
Plant-Associated Novel Didymellaceous Taxa in the South China Botanical Garden (Guangzhou, China)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling Sites, Specimens, and Isolates
2.2. Morphological Studies
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Analyses
2.5. PHI Analyses
3. Results
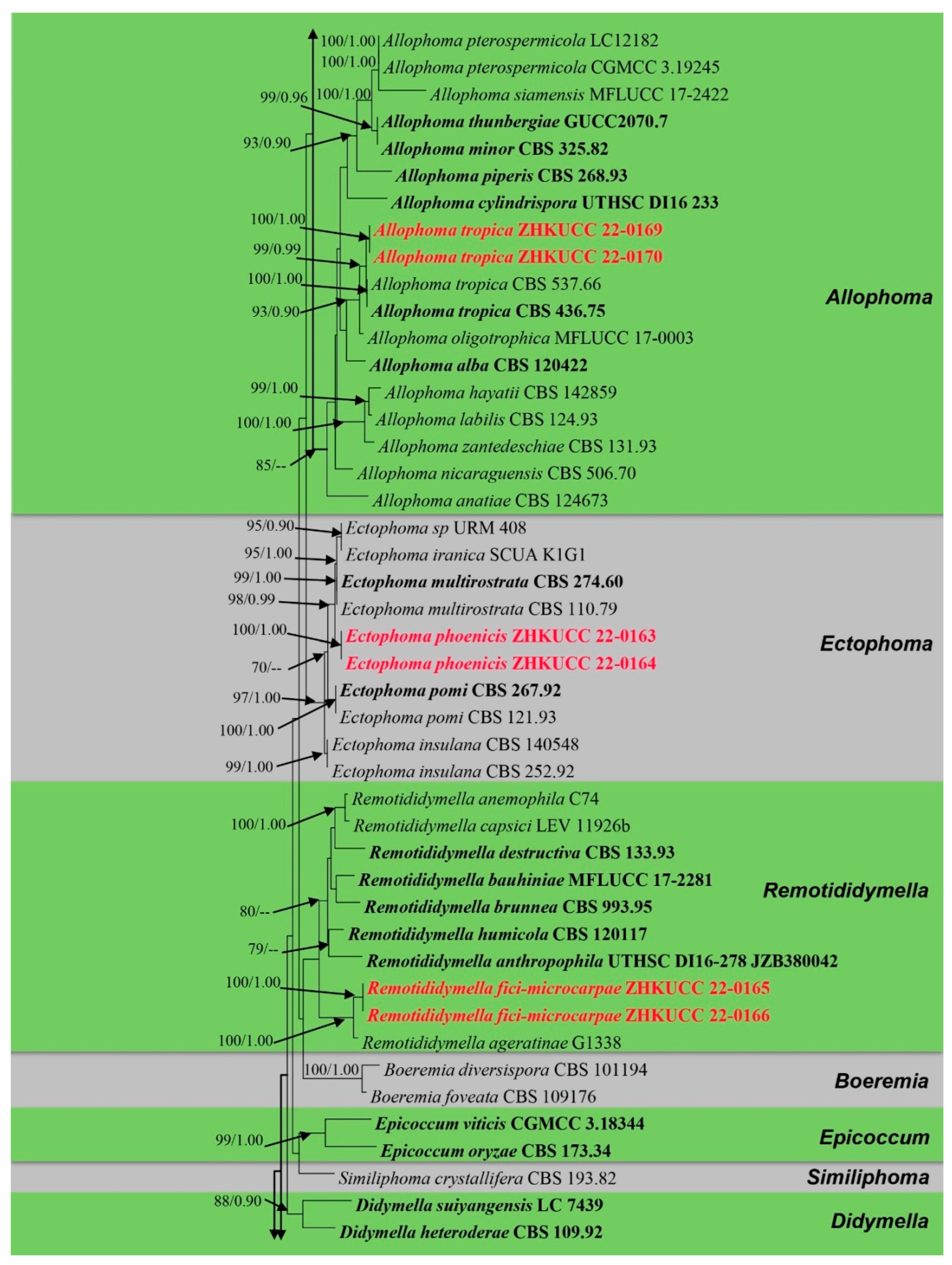
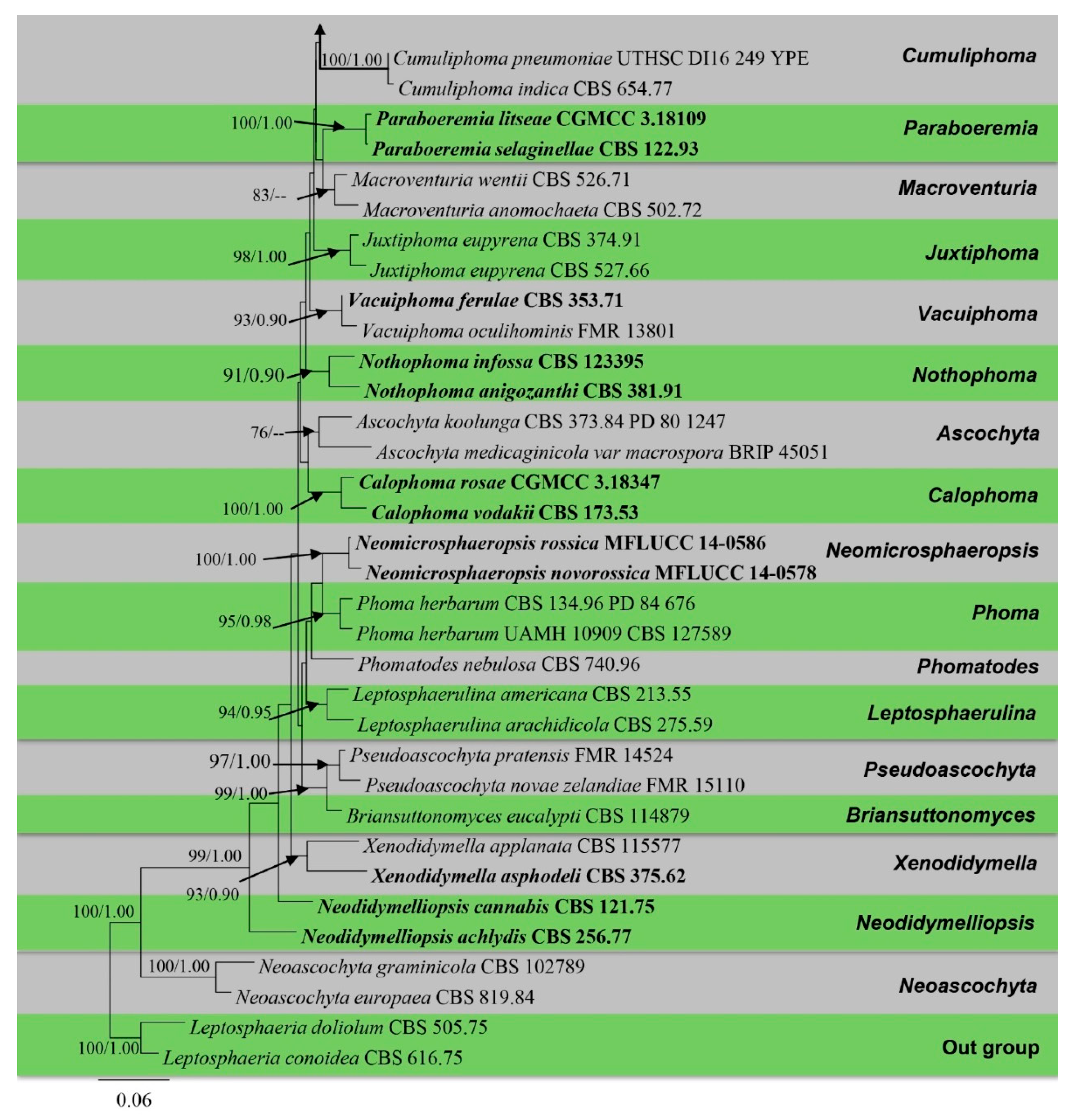
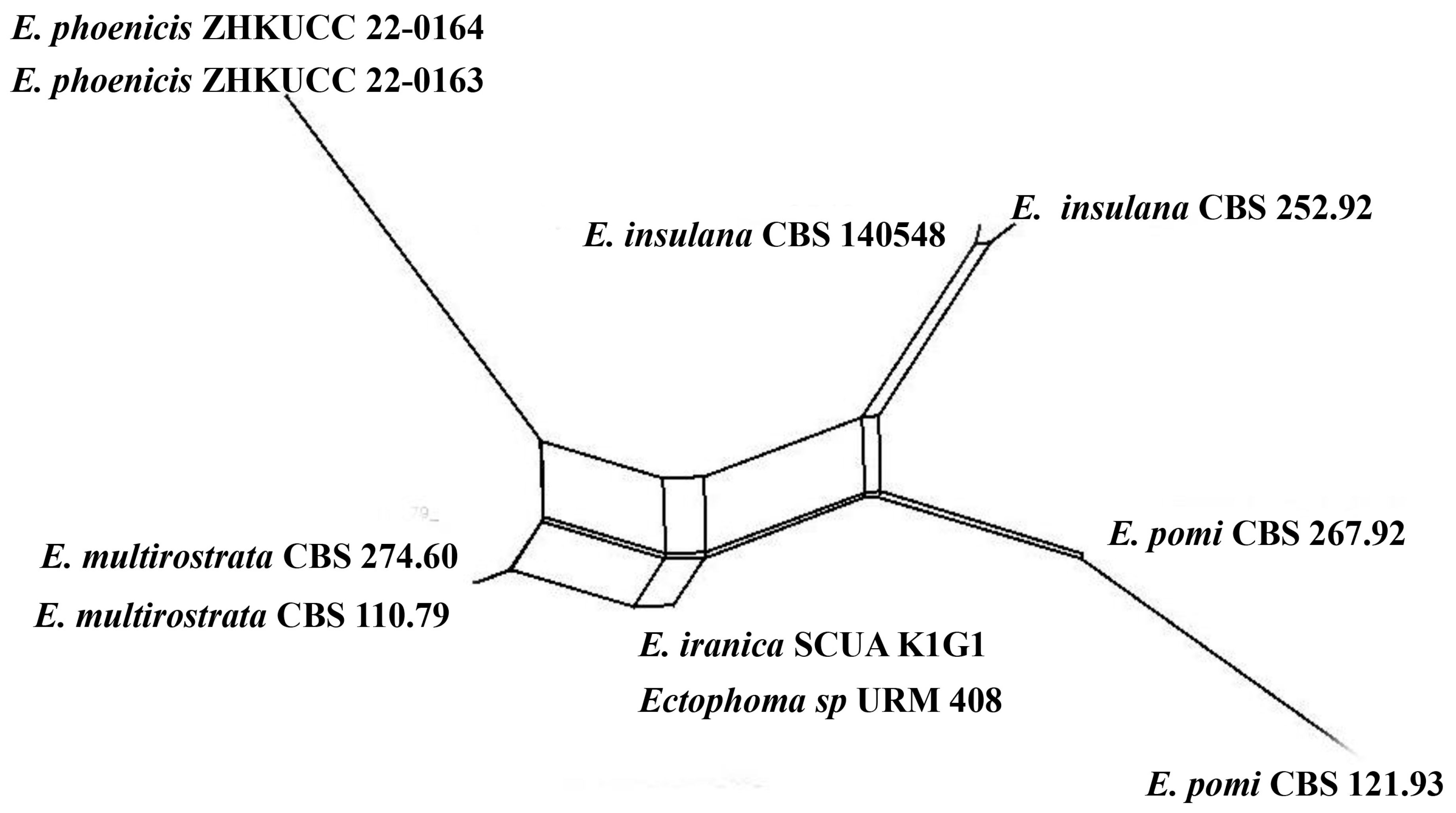
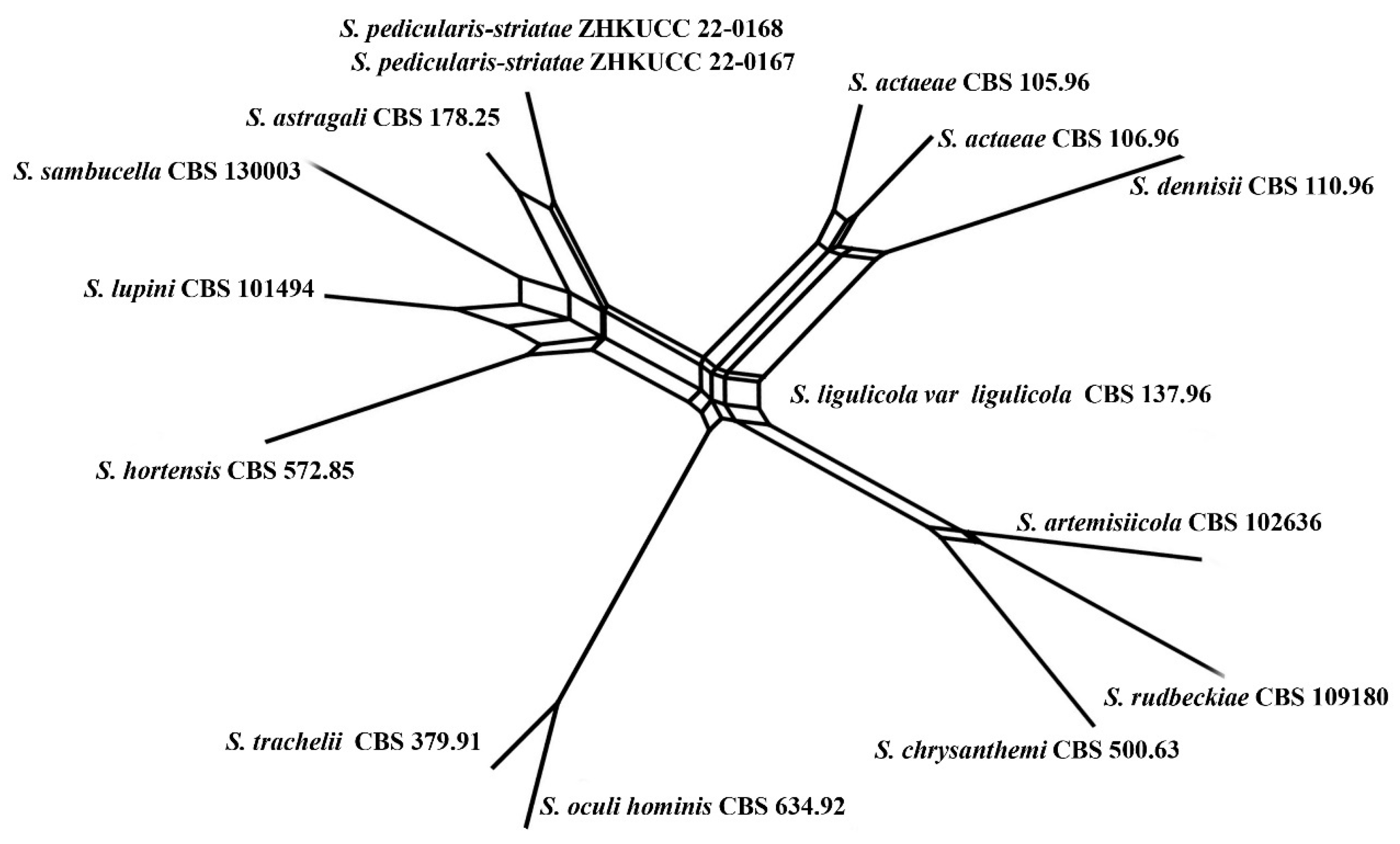
3.1. Phylogenetic Analyses
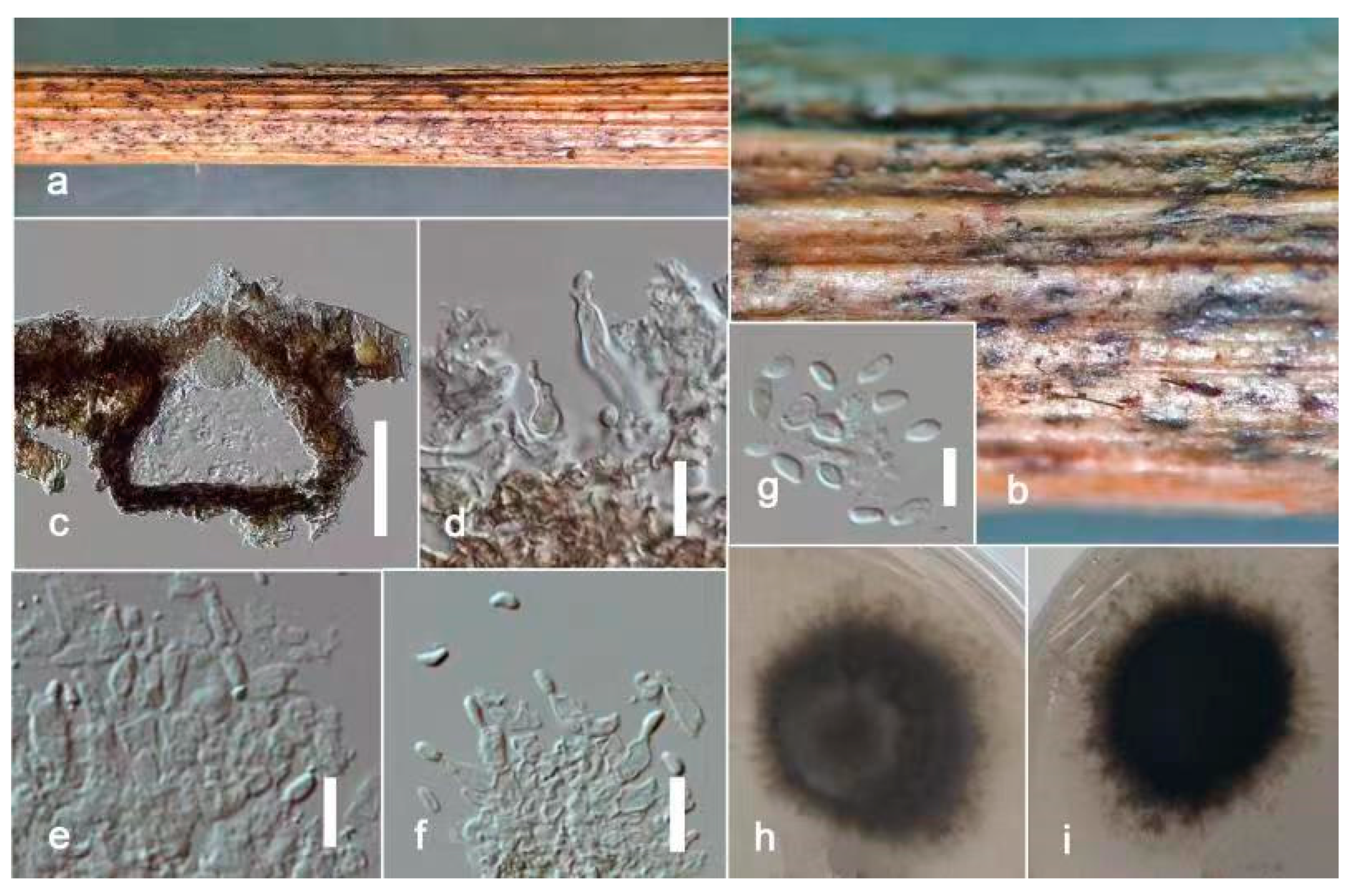
3.2. Taxonomy
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, H.W. The Chinese Botanical Gardens; Chinees Forestry Press: Beijing, China, 2018. [Google Scholar]
- Huang, H.W. The Chinese Botanical Gardens; EDP Sciences: Les Ulis, France, 2022; p. 437. [Google Scholar]
- Ren, H.; Jian, S.G.; Liu, H.X.; Zhang, Q.M.; Lu, H.F. Advances in the reintroduction of rare and endangered wild plant species. Sci. China Life Sci. 2014, 57, 603–609. [Google Scholar] [CrossRef]
- de Gruyter, J.; Aveskamp, M.M.; Woudenberg, J.H.C.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Molecular phylogeny of Phoma and allied anamorph genera towards a reclassification of the Phoma complex. Mycol. Res. 2009, 113, 508–519. [Google Scholar] [CrossRef]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Wanasinghe, D.N.; Papizadeh, M.; Goonasekara, I.D.; Camporesi, E.; Bhat, D.J.; McKenzie, E.H.; Phillips, A.J.; Diederich, P.; et al. Taxonomy and phylogeny of dematiaceous coelomycetes. Fungal Divers. 2016, 77, 1–316. [Google Scholar]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.; Bulgakov, T.; Bhat, D.J.; Camporesi, E.; Bahkali, A.H.; Eungwanichayapant, P.D.; Liu, Z.-Y.; Hyde, K.D. Microfungi on Tamarix. Fungal Divers. 2017, 82, 239–306. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Jeewon, R.; Peršoh, D.; Jones, E.B.G.; Camporesi, E.; Bulgakov, T.S.; Gafforov, Y.S.; Hyde, K.D. Taxonomic circumscription and phylogenetics of novel didymellaceous taxa with brown muriform spores. Stud. Fungi 2018, 3, 152–175. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.; Sarma, V.V.; Lücking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.G.; et al. Refined families of Dothideomycetes: Orders and families incertaesedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Hou, L.W.; Hernández-Restrepo, M.; Groenewald, J.Z.; Cai, L.; Crous, P.W. Citizen science project reveals high diversity in Didymellaceae (Pleosporales, Dothideomycetes). MycoKeys 2020, 65, 49–99. [Google Scholar] [CrossRef]
- Keirnan, E.C.; Tan, Y.P.; Laurence, M.H.; Mertin, A.A.; Liew, E.C.; Summerell, B.A.; Shivas, R.G. Cryptic diversity found in Didymellaceae from Australian native legumes. MycoKeys 2021, 78, 1–20. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of fungi and fungus-like taxa 2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Aveskamp, M.M.; de Gruyter, J.; Crous, P.W. Biology and recent developments in the systematics of Phoma, a complex genus of major quarantine significance. Fungal Divers. 2008, 31, 1–18. [Google Scholar]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Divakar, P.K.; Rajeshkumar, K.C.; Weerahewa, D.; Delgado, G.; Wang, Y.; Fu, L. Notes for genera update—Ascomycota: 6616–6821. Mycosphere 2018, 9, 115–140. [Google Scholar] [CrossRef]
