
Three New Species of Microdochium (Microdochiaceae, Xylariales) on Bambusaceae sp. and Saprophytic Leaves from Hainan and Yunnan, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Fungal Isolation and Morphology
2.2. DNA Extraction, PCR and Sequencing
2.3. Phylogeny
3. Results
3.1. Phylogenetic Analyses

3.2. Taxonomy
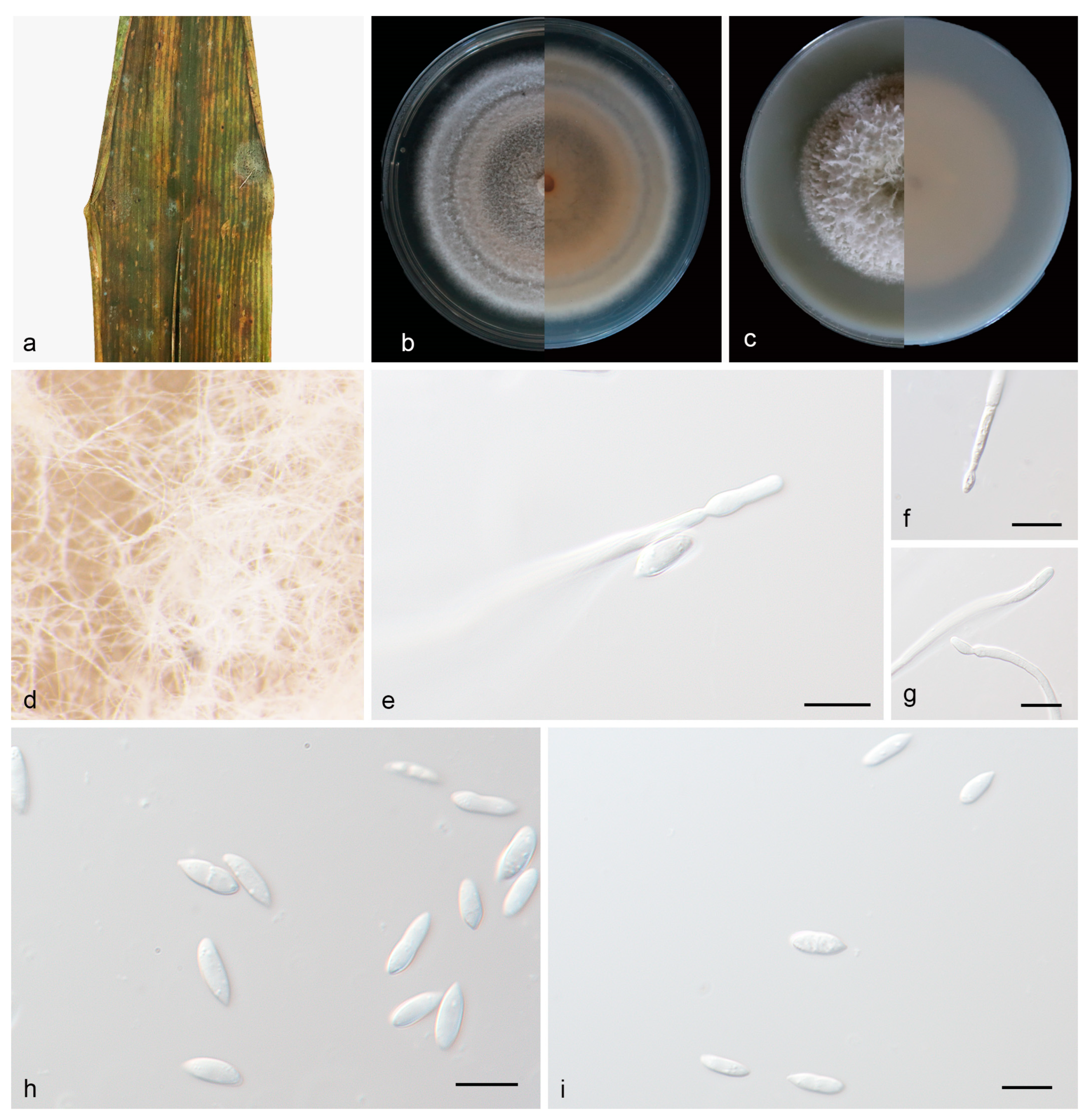
3.2.1. Microdochium bambusae J. Zhang, Z.X. Zhang, & Z. Li, sp. Nov.; Figure 2

3.2.2. Microdochium nannuoshanense J. Zhang, Z.X. Zhang, & Z. Li, sp. Nov.; Figure 3

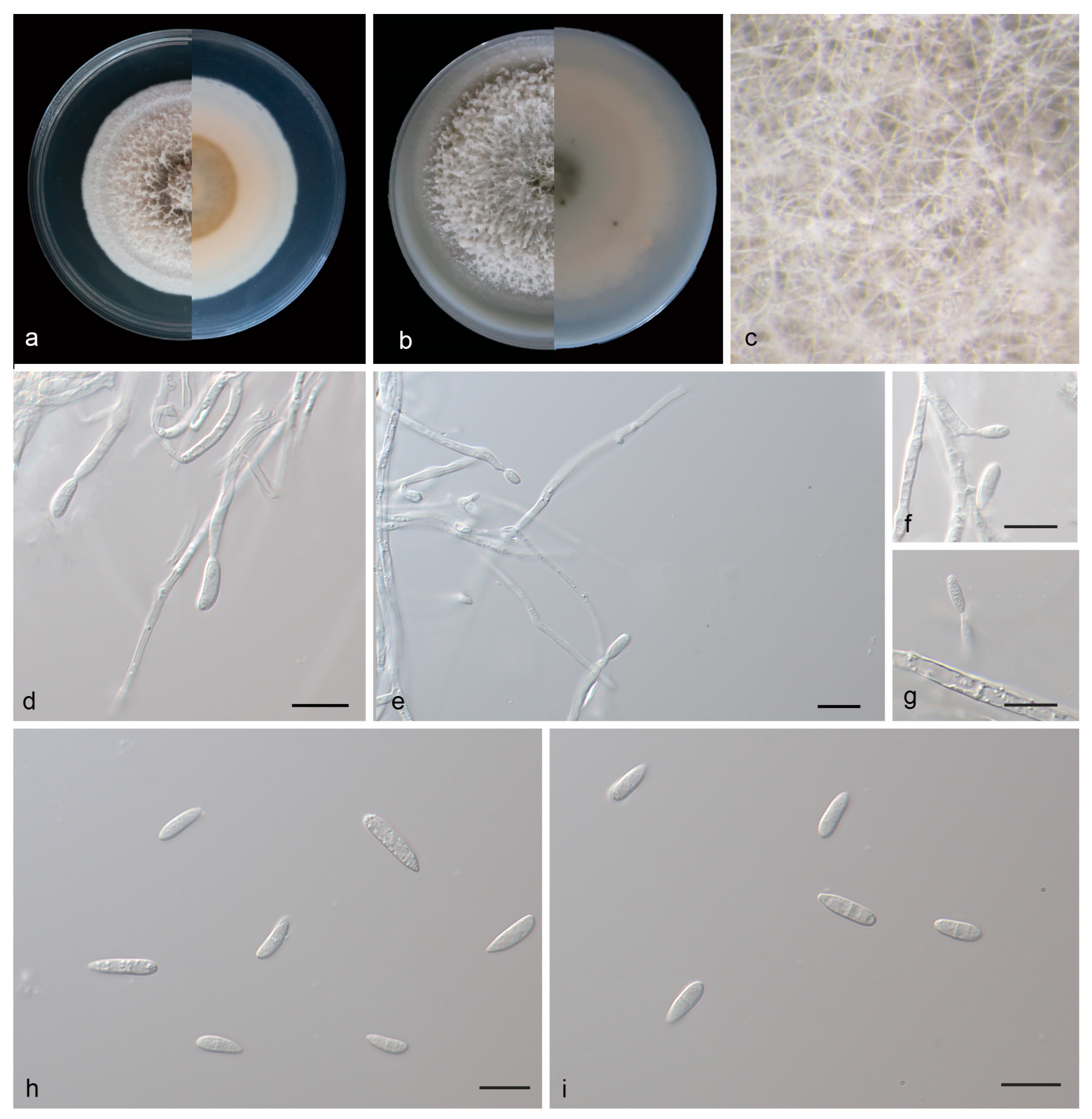
3.2.3. Microdochium phyllosaprophyticum J. Zhang, Z.X. Zhang, & Z. Li, sp. Nov.; Figure 4


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Conidiogenous Cells | Size of Conidiogenous Cells (μm) | Conidia | Size of Conidia (μm) | References |
|---|---|---|---|---|---|
| Microdochium albescens | Doliiform to obpyriform | 6–15 × 1.5–4 | Falcate, apex pointed | 11–16 × 3.5–4.5 | [13] |
| M.bambusae | Cylindrical and smooth | 17.4–30.0 × 2.5–3.0 | Oblong to ellipsoid | 13.0–17.0 × 2.5–3.5 | This study |
| M. bolleyi | Ampullate or cylindrical | 3.1–6.4 × 2.5–3.8 | Crescent, lunate | 5.0–8.7 × 1.6–2.3 | [13] |
| M. chrysanthemoides | Cylindrical to ellipsoidal | 5–12 × 3.0–4.5 | Ellipsoid or allantoid | 4.5–7 × 2–3 | [13] |
| M. chuxiongense | Solitary, clavate | 27–74 × 2–3 | Shuttle or sickle | 4–12 × 2–5 | [39] |
| M. citrinidiscum | Denticulate, cylindrical | 11–29 × 1.5–2 | clavate to obovoid | 7–31 × 2–3 | [13] |
| M. colombiense | Ampulliform, polyblastic | 5–11.5 × 2.5–3.5 | Fusiform, allantoid | 5–8 × 1.5–2.5 | [13] |
| M. dawsoniorum | Cylindrical to irregular | 20–30 × 1–2 | Flexuous to falcate | 25–75 × 1–2 | [13] |
| M. fisheri | Denticulate, cylindrical | 19–60 × 1.5–2 | Obovoid, subpyriform | 7–12 × 3–4 | [13] |
| M. hainanense | Ampulliform and lageniform | 4.8–8.2 × 2.0–2.5 | Spindle to rod shaped | 7.0–16.1 × 2.5–4.7 | [21] |
| M. indocalami | Cylindrical, straight or bent | 11.0–28.3 × 1.5–2.9 | Clavate to obovoid | 13.0–15.5 × 3.5–5.5 | [17] |
| M. lycopodinum | Ampulliform to lageniform | 4–12 × 2.5–3.5 | Fusiform or with one side | 8–15 × 2.5–3.5 | [13] |
| M. maculosum | Denticulate, raduliform | 7–39 × 1–3 | Fusiform, straight or curved | 6–15 × 2–4 | [18] |
| M. miscanthi | Smooth and cylindrical | 9.7–14.5 × 3.6–4.1 | Spindle to rod shaped | 7.0–16.1 × 2.5–4.7 | [21] |
