
Novelties in Microthyriaceae (Microthyriales): Two New Asexual Genera with Three New Species from Freshwater Habitats in Guizhou Province, China
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Examination of Specimens
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
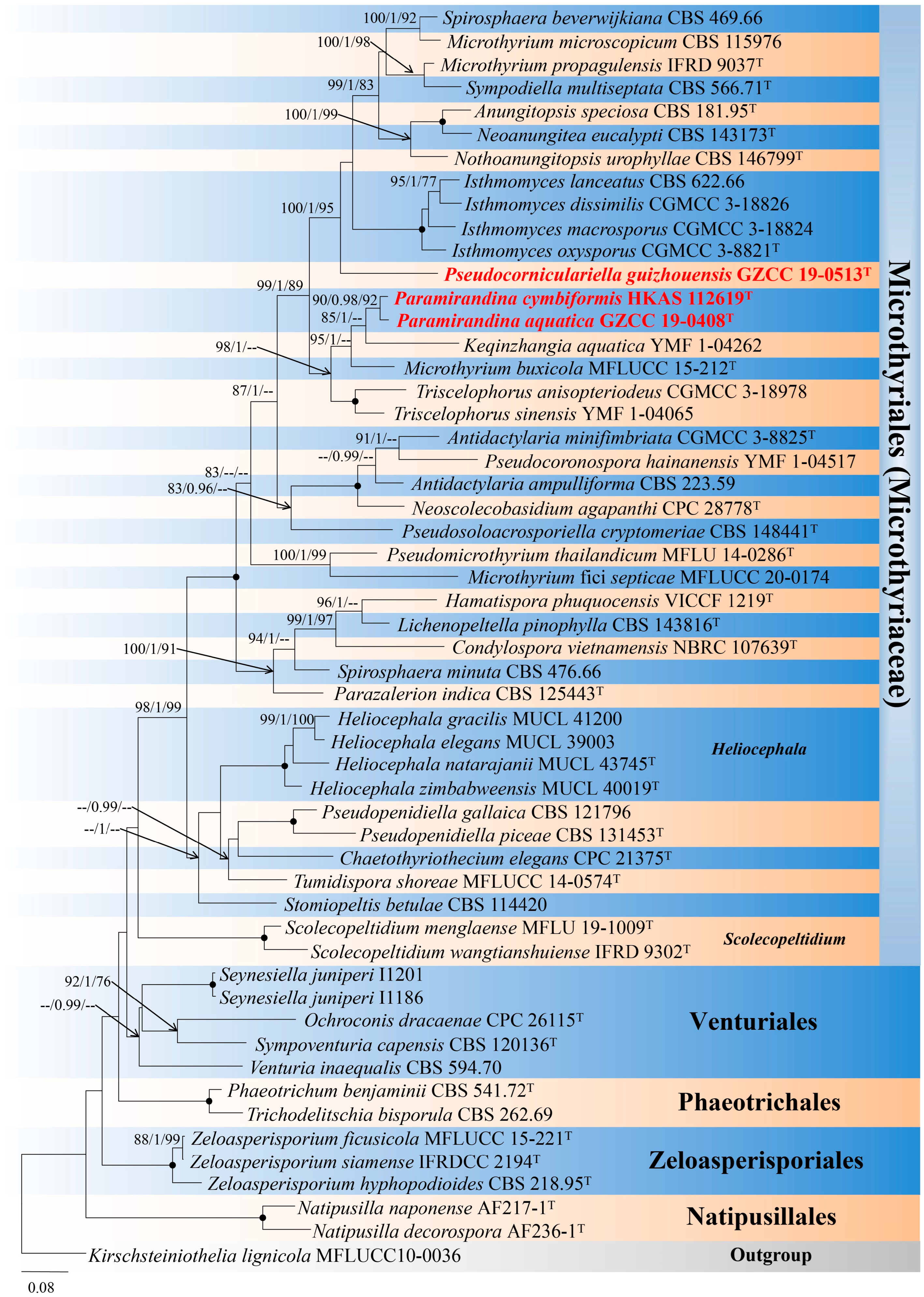
3. Phylogenetic Results
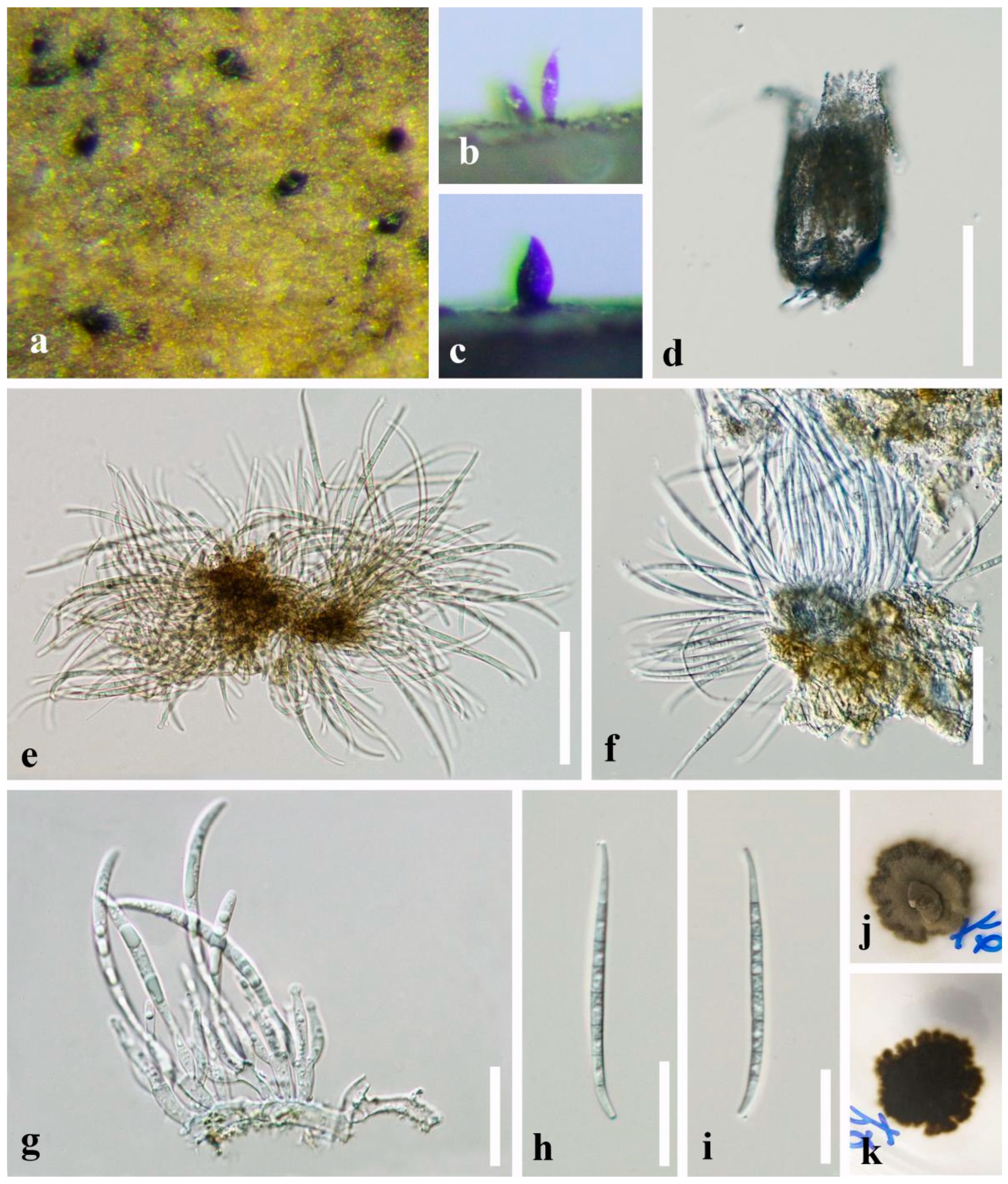
4. Taxonomy
4.1. Paramirandina L.L. Liu & Z.Y. Liu, gen. nov.
4.2. Pseudocorniculariella L.L. Liu & Z.Y. Liu, gen. nov.
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saccardo, P.A. Sylloge Pyrenomycetum. Sylloge Fungorum 1883, 2, 1–813. [Google Scholar]
- Hongsanan, S.; Hyde, K.D. Phylogenetic placement of Micropeltidaceae. Mycosphere 2017, 8, 1930–1942. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Wu, H.X.; Hyde, K.D.; Chen, H. Studies on Microthyriaceae: Placement of Actinomyxa, Asteritea, Cirsosina, Polystomellina and Stegothyrium. Cryptogam Mycol. 2011, 32, 3–12. [Google Scholar] [CrossRef]
- Wu, H.X.; Li, Y.M.; Chen, H.; Hyde, K.D. Studies on Microthyriaceae: Some excluded genera. Mycotaxon 2010, 113, 147–156. [Google Scholar] [CrossRef]
- Wu, H.X.; Tian, Q.; Li, W.J.; Hyde, K.D. A reappraisal of Microthyriaceae. Phytotaxa 2014, 176, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.X.; Schoch, C.L.; Boonmee, S.; Bahkali, A.H.; Chomnunti, P.; Hyde, K.D. A reappraisal of Microthyriaceae. Fungal Divers. 2011, 51, 189–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; McKenzie, E.H.C.; Wang, Y. Notes for genera update Ascomycota: 6822–6917. Mycosphere 2018, 9, 1222–1234. