Province, China, 2017 | MT807903 | MT611125 | MT807904 | MT807905 |
| L. biglobosa Lb982 | Anshun, Guizhou Province, China, 2018 | MT611111 | MT611126 | MT650708 | MT683499 |
| L. biglobosa Lb983 | Anshun, Guizhou Province, China, 2018 | MT611112 | MT611127 | MT650709 | MT683500 |
| L. biglobosa Lb1024 | Bijie, Guizhou Province, China, 2018 | MT611113 | MT611128 | MT650710 | MT683501 |
| L. biglobosa Lb1025 | Bijie, Guizhou Province, China, 2018 | MT611114 | MT611129 | MT650711 | MT683502 |
| L. biglobosa Lb1041 | Bijie, Guizhou Province, China, 2018 | MT611115 | MT611130 | MT650712 | MT683503 |
| L. biglobosa Lb3974 | Longnan, Gansu Province, China, 2019 | MT611116 | MT611131 | MT650713 | MT683504 |
| L. biglobosa Lb3995 | Longnan, Gansu Province, China, 2019 | MT611117 | MT611132 | MT650714 | MT683505 |
| L. biglobosa Lb4055 | Longnan, Gansu Province, China, 2019 | MT611118 | MT611133 | MT650715 | MT683506 |
| L. biglobosa Lb4068 | Longnan, Gansu Province, China, 2019 | MT611119 | MT611134 | MT650716 | MT683507 |
| L. biglobosa Lb4079 | Longnan, Gansu Province, China, 2019 | MT611120 | MT611135 | MT650717 | MT683508 |
| L. biglobosa Lb4099 | Longnan, Gansu Province, China, 2019 | MT611121 | MT611136 | MT650718 | MT683509 |
| L. biglobosa Lb4149 | Tianshui, Gansu Province, China, 2019 | MT611122 | MT611137 | MT650719 | MT683510 |
| L. biglobosa Lb4150 | Tianshui, Gansu Province, China, 2019 | MT611123 | MT611138 | MT650720 | MT683511 |
| L. biglobosa Lb4152 | Tianshui, Gansu Province, China, 2019 | MT611124 | MT611139 | MT650721 | MT683512 |
| Didymella sp. P1 | Enshi, Hubei Province, China, 2017 | MK121680 | MK121684 | MK160142 | MK127941 |
| D. macrostoma P2 | Yunxi, Hubei Province, China, 2017 | MK121681 | MK121685 | MK160143 | MK127942 |
| Didymella sp. P3 | Yunxi, Hubei Province, China, 2017 | MK121682 | MK121686 | MK160144 | MK127943 |
| D. glomerata P4 | Huzhu, Qinghai Province, China, 2017 | MK121683 | MK121687 | MK160145 | MK127944 |
| D. macropodii P5 | Huzhu, Qinghai Province, China, 2018 | MK579368 | MK579371 | MK579377 | MK579374 |
| D. macropodii P6 | Huzhu, Qinghai Province, China, 2018 | MK579369 | MK579372 | MK579378 | MK579375 |
| D. macropodii P7 | Huzhu, Qinghai Province, China, 2018 | MK579370 | MK579373 | MK579379 | MK579376 |
| Boeremia exigua P8 | Lincang, Yunnan Province, China, 2019 | MT505299 | MT505300 | MT505301 | MT505302 |
| Strain | Mycelial Growth Rate (mm/d) | Pycnidiospores | ||||
|---|---|---|---|---|---|---|
| Malt Extract Agar | Oatmeal Agar | Potato Dextrose Agar | Shape | Size in μm (n = 50) | ||
| Leptosphaeria biglobosa Lb20 | 0.7 ± 0.2 d 1 | 2.4 ± 0.4 c | 2.3 ± 0.1 b | Cylindroid | 4.6–5.3 (4.9) × 2.0–2.3 (2.1) | |
| Didymella sp. P1 | 2.2 ± 0.4 ab | 1.7 ± 0.1 d | 2.8 ± 0.3 b | Cylindroid | 4.7–5.3 (4.9) × 2.0–2.3 (2.1) | |
| Didymella macrostoma P2 | 2.6 ± 0.1 a | 3.5 ± 0.7 a | 2.7 ± 0.5 b | Ellipsoid | 7.3–9.3 (8.1) × 3.3–4.0 (3.6) | |
| Didymella sp. P3 | 2.2 ± 0.5 ab | 2.9 ± 0.3 b | 2.2 ± 0.5 b | Cylindroid | 4.0–4.7 (4.2) × 1.7–2.0 (1.8) | |
| Didymella glomerata P4 | 2.6 ± 0.3 a | 3.4 ± 0.3 a | 3.5 ± 0.7 a | Ellipsoid | 7.3–8.7 (8.1) × 4.0–4.7 (4.4) | |