| M. musae | |||||
| M. nannuoshanense | Cylindrical and smooth | 19.0–27.0 × 2.0–3.0 | Oblong to ellipsoid | 7.0–12.0 × 2.5–4.0 | This study |
| M. neoqueenslandicum | Lageniform to subcylindrical | 4.5–10 × 2–3.5 | Lunate, allantoid | 4–9 × 1.5–3 | [13] |
| M. nivale | Doliiform to obpyriform | 6–15 × 2.2–4 | Falcate, apex pointed | 5–36 × 2–4.5 | [13] |
| M. paspali | Lageniform to cylindrical | 6.5–15.5 × 2.5–4 | Falcate, apex pointed | 7–20.5 × 2.5–4.5 | [13] |
| M. phyllosaprophyticum | Cylindrical and smooth | 16.0–32.0 × 2.0–2.5 | Oblong to ellipsoid | 7.5–15.0 × 2.0–3.5 | This study |
| M. phragmitis | Ampulliform to lageniform | 5–12(–30) × 2.5–3 | Ellipsoid-fusiform | 10–16 × 2–3.5 | [13] |
| M. poae | Cylindrical or subcylindrical | 1.5–6.5 ×1–2 | Fusiform, ovoid, pyriform | 3.5–8.5 ×2–3 | [13] |
| M. ratticaudae | – | – | Fusoid, falcat | 7-11 × 1.5-2.5 | [13] |
| M. rhopalostylidis | Ampulliform | 4–10 × 3–3.5 | Fusoid, curved | 16–20(–23) × 2.5–3 | [15] |
| M. seminicola | Ampulliform to lageniform | 7–9.5 × 3–4 | Cylindrical to fusiform | 19–54 × 3–4.5 | [13] |
| M. sinense | Smooth and cylindrical | 16.3–22.4 × 4.1–5.7 | Spindle shaped or cylindrical | 11.5–19.34 × 2.8–5.4 | [21] |
| M. sorghi | Ampulliform to obclavate | 5–13 × 3–4 | Filiform, obclavate | 20–90 × 1.5–4.5 | [13] |
| M. tainanense | Cylindrical or ampulliform | 3–10 × 1–3 | Lunate | 10–15 × 2–3 | [13] |
| M. trichocladiopsis | Cylindrical to clavate | 4–37 × 2–3 | Oblong, fusiform to obovoid | 6–18 × 2–3.5 | [13] |
| M. yunnanense | Ampulliform, lageniform | 6.5–10.0 × 2.5–3.4 | Ellipsoid and cylindrical | 6.8–10.0 × 2.4–3.5 | [17] |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sydow, H. Mycotheca germanic. Fasc. XLII–XLV (No. 2051–2250). Annls Mycol. 1924, 22, 257–268. [Google Scholar]
- Liang, J.M.; Li, G.S.; Zhao, M.Q.; Cai, L. A new leaf blight disease of turfgrasses caused by Microdochium poae, sp. nov. Mycologia 2019, 111, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Von Arx, J.A. Plant pathogenic fungi. Nova Hedwig. 1987, 87, 1–288. [Google Scholar]