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.N.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Yen, L.T.H.; Yamaguchi, K.; Tsurumi, Y.; Hop, D.V.; Ando, K. Hamatispora, a new genus of aquatic fungi in Microthyriales isolated from fallen leaves in Vietnam. Mycoscience 2018, 59, 467–472. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.S.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Hobraken, J.; et al. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef] [PubMed]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Lucking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.G. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Qiao, M.; Zheng, H.; Guo, J.S.; Castañeda-Ruiz, R.F.; Xu, J.P.; Peng, J.; Zhang, K.Q.; Yu, Z.F. Two new asexual genera and six new asexual species in the family Microthyriaceae (Dothideomycetes, Ascomycota) from China. MycoKeys 2021, 85, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Qiao, M.; Guo, J.; Castañeda-Ruiz, R.F.; Peng, J.; Yu, Z.F. Keqinzhangia aquatica gen. et sp. nov. and Pseudocoronospora hainanense gen. et sp. nov. isolated from freshwater in southern China. Antonie van Leeuwenhoek 2022, 115, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Kandawatte, T.C.; Clericuzio, M.; Dayarathne, M.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Al-Sadi, A.M.; Liu, Z.Y. Pseudostanjehughesia aquitropica gen. et sp. nov. and Sporidesmium sensu lato species from freshwater habitats. Mycol. Prog. 2018, 17, 591–616. [Google Scholar] [CrossRef]