| Didymella macropodii P5 | 1.8 ± 0.1 b | 2.3 ± 0.3 c | 1.6 ± 0.6 c | Allantoid | 7.2–9.6 (8.4) × 2.4–2.8 (2.6) | |
| Didymella macropodii P6 | 2.5 ± 0.6 a | 2.3 ± 0.4 c | 3.5 ± 0.9 a | Allantoid | 7.6–10.4 (8.6) × 2.8–3.5 (3.1) | |
| Didymella macropodii P7 | 2.6 ± 0.6 a | 3.8 ± 0.4 a | 3.9 ± 0.2 a | Allantoid | 6.3–11.1 (8.4) × 2.1–3.5 (2.7) | |
| Boremia exigua P8 | 1.2 ± 0.4 c | 2.0 ± 0.4 cd | 1.5 ± 0.2 c | Ellipsoid | 6.9–9.7 (8.0) × 3.5–5.2 (4.4) | |
| Strain | Cotyledon Lesion Diameter (mm) | Lesions on Stem Epidermis | Lesion in Stem Pith | ||||
|---|---|---|---|---|---|---|---|
| Length (mm) | Width (mm) | Length (mm) | Width (mm) | ||||
| Leptosphaeria biglobosa Lb20 | 8.2 ± 1.1 a 1 | 16.3 ± 2.6 a | 3.8 ± 0.8 a | 65.3 ± 6.6 a | 7.5 ± 0.8 a | ||
| Didymella sp. P1 | 1.1 ± 0.1 b | 2.0 ± 0.0 f | 1.0 ± 0.0 d | 2.0 ± 0.0 e | 1.0 ± 0.0 c | ||
| Didymella macrostoma P2 | 1.2 ± 0.1 b | 8.7 ± 0.8 cd | 2.7 ± 0.5 bc | 5.0 ± 0.9 e | 1.5 ± 0.5 c | ||
| Didymella sp. P3 | 1.2 ± 0.1 b | 9.2 ± 1.3 c | 2.5 ± 0.5 bc | 6.5 ± 0.5 e | 1.8 ± 0.8 c | ||
| Didymella glomerata P4 | 1.3 ± 0.1 b | 11.7 ± 4.1 b | 2.8 ± 0.4 b | 12.0 ± 2.4 d | 1.8 ± 0.8 c | ||
| Didymella macropodii P5 | 1.6 ± 0.3 b | 10.2 ± 1.7 bc | 2.8 ± 0.8 b | 30.8 ± 2.8 b | 6.2 ± 0.8 b | ||
| Didymella macropodii P6 | 1.4 ± 0.4 b | 9.3 ± 2.3 bc | 2.8 ± 1.0 b | 26.7 ± 4.1 b | 6.3 ± 0.8 b | ||
| Didymella macropodii P7 | 1.5 ± 0.3 b | 6.7 ± 0.8 de | 1.8 ± 0.8 c | 27.2 ± 3.8 b | 7.0 ± 1.1 ab | ||
| Boeremia exigua P8 | 1.7 ± 0.3 b | 6.0 ± 0.6 e | 2.3 ± 0.8 c | 20.5 ± 7.0 c | 7.0 ± 0.6 ab | ||
| Position | δH (m, J in Hz) 600 MHz | δC | |||
|---|---|---|---|---|---|
| This Study | [35] | This Study | [35] | ||
| 1 | 138.5 | 135.4 | |||
| 2 | 125.7 | 124.4 | |||
| 3 | 6.72, d | 6.73, d | 110.6 | 110.5 | |
| 4 | ND | 150.6 | |||
| 5 | 6.65, d | 6.65, d | 104.8 | 104.7 | |
| 6 | ND | 152.9 | |||
| 7 | 56.6 | 56.1 | |||
| 8 | ND | 164.5 | |||
| 9 | 51.9 | 51.6 | |||
| 1′ | ND | 153.4 | |||
| 2′ | 107.7 | 107.3 | |||
| 3′ | ND | 154.5 | |||
| 4′ | 6.51, s | 6.51, s | ND | 110.8 | |
| 5′ | 141.5 | 137.9 | |||
| 6′ | ND | 113.5 | |||
| 7′ | 2.27, s | 2.30, s | 20.5 | 20.2 | |
| 8′ | 167.3 | 164.6 | |||
| 9′ | ND | 51.8 | |||
| 3′-OH | 8.40, s | 9.95, br s | |||
| 7-OCH3 | 3.22, s | 3.23, s | |||
| 9-OCH3 | 3.62, s | 3.63 s | |||
| 9′-OCH3 | 3.61, s | 3.60, s | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, J.; Luo, T.; Wu, M.; Yang, L.; Chen, W.; Li, G.; Zhang, J. The Identity, Virulence, and Antifungal Effects of the Didymellacesous Fungi Associated with the Rapeseed Blackleg Pathogen Leptosphaeria biglobosa. J. Fungi 2023, 9, 1167. https://doi.org/10.3390/jof9121167
Cheng J, Luo T, Wu M, Yang L, Chen W, Li G, Zhang J. The Identity, Virulence, and Antifungal Effects of the Didymellacesous Fungi Associated with the Rapeseed Blackleg Pathogen Leptosphaeria biglobosa. Journal of Fungi. 2023; 9(12):1167. https://doi.org/10.3390/jof9121167
Chicago/Turabian StyleCheng, Junyu, Tao Luo, Mingde Wu, Long Yang, Weidong Chen, Guoqing Li, and Jing Zhang. 2023. "The Identity, Virulence, and Antifungal Effects of the Didymellacesous Fungi Associated with the Rapeseed Blackleg Pathogen Leptosphaeria biglobosa" Journal of Fungi 9, no. 12: 1167. https://doi.org/10.3390/jof9121167