- Hiruma, K.; Kobae, Y.; Toju, H. Beneficial associations between Brassicaceae plants and fungal endophytes under nutrientlimiting conditions: Evolutionary origins and host–symbiont molecular mechanisms. Curr. Opin. Plant Biol. 2018, 44, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Mandyam, K.G.; Roe, J.; Jumpponen, A. Arabidopsis thaliana model system reveals a continuum of responses to root endophyte colonization. Fungal Biol. 2013, 117, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Glynn, N.C.; Hare, M.C.; Parry, D.W.; Edwards, S.G. Phylogenetic analysis of EF-1 alpha gene sequences from isolates of Microdochium nivale leads to elevation of varieties Majus and Nivale to species status. Fungal Biol. 2005, 109, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Jewell, L.E.; Hsiang, T. Multigene differences between Microdochium nivale and Microdochium majus. Botany 2013, 91, 99–106. [Google Scholar] [CrossRef]
- Hernández-Restrepo, M.; Groenewald, J.Z.; Crous, P.W. Taxonomic and phylogenetic re-evaluation of Microdochium, Monographella and Idriella. Pers. Mol. Phylogeny Evol. Fungi. 2016, 36, 57–82. [Google Scholar] [CrossRef]
- Parkinson, V.O.; Sivanesan, A.; Booth, C. The perfect state of the rice leaf-scald fungus and the taxonomy of both the perfect and imperfect states. Trans. Br. Mycol. Soc. 1981, 76, 59–69. [Google Scholar] [CrossRef]
- Samuels, G.J.; Hallett, I.C. Microdochium stoveri and Monographella stoveri, new combinations for Fusarium stoveri and Micronectriella stoveri. Trans. Br. Mycol. Soc. 1983, 81, 473–483. [Google Scholar] [CrossRef]
- Von Arx, J.A. Notes on Monographella and Microdochium. Trans. Br. Mycol. Soc. 1984, 83, 373–374. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Phylogenetic relationships of five genera of Xylariales and Rosasphaeria gen. nov. (Hypocreales). Fungal Divers. 2012, 52, 75–98. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable Mycobiota from karst caves in China, with descriptions of 20 new species. Pers. Mol. Phylogeny Evol. Fungi. 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumacher, R.K.; Wingfield, M.J.; Akulov, A.; Denman, S.; Roux, J.; Braun, U.; Burgess, T.I.; Carnegie, A.J.; Váczy, K.Z. New and interesting fungi 1. Fuse 2018, 1, 169–215. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W. New and interesting fungi 2. Fuse 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F. Genera of phytopathogenic fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef]