- Babaahmadi, G.; Mehrabi-Koushki, M.; Hayati, J. Allophoma hayatii sp. nov.; an undescribed pathogenic fungus causing dieback of Lantana camara in Iran. Mycol. Prog. 2018, 17, 365–379. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Bhat, D.J.; Chang-Hsin, K.; Hyde, K.D. Leaf litter saprobic Dictyosporiaceae (Pleosporales, Dothideomycetes): Pseudocoleophoma zingiberacearum sp. nov. from Hedychium coronarium. Kavaka 2019, 53, 1–67. [Google Scholar] [CrossRef]
- Bakerspigel, A.; Lowe, D.; Rostas, A. The isolation of Phoma eupyrena from a human lesion. Arch. Dermatol. 1981, 117, 362–363. [Google Scholar] [CrossRef]
- Gkoutselis, G.; Rohrbach, S.; Harjes, J.; Obst, M.; Brachmann, A.; Horn, M.A.; Rambold, G. Microplastics accumulate fungal pathogens in terrestrial ecosystems. Sci. Rep. 2021, 11, 13214. [Google Scholar] [CrossRef]
- Kularathnage, N.D.; Wanasinghe, D.N.; Senanayake, I.C.; Yang, Y.; Manawasinghe, I.S.; Phillips, A.J.L.; Hyde, K.D.; Dong, W.; Song, J. Microfungi associated with ornamental palms: Byssosphaeria phoenicis sp.nov. (Melanommataceae) and Pseudocoleophoma rhapidis sp. nov. (Dictyosporiaceae) from south China. Phytotaxa 2022, 568, 149–169. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Abd-Elsalam, K.A.; Abdel-Wahab, M.A.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Dai, Y.C.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, molecular and human attributes. Fungal Divers. 2015, 74, 18–357. [Google Scholar] [CrossRef]
- de Oliveira, D.A.S.; Debing, Y.; Dieryck, I.; Lyimu, W.M.; Paeshuyse, J. Genome sequences and phylogeny of two duck Hepatitis B viruses. Microbiol. Resour. Announc. 2021, 10, e01327-20. [Google Scholar]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.G.; Ariyawansae, H.A.; Bahkali, A.H.; Elgorban, A.M.; Kang, J.C. A new hysteriform Dothideomycetes (Gloniaceae, Pleosporomycetidae Incertae sedis), Purpurepithecium murisporum gen. et sp.nov.on pine cone scales. Cryptogam. Mycol. 2017, 38, 241–251. [Google Scholar] [CrossRef]
- Valenzuela-Lopez, N.; Cano-Lira, J.F.; Guarro, J.; Sutton, D.A.; Wiederhold, N.; Crous, P.W.; Stchigel, A.M. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud. Mycol. 2018, 90, 1–69. [Google Scholar] [CrossRef]
- Larki, R.; Mehrabi-Koushki, M.; Farokhinejad, R. Ectophoma iranica sp.nov. and new hosts and records of Allophoma spp. in Iran. J. Phytopathol. 2019, 167, 538–545. [Google Scholar] [CrossRef]
- Li, W.J.; McKenzie, E.H.C.; Liu, J.K.; Bhat, D.J.; Dai, D.Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.N.; Luo, Z.L.; Shang, Q.J.; et al. Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar]