- Liu, L.L.; Liu, N.G.; Yang, J.; Chen, Y.Y.; Gui, X.X.; Liu, Z.Y. Sporidesmium guizhouense sp. nov. (Sordariomycetes incertae sedis), a new species from a freshwater habitat in Guizhou Province, China. Phytotaxa 2019, 422, 144–156. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.L.; Liu, N.G.; Yang, J.; Chen, Y.Y.; Liu, Z.Y. Cucurbitinus gen. nov. (Halosphaeriaceae, Microascales), a new genus to accommodate Cucurbitinus constrictus comb. nov. and Cucurbitinus ibericus comb. nov. Phytotaxa 2020, 455, 119–136. [Google Scholar] [CrossRef]
- Yang, J.; Liu, L.L.; Jones, E.B.G.; Li, W.L.; .Hyde, K.D.; Liu, Z.Y. Morphological Variety in Distoseptispora and Introduction of Six Novel Species. J. Fungi. 2021, 7, 945. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.L.; Hyde, K.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Su, H.Y. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Indunil Chinthani, S.; Achala, R.R.; Diana, S.M.; Mark, S.C.; Eleni, G.; Hyang Burm, L.; Vedprakash, G.H.; Dhandevi, P.; Lakmali, S.D.; Subodini, N.W.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, D.J.; Buyck, B.; Cai, L.; Dai, Y.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 12 January 2023).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multi-gene phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phyl. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA Polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J. MrModeltest, version 2.3—Program for Selecting DNA Substitution Models Using PAUP*; Evolutionary Biology Centre: Uppsala, Sweden, 2008.