- Huang, S.T.; Xia, J.W.; Zhang, X.G.; Sun, W.X.; Li, Z. Two new species of Microdochium from Indocalamus longiauritus in south-western China. MycoKeys 2020, 72, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet description sheets: 1182–1283. Pers. Mol. Phylogeny Evol. Fungi. 2021, 46, 313–528. [Google Scholar] [CrossRef]
- Kwasna, H.; Bateman, G.L. Microdochium Triticicola sp. nov. from roots of Triticum aestivum in the United Kingdom. Mycologia 2007, 99, 765–776. [Google Scholar] [CrossRef]
- Zhang, W.; Nan, Z.B.; Tian, P.; Hu, M.J.; Gao, Z.Y.; Li, M.; Liu, G.D. Microdochium paspali, a new species causing seashore paspalum disease in southern China. Mycologia 2015, 107, 80–89. [Google Scholar] [CrossRef]
- Liu, S.B.; Liu, X.Y.; Zhang, Z.X.; Xia, J.W.; Zhang, X.G.; Meng, Z. Three new species of Microdochium (Sordariomycetes, Amphisphaeriales) on Miscanthus sinensis and Phragmites australis from Hainan, China. J. Fungi 2022, 8, 577. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, L.S.; Kang, J.C.; Maharachchikumbura, S.S.N. Microdochium sichuanense sp. nov. (Microdochiaceae, Xylariales), from a Poaceae host in Sichuan, China. Phytotaxa 2023, 600, 206–216. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Ma, C.Y.; Xue, H.; Piao, C.G.; Li, Y. A new Arthrinium-like genus of Amphisphaeriales in China. MycoKeys 2022, 92, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.X.; Liu, X.Y.; Tao, M.F.; Liu, X.Y.; Xia, J.W.; Zhang, X.G.; Meng, Z. Taxonomy, Phylogeny, Divergence Time Estimation, and Biogeography of the Family Pseudoplagiostomataceae (Ascomycota, Diaporthales). J. Fungi 2023, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, F.J.R.M.; Lee, S.H.; Taylor, L.; Shawe-Taylor, J. Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModelTest v. 2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment. Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: San Diego, CA, USA, 2012; p. 8. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Zou, W.; Wang, Y.; Huang, O.U.; Yu, H. Morphology and phylogeny of Microdochium chuxiongense sp. nov., a fungus from Southwest China. Phytotaxa 2022, 555, 147–158. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Taylor, J.W.; Wingfield, M.J.; Abaci, O.; Aime, C. The Amsterdam declaration on fungal nomenclature. IMA Fungus 2011, 2, 105–111. [Google Scholar] [CrossRef]