- Dong, Z.Y.; Huang, Y.H.; Manawasinghe, I.S.; Wanasinghe, D.N.; Liu, J.W.; Shu, Y.X.; Zhao, M.P.; Xiang, M.M.; Luo, M. Stagonosporopsis pogostemonis: A novel ascomycete fungus causing leaf spot and stem blight on Pogostemon cablin (Lamiaceae) in South China. Pathogens 2021, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.-L.; Chen, L.; Fang, K.; Dong, X.-F.; Li, Y.-X.; Zhang, H.-B.; Yu, Z.-F. Remotididymella ageratinae sp. nov. and Remotididymella anemophila sp.nov., two novel species isolated from the invasive weed Ageratina adenophora in PR China. Int. J. Syst. Evol. Microbiol. 2021, 71, 004572. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A.; Alachiotis, N. Time and memory efficient likelihood-based tree searches on phylogenomic alignments with missing data. Bioinformatics 2010, 26, i132–i139. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.0. Program distributed by the author. In Evolutionary Biology Centre; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2.efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J. Tracer v1, 4. 2017. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 26 August 2022).
- Rambaut, A. FigTree v1. 4.0. A Graphical Viewer of Phylogenetic Trees. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 25 August 2022).
- Schneider, R.; Boerema, G.H. Phoma tropica n. sp., ein a Gewächshauspflanzen häufig vorkommender, nicht pathogener Pilz. Phytopathol. Z. 1975, 83, 239–243. [Google Scholar] [CrossRef]
- Valenzuela-Lopez, N.; Sutton, D.A.; Cano-Lira, J.F.; Paredes, K.; Wiederhold, N.; Guarro, J.; Stchigel, A.M. Coelomycetous fungi in the clinical setting: Morphological convergence and cryptic diversity. J. Clin. Microbiol. 2017, 55, 552–567. [Google Scholar] [CrossRef] [Green Version]
- Zimowska, B. Characteristics and occurrence of Phoma spp. on herbs from the family Lamiaceae. Acta Sci. Pol. Hortorum Cultus 2011, 10, 213–224. [Google Scholar]
- Garibaldi, A.; Gilardi, G.; Ortu, G.; Gullino, M.L. First report of leaf spot of lettuce (Lactuca sativa L.) caused by Phoma tropica in Italy. Plant Dis. 2012, 96, 9. [Google Scholar] [CrossRef]
- Nagarjun, N.; Suryanarayana, V. Documentation, characterization and management of leaf spot of Syzygium cumini (L.) Skeels. J. Farm Sci. 2016, 29, 3. [Google Scholar]
- O’Neill, T.; Mayne, S. An Unusual Phoma stem rot of tomato. AHDB Hortic. 2016, 6–16. [Google Scholar]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Chen, Q.; Hou, L.; Duan, W.; Cai, L. Didymellaceae revisited. Stud. Mycol. 2017, 87, 105–159. [Google Scholar] [CrossRef] [PubMed]
- Gullino, M.L.; Gilardi, G.; Garibaldi, A. Evaluating severity of leaf spot of lettuce, caused by Allophoma tropica, under a climate change scenario. Phytopathol. Mediterr. 2017, 56, 235–241. [Google Scholar]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual. Differentiation of Specific and Infra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004; p. 470. [Google Scholar]
- de Gruyter, J.; Noordeloos, M.E. Contributions towards a monograph of Phoma (Coelomycetes)—I. 1. Section Phoma: Taxa with very small conidia in vitro. Persoonia 1992, 15, 71–92. [Google Scholar]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–24. [Google Scholar] [CrossRef]