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larget, B.; Simon, D.L. Markov Chain Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*, Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4; Sinauer Associates: Sunderland, UK, 2003.
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Rao, V.G.; Reddy, K.A.; de Hoog, G.S. Heliocephala, a new genus of dematiaceous Hyphomycetes. Persoonia 1984, 12, 239–242. [Google Scholar]
- Goos, R.D.; DeHoog, G.S. Taxonomy of the Dactylaria Complex, IV-VI. Mycologia 1986, 78, 978. [Google Scholar] [CrossRef]
- Réblová, M.; Seifert, K.A.; Fournier, J.; Stepánek, V. Phylogenetic classification of Pleurothecium and Pleurotheciella gen. nov. and its dactylaria-like anamorph (Sordariomycetes) based on nuclear ribosomal and protein-coding genes. Mycologia 2012, 104, 1299–1314. [Google Scholar] [CrossRef]
- Ma, Y.R.; Xia, J.W.; Zhang, X.G.; Castaeda-Ruiz, R.F. New species of Phaeomonilia and Mirandina from dead branches in China. Mycotaxon 2015, 130, 775–781. [Google Scholar] [CrossRef]
- Li, W.J.; McKenzie, E.H.C.; Liu, J.K.; Bhat, D.J.; Dai, D.Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.N.; Luo, Z.L.; Shang, Q.J.; et al. Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar] [CrossRef]
- Shearer, C.A. The freshwater ascomycetes. Nova Hedwig 1993, 56, 1–33. [Google Scholar]
- Hyde, K.D.; Fryar, S.; Tian, Q.; Bahkali, A.H.; Xu, J.C. Lignicolous freshwater fungi along a north-south latitudinal gradient in the Asian/Australian region; can we predict the impact of global warming on biodiversity and function? Fungal Ecol. 2016, 19, 190–200. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Doilom, M.; Phookamsak, R.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Yang, J.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Liu, Z.Y. New species in Dictyosporium, new combinations in Dictyocheirospora and an updated backbone tree for Dictyosporiaceae. MycoKeys 2018, 36, 83–105. [Google Scholar] [CrossRef]
- Yang, J.; Liu, N.G.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Liu, Z.Y. Phylogenetic placement of Cryptophiale, Cryptophialoidea, Nawawia, Neonawawia gen. nov. and Phialosporostilbe. Mycosphere 2018, 9, 1132–1150. [Google Scholar] [CrossRef]
- Yu, X.D.; Dong, W.; Bhat, D.J.; Boonmee, S.; Zhang, D.I.; Zhang, H. Cylindroconidiis aquaticus gen. et sp. nov., a new lineage of aquatic hyphomycetes in Sclerococcaceae (Eurotiomycetes). Phytotaxa 2018, 372, 79–87. [Google Scholar] [CrossRef]
- Wang, G.N.; Yu, X.D.; Dong, W.; Bhat, D.J.; Boonmee, S.; Zhang, D.; Zhang, H. Freshwater hyphomycetes in Eurotiomycetes: A new species of Minimelanolocus and a new collection of Thysanorea papuana (Herpotrichiellaceae). Mycol Prog. 2019, 18, 511–522. [Google Scholar] [CrossRef]