- Abdelhalim, M.; Brurberg, M.B.; Hofgaard, I.S.; Rognli, O.A.; Tronsmo, A.M. Pathogenicity, host specificity and genetic diversity in Norwegian isolates of Microdochium nivale and Microdochium majus. Eur. J. Plant Pathol. 2020, 156, 885–895. [Google Scholar] [CrossRef]
- Gao, Y.; Ren, G.; Wanasinghe, D.N.; Xu, J.; Gomes De Farias, A.R.; Gui, H. Two New Species and a New Record of Microdochium from Grasses in Yunnan Province, South-West China. J. Fungi 2022, 8, 1297. [Google Scholar] [CrossRef]
- Monkai, J.; Phookamsak, R.; Tennakoon, D.S.; Bhat, D.J.; Xu, S.; Li, Q.; Lumyong, S. Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand. Diversity 2022, 14, 918. [Google Scholar] [CrossRef]
- Shadmani, L.; Jamali, S.; Fatemi, A. Biocontrol activity of endophytic fungus of barley, Microdochium bolleyi, against Gaeumannomyces graminis var. tritici. Mycol. Iran. 2018, 5, 7–14. [Google Scholar] [CrossRef]
| Loci | PCR Primers | Sequence (5′–3′) | PCR Cycles | References |
|---|---|---|---|---|
| LSU | LR0R | GTA CCC GCT GAA CTT AAG C | (95 °C: 30 s, 51 °C: 60 s, 72 °C: 1 min) × 35 cycles | [27] |
| LR5 | TCC TGA GGG AAA CTT CG | |||
| ITS | ITS5 | GGA AGT AAA AGT CGT AAC AAG G | (95 °C: 30 s, 55 °C: 30 s, 72 °C: 30 s) × 35 cycles | [28] |
| ITS4 | TCC TCC GCT TAT TGA TAT GC | |||
| RPB2 | RPB2-5F | GAY GAY MGW GAT CAY TTY GG | (95 °C: 30 s, 56 °C: 30 s, 72 °C: 1 min) × 35 cycles | [29,30] |
| RPB2-7CR | CCC ATW GCY TGC TTM CCC AT | |||
| TUB2 | Btub526_F | CGA GCG YAT GAG YGT YTA CTT | (95 °C: 30 s, 56 °C: 30 s, 72 °C: 45 s) × 35 cycles | [7] |
| Btub1332_R | TCA TGT TCT TGG GGT CGA A |
| Species | Strain No. | Region | GenBank Accession No. | |||
|---|---|---|---|---|---|---|
| ITS | LSU | RPB2 | TUB2 | |||
| Idriela lunata | CBS 204.56 T | USA | KP859044 | KP858981 | – | – |
| CBS 177.57 | USA | KP859043 | KP858980 | – | – | |
| Microdochium albescens | CBS 243.83 | Ivory Coast | KP858994 | KP858930 | KP859103 | KP859057 |
| CBS 291.79 | Ivory Coast | KP858996 | KP858932 | KP859105 | KP859059 | |
| M.bambusae | SAUCC 1862-1T | China | OR702567 | OR702576 | OR715791 | OR715785 |
| SAUCC 1866-1 | China | OR702568 | OR702577 | OR715792 | OR715786 | |
| M. bolleyi | CBS 540.92 | Syria | KP859010 | KP858946 | KP859119 | KP859073 |
| M. chrysanthemoides | CGMCC 3.17929 T | China | KU746690 | KU746736 | – | – |
| M. chuxiongense | YFCC 8794 T | China | OK586161 | OK586160 | OK584019 | OK556901 |