- Mathur, P.N.; Thirumalachar, M.J. Studies on some Indian soil fungi 1. Some new or noteworthy Sphaeropsidales. Sydowia 1959, 13, 143–146. [Google Scholar]
- Vaghefi, N.; Pethybridge, S.J.; Ford, R.; Nicolas, M.E.; Crous, P.W.; Taylor, P.W.J. Stagonosporopsis spp. associated with ray blight disease of Asteraceae. Australas. Plant Pathol. 2012, 41, 675–686. [Google Scholar] [CrossRef]
- Vaghefi, N.; Hay, F.S.; Ades, P.K.; Pethybridge, S.J.; Ford, R.; Taylor, P.W.J. Rapid changes in the genetic composition of Stagonosporopsis tanaceti population in Australian Pyrethrum fields. Phytopathology 2015, 105, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Basım, E.; Basım, H.; Abdulai, M.; Baki, D.; Nurhan, Z. Identification and characterization of Didymella bryoniae causing gummy stem blight disease of watermelon (Citrullus lanatus) in Turkey. Crop Prot. 2016, 90, 150–156. [Google Scholar] [CrossRef]
- Zhang, C.; Qian, Y.; Zheng, X.; Zhou, Y.; Xiong, Q. Stagonosporopsis trachelii causes leaf spot on Ningpo Figwort (Scrophularia ningpoensis) in China. Australas. Plant Dis. Notes 2019, 14, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mahapatra, S.; Rao, E.S.; Sandeepkumar, G.M.; Sriram, S. Stagonosporopsis cucurbitacearum the causal agent of gummy stem blight of watermelon in India. Australas. Plant Dis. Notes 2020, 15, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Aveskamp, M.M.; Verkley, G.J.; de Gruyter, J.; Murace, M.A.; Perello, A.; Woudenberg, J.H.; Groenewald, J.Z.; Crous, P.W. DNA phylogenyreveals polyphyly of Phoma section Peyronellaea and multiple taxonomic novelties. Mycologia 2009, 101, 363–382. [Google Scholar] [CrossRef] [Green Version]
- Chethana, K.W.T.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them. Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; de Farias, A.R.G.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Herath, I.S.; Thambugala, K.M.; Manawasinghe, I.S.; et al. What is a species in fungal plant pathogens? Fungal Divers. 2021, 109, 239–266. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Niskanen, T.; Suwannarach, N.; Wannathes, N.; Chen, Y.J.; McKenzie, E.H.; Maharachchikumbura, S.S.; Buyck, B.; Zhao, C.L.; Fan, Y.G.; et al. The numbers of fungi: Are the most speciose genera truly diverse? Fungal Divers. 2022, 114, 387–462. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Phillips, A.J.L.; Pereira, D.S.; Dai, D.-Q.; Aptroot, A.; Monteiro, J.S.; Druzhinina, I.S.; Cai, F.; Fan, X.; Selbmann, L.; et al. Forecasting the number of species of asexually reproducing fungi (Ascomycota and Basidiomycota). Fungal Divers. 2022, 114, 1–28. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Chen, Y.; Ariyawansa, H.A.; Hyde, K.D.; Haelewaters, D.; Perera, R.H.; Samarakoon, M.C.; Wanasinghe, D.N.; Bustamante, D.E.; Liu, J.; et al. Integrative approaches for species delimitation in Ascomycota. Fungal Divers. 2021, 109, 155–179. [Google Scholar] [CrossRef]
- He, Y.L.; He, G.; Li, Q.Q.; Lin, W.; Yuan, G.Q. First report of Stagonosporopsis vannaccii causing leaf spot on Crassocephalum crepidioides in China. Plant Dis. 2020, 105, 1–499. [Google Scholar] [CrossRef]