- Calabon, M.S.; Hyde, K.D.; Jones, E.B.G.; Luo, Z.L.; Dong, W.; Hurdeal, V.G.; Gentekaki, E.; Rossi, W.; Leonardi, M.; Thiyagaraja, V.; et al. Freshwater fungal numbers. Fungal Divers. 2022, 003, 3–235. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Crous, P.W.; Kirk, P.M.; Hawksworth, D.L.; Boonmee, S.; Braun, U.; Dai, D.Q.; D’souza, M.; Diederich, P.; Dissanayake, A.; et al. Naming and outline of Dothideomycetes-2014 including proposals for the protection or suppression of generic names. Fungal Divers. 2014, 69, 1–55. [Google Scholar] [CrossRef]
- Heredia-Abarca, G.; Castañeda-Ruiz, R.F.; Arias-Mota, R.M.; Becerra-Hernandez, C.I.; Gómez, S.; Bogale, M.; Untereiner, W.A. A new species of Heliocephala from México with an assessment of the systematic positions of the anamorph genera Heliocephala and Holubovaniella. Mycologia 2011, 103, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, I.I.; Garcia, D.; Guarro, J.; Gene, J. Heliocephala variabilis and Pseudopenidiella vietnamensis: Two new Hyphomycetous species in the Microthyriaceae (Dothideomycetes) from Vietnam. Microorganisms 2020, 8, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher/Strain Number | GenBank Accession Number | |
|---|---|---|---|
| LSU | ITS | ||
| Antidactylaria ampulliforma | CBS 223.59 | MH869386 | MH857845 |
| Antidactylaria minifimbriata | CGMCC 3-18825T | MK577808 | MK569506 |
| Anungitopsis speciosa | CBS 181.95T | EU035401 | EU035401 |
| Chaetothyriothecium elegans | CPC 21375T | KF268420 | - |
| Condylospora vietnamensis | NBRC 107639T | LC146725 | LC146723 |
| Hamatispora phuquocensis | VICCF 1219T | LC064073 | LC064074 |
| Heliocephala elegans | MUCL 39003 | HQ333478 | HQ333478 |
| Heliocephala gracilis | MUCL 41200 | HQ333479 | HQ333479 |
| Heliocephala natarajanii | MUCL 43745T | HQ333480 | HQ333480 |
| Heliocephala zimbabweensis | MUCL 40019T | HQ333481 | HQ333481 |
| Isthmomyces dissimilis | CGMCC 3 18826 | MK577811 | MF740794 |
| Isthmomyces lanceatus | CBS 622.66 | MH870563 | MH858897 |
| Isthmomyces macrosporus | CGMCC 3-18824 | MK577812 | MF740796 |
| Isthmomyces oxysporus | CGMCC 3-18821T | MK577810 | MF740793 |
| Keqinzhangia aquatica | YMF 1-04262 | MK577809 | MK569507 |
| Kirschsteiniothelia lignicola | MFLUCC 10-0036 | HQ441568 | HQ441567 |
| Lichenopeltella pinophylla | CBS 143816T | MG844152 | - |
| Microthyrium buxicola | MFLUCC 15-0212T | KT306551 | - |
| Microthyrium fici-septicae | MFLUCC 20-0174T | MW063252 | - |
| Microthyrium microscopicum | CBS 115976 | GU301846 | - |
| Microthyrium propagulensis | IFRD 9037T | KU948989 | - |
| Natipusilla decorospora | AF236-1T | HM196369 | - |
| Natipusilla naponense | AF217-1T | HM196371 | - |
| Neoanungitea eucalypti | CBS 143173T | MG386031 | MG386031 |
| Neoscolecobasidium agapanthi | CPC 28778T | NG_059748 | NR_152546 |