| M. citrinidiscum | CBS 109067 T | Peru | KP859003 | KP858939 | KP859112 | KP859066 |
| M. colombiense | CBS 624.94 | Colombia | KP858999 | KP858935 | KP859108 | KP859062 |
| M. dawsoniorum | BRIP 65649 T | Australia | MK966337 | – | – | – |
| M. fisheri | CBS 242.90 T | UK | KP859015 | KP858951 | KP859124 | KP859078 |
| M. graminearum | CGMCC 3.23525 T | China | OP103966 | OP104016 | OP236027 | – |
| CGMCC 3.23524 | China | OP103965 | OP104015 | OP236026 | – | |
| M. hainanense | SAUCC 210782 | China | OM956296 | OM959324 | OM981154 | OM981147 |
| SAUCC 210781 T | China | OM956295 | OM959323 | OM981153 | OM981146 | |
| M. indocalami | SAUCC 1016 T | China | MT199884 | MT199878 | MT510550 | MT435653 |
| M. lycopodinum | CBS 146.68 | The Netherlands | KP858993 | KP858929 | KP859102 | KP859056 |
| CBS 122885 T | Germany | KP859016 | KP858952 | KP859125 | KP859080 | |
| M. maculosum | COAD 3358 T | Brazil | Ok966954 | Ok966953 | – | – |
| M. majus | CBS 741.79 | Germany | KP859001 | KP858937 | KP859110 | KP859064 |
| M. miscanthi | SAUCC 211092 T | China | OM956214 | OM957532 | OM981148 | OM981141 |
| SAUCC 211093 | China | OM956215 | OM957533 | OM981149 | OM981142 | |
| M. musae | CPC 32809 | Malaysia | MH107894 | MH107941 | – | – |
| CBS 143500 T | Malaysia | MH107895 | MH107942 | MH108003 | – | |
| M. nannuoshanense | SAUCC 2450-1 T | China | OR702569 | OR702578 | OR715793 | OR715787 |
| SAUCC 2450-3 | China | OR702570 | OR702579 | OR715794 | OR715788 | |
| M. neoqueenslandicum | CBS 445.95 | The Netherlands | KP858997 | KP858933 | KP859106 | KP859060 |
| CBS 108926 T | The Netherlands | KP859002 | KP858938 | KP859111 | KP859065 | |
| M. nivale | CBS 116205 T | UK | KP859008 | KP858944 | KP859117 | KP859071 |
| M. nivale var. majus | CBS 177.29 | The Netherlands | MH855031 | MH866500 | – | – |
| M. nivale var. nivales | CBS 288.50 | The Netherlands | – | MH868135 | – | – |
| M. novae-zelandiae | CPC 29376 T | The Netherlands | LT990655 | – | LT990641 | LT990608 |
| CPC 29693 | The Netherlands | LT990656 | n/a | LT990642 | LT990609 | |
| M. paspali | HK-ML-1371 | China | KJ569509 | – | – | KJ569514 |
| CBS 138620 T | China | KJ569513 | – | – | KJ569518 | |
| M. phyllosaprophyticum | SAUCC 3583-1 T | China | OR702571 | OR702580 | OR715795 | OR715789 |
| SAUCC 3583-6 | China | OR702572 | OR702581 | OR715796 | OR715790 | |