- Zhao, Q.; Wu, J.; Zhang, L.; Xu, L.; Yan, C.; Gong, Z. Identification and characteristics of Stagonosporopsis cucurbitacearum pathogenic factors influencing pumpkin seeding survival in northeast China. J. Phytopathol. 2018, 167, 41–55. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Region | Primer Pairs | Optimized Thermal Cycler Conditions |
|---|---|---|
| ITS | ITS1/ITS4 | 94 °C: 3 min; (94 °C: 45 s, 56 °C: 50 s) 35 cycles 72 °C: 10 min |
| LSU | LR0R/LR5 | 94 °C: 3 min; (94 °C: 45 s, 56 °C: 50 s) 35 cycles 72 °C: 10 min |
| RPB2 | fRPB2-5F/fRPB2-7cR | 95 °C: 5 min; (95 °C: 1 min, 52 °C: 50 s) 35 cycles 72 °C: 10 min |
| β-tubulin | bt2a/bt2b | 95 °C: 5 min; (94 °C: 1 min, 55 °C: 90 s) 35 cycles 72 °C: 10 min |
| Species | Strain no | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| ITS | LSU | RPB2 | β-tubulin | ||
| Allophoma anatiae | CBS 124673 | MN973472 | MN943674 | MT018048 | MT005571 |
| Allophoma cylindrispora | UTHSC DI16-233 | LT592920 | LN907376 | LT593058 | LT592989 |
| Allophoma zantedeschiae | CBS 131.93 | FJ427084 | GU238159 | KT389557 | FJ427188 |
| Allophoma hayatii | CBS 142859 | KY684812 | KY684814 | MF095108 | KY684816 |
| Allophoma labilis | CBS 124.93 | GU237765 | GU238091 | KT389552 | GU237619 |
| Allophoma piperis | CBS 268.93 | GU237816 | GU238129 | KT389554 | GU237644 |
| Allophoma siamensis | MFLU 17-2422 | MK347742 | MK347959 | MK434912 | MK412867 |
| Allophoma pterospermicola | LC12182 | MK088570 | MK088577 | MK088584 | MK088591 |
| Allophoma pterospermicola | CGMCC 3.19245 | MK088573 | MK088580 | MK088587 | MK088594 |
| Allophoma thunbergiae | GUCC2070.7 | MW036298 | MW040201 | MW116819 | MW116823 |
| Allophoma minor | CBS 325.82 | GU237831 | GU238107 | KT389553 | GU237632 |
| Allophoma alba | CBS 120422 | MN973469 | MN943671 | MT018044 | MT005568 |
| Allophoma nicaraguensis | CBS 506.91 | GU237876 | GU238058 | KT389551 | GU237596 |
| Allophoma tropica | CBS 436.75 | GU237864 | GU238149 | KT389556 | GU237663 |
| Allophoma tropica | CBS 537.66 | MH858877 | MH870533 | MT018043 | N/A |
| Allophoma tropica | ZHKUCC 22-0169 | OQ275206 | OQ275192 | OQ343373 | OQ336257 |
| Allophoma tropica | ZHKUCC 22-0170 | OQ275207 | OQ275193 | OQ343374 | OQ336258 |
| Allophoma oligotrophica | LC 6245 | KY742040 | KY742194 | KY742128 | KY742282 |
| Ectophoma insulana | CBS 140548 | MN973482 | MN943686 | MT018071 | MT005582 |
| Ectophoma insulana | CBS 252.92 | MN973481 | MN943685 | MT018070 | MT005581 |
| Ectophoma pomi | CBS 267.92 | GU237814 | GU238128 | N/A | GU237643 |
| Ectophoma iranica | SCUA-K1G1 | MK519382 | MK519389 | N/A | MK519562 |
| Ectophoma pomi | CBS 121.93 | MN972933 | N/A | MN983570 | MN983948 |
| Ectophoma multirostrata | CBS 110.79 | FJ427030 | GU238110 | LT623264 | FJ427140 |
| Ectophoma multirostrata | CBS 274.60 | FJ427031 | GU238111 | LT623265 | FJ427141 |
| Ectophoma sp. | URM 408 | MH384820 | MH370603 | MH370607 | N/A |
| Ectophoma phoenicis | ZHKUCC 22-0163 | OQ275208 | OQ275194 | OQ343375 | OQ336259 |