| Nothoanungitopsis urophyllae | CBS 146799T | MW883825 | MW883433 |
| Ochroconis dracaenae | CPC 26115T | KX228334 | KX228283 |
| Paramirandina aquatica | GZCC 19-0408T | OQ025201 | OQ025199 |
| Paramirandina cymbiformis | HKAS 112619T | OQ025202 | - |
| Parazalerion indica | CBS 125443T | MH874977 | MH863483 |
| Phaeotrichum benjaminii | CBS 541.72T | MH872266 | MH860561 |
| Pseudocorniculariella guizhouensis | GZCC 19-0513T | OQ025203 | OQ025200 |
| Pseudocoronospora hainanensis | YMF 1-04517 | MK577807 | MK569505 |
| Pseudomicrothyrium thailandicum | MFLU 14-0286T | MT741680 | - |
| Pseudopenidiella gallaica | CBS 121796 | LT984843 | LT984842 |
| Pseudopenidiella piceae | CBS 131453T | JX069852 | JX069868 |
| Pseudosoloacrosporiella cryptomeriae | CBS 148441T | NG_081320 | NR_175206 |
| Scolecopeltidium menglaense | MFLU 19-1009T | MW003710 | MW003724 |
| Scolecopeltidium wangtianshuiense | IFRD 9302T | NG_067860 | NR_166263 |
| Seynesiella juniperi | I1201 | MW405232 | MW405223 |
| Seynesiella juniperi | I1186 | MW405230 | MW405222 |
| Spirosphaera beverwijkiana | CBS 469.66 | HQ696657 | HQ696657 |
| Spirosphaera minuta | CBS 476.66 | HQ696659 | HQ696659 |
| Stomiopeltis betulae | CBS 114420 | GU214701 | GU214701 |
| Sympodiella multiseptata | CBS 566.71T | MH872028 | MH860264 |
| Sympoventuria capensis | CBS 120136T | KF156104 | DQ885906 |
| Trichodelitschia bisporula | CBS 262.69 | MH871039 | MH859305 |
| Triscelophorus anisopteriodeus | CGMCC 3-18978 | MK577818 | MK569511 |
| Triscelophorus sinensis | YMF 1-04065 | MK577820 | MK569513 |
| Tumidispora shoreae | MFLUCC 14-0574T | KT314074 | - |
| Venturia inaequalis | CBS 594.70 | GU301879 | KF156040 |
| Zeloasperisporium ficusicola | MFLUCC 15-0221T | KT387733 | - |
| Zeloasperisporium hyphopodioides | CBS 218.95T | EU035442 | EU035442 |
| Zeloasperisporium siamense | IFRDCC 2194T | JQ036228 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Yang, J.; Zhou, S.; Gu, X.; Gou, J.; Wei, Q.; Zhang, M.; Liu, Z. Novelties in Microthyriaceae (Microthyriales): Two New Asexual Genera with Three New Species from Freshwater Habitats in Guizhou Province, China. J. Fungi 2023, 9, 178. https://doi.org/10.3390/jof9020178
Liu L, Yang J, Zhou S, Gu X, Gou J, Wei Q, Zhang M, Liu Z. Novelties in Microthyriaceae (Microthyriales): Two New Asexual Genera with Three New Species from Freshwater Habitats in Guizhou Province, China. Journal of Fungi. 2023; 9(2):178. https://doi.org/10.3390/jof9020178
Chicago/Turabian StyleLiu, Lingling, Jing Yang, Si Zhou, Xiaofeng Gu, Jiulan Gou, Quanquan Wei, Meng Zhang, and Zuoyi Liu. 2023. "Novelties in Microthyriaceae (Microthyriales): Two New Asexual Genera with Three New Species from Freshwater Habitats in Guizhou Province, China" Journal of Fungi 9, no. 2: 178. https://doi.org/10.3390/jof9020178