| M. phragmitis | CBS 285.71 T | Poland | KP859013 | KP858949 | KP859122 | KP859077 |
| CBS 423.78 | Poland | KP859012 | KP858948 | KP859121 | KP859076 | |
| M. poae | CGMCC3.19170 T | China | MH740898 | – | MH740906 | MH740914 |
| LC 12115 | China | MH740901 | – | MH740909 | MH740917 | |
| LC 12116 | China | MH740902 | – | MH740910 | MH740918 | |
| M. ratticaudae | BRIP 68298 T | Australia | MW481661 | MW481666 | MW626890 | – |
| M. rhopalostylidis | CBS 145125 T | The Netherlands | MK442592 | MK442532 | MK442667 | – |
| M. seminicola | CBS 139951 T | Switzerland | KP859038 | KP858974 | KP859147 | KP859101 |
| CPC 26001 | Canada | KP859025 | KP858961 | KP859134 | KP859088 | |
| DAOM 250161 | Canada | KP859034 | KP858970 | KP859143 | KP859097 | |
| M. shilinense | CGMCC 3.23531 T | China | OP103972 | OP104022 | – | OP242834 |
| M. sinense | SAUCC 211097 T | China | OM956289 | OM959225 | OM981151 | OM981144 |
| SAUCC 211098 | China | OM956290 | OM959226 | OM981152 | OM981145 | |
| M. sorghi | CBS 691.96 | Cuba | KP859000 | KP858936 | KP859109 | KP859063 |
| M. tainanense | CBS 269.76 T | The Netherlands | KP859009 | KP858945 | KP859118 | KP859072 |
| CBS 270.76 | The Netherlands | KP858995 | KP858931 | KP859104 | KP859058 | |
| M. trichocladiopsis | CBS 623.77 T | The Netherlands | KP858998 | KP858934 | KP859107 | KP859061 |
| M. yunnanense | SAUCC 1011 T | China | MT199881 | MT199875 | MT510547 | MT435650 |
| SAUCC 1012 | China | MT199882 | MT199876 | MT510548 | MT435651 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, Z.; Li, D.; Xia, J.; Li, Z. Three New Species of Microdochium (Microdochiaceae, Xylariales) on Bambusaceae sp. and Saprophytic Leaves from Hainan and Yunnan, China. J. Fungi 2023, 9, 1176. https://doi.org/10.3390/jof9121176
Zhang J, Zhang Z, Li D, Xia J, Li Z. Three New Species of Microdochium (Microdochiaceae, Xylariales) on Bambusaceae sp. and Saprophytic Leaves from Hainan and Yunnan, China. Journal of Fungi. 2023; 9(12):1176. https://doi.org/10.3390/jof9121176
Chicago/Turabian StyleZhang, Jie, Zhaoxue Zhang, Duhua Li, Jiwen Xia, and Zhuang Li. 2023. "Three New Species of Microdochium (Microdochiaceae, Xylariales) on Bambusaceae sp. and Saprophytic Leaves from Hainan and Yunnan, China" Journal of Fungi 9, no. 12: 1176. https://doi.org/10.3390/jof9121176