| Ectophoma phoenicis | ZHKUCC 22-0164 | OQ275209 | OQ275195 | OQ343376 | OQ336260 |
| Remotididymella ageratinae | G13388 | MN864294 | MN864298 | MN871530 | MN871533 |
| Remotididymella anemophila | C74 | MN864293 | MN864296 | MN871529 | MN871532 |
| Remotididymella anthropophila | CBS 142462 | LT592936 | N/A | LT593075 | LT593005 |
| Remotididymella bauhiniae | MFLU 18-2118 | MK347737 | MK347954 | MK434914 | MK412884 |
| Remotididymella brunnea | CBS 993.95 | NR_170781 | N/A | MT018064 | MT005576 |
| Remotididymella capsici | CBS 679.77 | MN973478 | MN943681 | MT018066 | MT005578 |
| Remotididymella destructiva | CBS 133.93 | GU237779 | GU238064 | LT623257 | GU237602 |
| Remotididymella fici-microcarpae | ZHKUCC 22-0165 | OQ275210 | OQ275196 | OQ343377 | OQ336261 |
| Remotididymella fici-microcarpae | ZHKUCC 22-0166 | OQ275211 | OQ275197 | OQ343378 | OQ336262 |
| Remotididymella humicola | CBS 120117 | NR_170782 | MN943680 | MT018065 | MT005577 |
| Stagonosporopsis actaeae | CBS 106.96 | GU237734 | GU238166 | KT389672 | GU237671 |
| Stagonosporopsis actaeae | CBS 105.96 | GU237733 | GU238165 | MT018018 | GU237670 |
| Stagonosporopsis ailanthicola | MFLUCC 16-1439 | KY100872 | KY100874 | KY100876 | KY100878 |
| Stagonosporopsis ajacis | CBS 176.93 | GU237790 | GU238167 | MT018035 | GU237672 |
| Stagonosporopsis andigena | CBS 269.80 | GU237817 | GU238170 | MT018026 | GU237675 |
| Stagonosporopsis artemisiicola | CBS 102636 | GU237728 | GU238171 | KT389674 | GU237676 |
| Stagonosporopsis astragali | CBS 178.25 | GU237792 | GU238172 | MT018030 | GU237677 |
| Stagonosporopsis bomiensis | CGMCC 3.18366 | KY742123 | KY742277 | KY742189 | KY742365 |
| Stagonosporopsis chrysanthemi | CBS 500.63 | GU237871 | GU238190 | MT018012 | GU237695 |
| Stagonosporopsis chrysanthemi | CBS 137.96 | GU237783 | GU238191 | MT018011 | GU237696 |
| Stagonosporopsis citrulli | FLAS-F-58996 | KJ855546 | N/A | N/A | KJ855602 |
| Stagonosporopsis crystalliniformis | CBS 713.85 | GU237903 | GU238178 | KT389675 | GU237683 |
| Stagonosporopsis cucumeris | CBS 386.65 | MN973455 | MN943657 | MT018021 | MT005554 |
| Stagonosporopsis caricae | CBS 119735 | MN973042 | N/A | MN983680 | MN984054 |
| Stagonosporopsis caricae | CBS 102399 | MN973041 | N/A | MN983679 | MN984053 |
| Stagonosporopsis dennisii | CBS 110.96 | MN973046 | N/A | MN983685 | MN984058 |
| Stagonosporopsis dorenboschii | CBS 320.90 | GU237830 | GU238184 | MT018039 | GU237689 |
| Stagonosporopsis helianthi | CBS 200.87 | KT389545 | KT389761 | KT389683 | KT389848 |
| Stagonosporopsis hortensis | CBS 572.85 | GU237893 | GU238199 | KT389681 | GU237704 |
| Stagonosporopsis inoxydabilis | CBS 425.90 | GU237861 | GU238188 | KT389682 | GU237693 |
| Stagonosporopsis loticola | CBS 562.81 | GU237890 | GU238192 | KT389684 | GU237697 |
| Stagonosporopsis lupini | CBS 101494 | GU237724 | GU238194 | KT389685 | GU237699 |
| Stagonosporopsis nemophilae | CBS 715.85 | MN973460 | MN943662 | MT018031 | MT005559 |
| Stagonosporopsis oculo-hominis | CBS 634.92 | GU237901 | GU238196 | KT389686 | GU237701 |
| Stagonosporopsis papillata | LC 8171 | KY742127 | KY742281 | KY742193 | KY742369 |
| Stagonosporopsis pini | MFLUCC 18-1549 | MK347800 | MK348019 | MK434860 | MK412886 |
| Stagonosporopsis rudbeckiae | CBS 109180 | GU237745 | GU238197 | MT018015 | GU237702 |
| Stagonosporopsis sambucella | CBS 130003 | MN973459 | MN943661 | MT018029 | MT005558 |
| Stagonosporopsis stuijvenbergii | CBS 144953 | MN823449 | MN823300 | MN824475 | MN824623 |
| Stagonosporopsis tanaceti | CBS 131484 | NR_111724 | KP161044 | MT018013 | JQ897496 |
| Stagonosporopsis pedicularis-striatae | ZHKUCC 22-0167 | OQ275212 | OQ275198 | OQ343379 | OQ336263 |
| Stagonosporopsis pedicularis-striatae | ZHKUCC 22-0168 | OQ275213 | OQ275199 | OQ343380 | OQ336264 |
| Stagonosporopsis trachelii | CBS 379.91 | GU237850 | GU238173 | KT389687 | GU237678 |
| Stagonosporopsis valerianellae | CBS 273.92 | GU237819 | GU238200 | MT018033 | GU237705 |
| Stagonosporopsis weymaniae | CBS 144959 | MN823453 | MN823304 | MN824479 | MN824627 |
| Stagonosporopsis centaureae | MFLUCC 16-0787 | KX611240 | KX611238 | N/A | N/A |
| Stagonosporopsis heliopsidis | CBS 109182 | GU237747 | GU238186 | KT389679 | GU237691 |
| Stagonosporopsis pogostemonis | ZHKUCC 21-0001 | MZ156571 | MZ191532 | MZ203135 | MZ203132 |
| Stagonosporopsis rhizophila | CGMCC 3.19852 | MG833824 | MG833789 | MN422105 | MN422099 |
| Stagonosporopsis cucurbitacearum | CBS 133.96 | GU237780 | GU238181 | KT389676 | GU237686 |
| Stagonosporopsis cucurbitacearum | CBS 109171 | GU237922 | GU238180 | N/A | GU237685 |
| Stagonosporopsis ailanthicola | CBS 140554 | MN973462 | MN943664 | MT018036 | MT005561 |
| Epicoccum oryzae | CBS 173.34 | MN973499 | N/A | MT018098 | MT005599 |
| Epicoccum viticis | LC 5126 | KY742118 | N/A | KY742186 | KY742360 |
| Similiphoma crystallifera | CBS 193.82 | GU237797 | GU238060 | LT623267 | GU237598 |
| Didymella heteroderae | CBS 109.92 | FJ426983 | GU238002 | KT389601 | FJ427098 |
| Didymella suiyangensis | LC 7439 | KY742089 | KY742243 | KY742168 | KY742330 |
| Cumuliphoma indica | CBS 65478 | FJ427044 | GU238123 | LT623262 | FJ427154 |
| Cumuliphoma pneumoniae | UTHSC DI16-249 | NR_158277 | N/A | LT593063 | LT592994 |
| Paraboeremia selaginellae | CBS 122.93 | GU237762 | GU238142 | MT018189 | GU237656 |
| Paraboeremia litseae | CGMCC 318109 | KX829029 | KX829037 | KX829045 | KX829053 |
| Macroventuria anomochaeta | CBS 502.72 | GU237873 | GU237985 | MT018193 | GU237545 |
| Macroventuria wentii | CBS 526.71 | GU237884 | GU237986 | KT389642 | GU237546 |
| Juxtiphoma eupyrena | CBS 527.66 | FJ427000 | GU238073 | LT623269 | FJ427111 |
| Juxtiphoma eupyrena | CBS 374.91 | FJ426999 | GU238072 | LT623268 | FJ427110 |
| Vacuiphoma ferulae | CBS 353.71 | MH860160 | MH871928 | MT018196 | MT005655 |
| Vacuiphoma oculihominis | FMR 138.01 | LT592954 | N/A | LT593093 | LT593023 |
| Nothophoma infossa | CBS 123395 | FJ427025 | FJ899743 | KT389659 | FJ427135 |
| Nothophoma anigozanthi | CBS 381.91 | GU237852 | GU238039 | KT389655 | GU237580 |
| Ascochyta medicaginicola | BRIP 45051 | KY742044 | KY742198 | KY742132 | KY742286 |
| Ascochyta koolunga | CBS 373.84 | KT389481 | KT389698 | KT389560 | KT389775 |
| Calophoma rosae | CGMCC 3.18347 | KY742049 | KY742203 | KY742135 | KY742291 |
| Calophoma vodakii | CBS 173.53 | KT389497 | KT389714 | MT018233 | KT389791 |
| Neomicrosphaeropsis novorossica | MFLUCC 14-0578 | KX198709 | KX198710 | N/A | N/A |
| Neomicrosphaeropsis rossica | MFLUCC 14-0586 | KU752192 | KU729855 | N/A | N/A |
| Phoma herbarum | CBS 127589 | KT389539 | MH876049 | KT389664 | KT389838 |
| Phoma herbarum | CBS 134.96 | KT389535 | KT389753 | KT389661 | KT389834 |
| Phomatodes nebulosa | CBS 740.96 | KT389540 | KT389758 | KT389667 | KT389839 |
| Leptosphaerulina americana | CBS 213.55 | GU237799 | GU237981 | KT389641 | GU237539 |
| Leptosphaerulina arachidicola | CBS 275.59 | GU237820 | GU237983 | MT018278 | GU237543 |
| Pseudoascochyta novaezelandiea | CBS 141689 | LT592892 | LT592893 | LT592895 | LT592894 |
| Pseudoascochyta pratensis | CBS 141688 | LT223130 | LT223131 | LT223133 | LT223132 |
| Briansuttonomyces eucalypti | CBS 114879 | KU728479 | KU728519 | MT018239 | KU728595 |
| Xenodidymella applanata | CBS 115577 | KT389546 | KT389762 | KT389688 | KT389850 |
| Xenodidymella asphodeli | CBS 375.62 | KT389549 | KT389765 | KT389689 | N/A |
| Neodidymelliopsis cannabis | CBS 121.75 | GU237761 | GU237972 | N/A | GU237535 |
| Neodidymelliopsis achlydis | CBS 256.77 | KT389531 | KT389749 | MT018293 | KT389829 |
| Neoascochyta europaea | CBS 819.84 | KT389510 | KT389728 | KT389645 | KT389808 |
| Neoascochyta graminicola | CBS 102789 | KT389518 | KT389736 | KT389649 | KT389816 |
| Leptosphaeria conoidea | CBS 616.75 | JF740201 | JF740279 | KT389639 | KT389804 |
| Heterophoma poolensis | CBS 113.20 | GU237751 | GU238119 | MT018056 | GU237638 |
| Boeremia diversispora | CBS 101194 | GU237716 | GU237929 | KT389564 | GU237491 |
| Boeremia foveata | CBS 109176 | GU237742 | GU237946 | KT389578 | GU237508 |
| Leptosphaeria doliolum | CBS 505.75 | JF740205 | GU301827 | KT389640 | JF740144 |
| Heterophoma nobills | CBS 507.91 | GU237877 | GU238065 | KT389638 | GU237603 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kularathnage, N.D.; Senanayake, I.C.; Wanasinghe, D.N.; Doilom, M.; Stephenson, S.L.; Song, J.; Dong, W.; Xu, B. Plant-Associated Novel Didymellaceous Taxa in the South China Botanical Garden (Guangzhou, China). J. Fungi 2023, 9, 182. https://doi.org/10.3390/jof9020182
Kularathnage ND, Senanayake IC, Wanasinghe DN, Doilom M, Stephenson SL, Song J, Dong W, Xu B. Plant-Associated Novel Didymellaceous Taxa in the South China Botanical Garden (Guangzhou, China). Journal of Fungi. 2023; 9(2):182. https://doi.org/10.3390/jof9020182
Chicago/Turabian StyleKularathnage, Nuwan D., Indunil C. Senanayake, Dhanushka N. Wanasinghe, Mingkwan Doilom, Steven L. Stephenson, Jiage Song, Wei Dong, and Biao Xu. 2023. "Plant-Associated Novel Didymellaceous Taxa in the South China Botanical Garden (Guangzhou, China)" Journal of Fungi 9, no. 2: 182. https://doi.org/10.3390/jof9020182