
The Potential Fungal Pathogens of Euonymus japonicus in Beijing, China


1
The Key Laboratory for Silviculture and Conservation of Ministry of Education, Beijing Forestry University, Beijing 100083, China
2
The Key Laboratory for Biodiversity Science and Ecological Engineering, College of Life Sciences, Beijing Normal University, Beijing 100101, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(2), 271; https://doi.org/10.3390/jof9020271
Submission received: 17 January 2023
/
Revised: 6 February 2023
/
Accepted: 16 February 2023
/
Published: 18 February 2023
(This article belongs to the Special Issue Taxonomy, Systematics and Evolution of Forestry Fungi)
Abstract
:Euonymus japonicus tolerates the dry and frigid climate of Beijing, China, and effectively filters out particles during the winter. However, fungal infestation frequently causes extreme illness and can even lead to shrub death. In this study, 104 diseased E. japonicus specimens were collected from seven districts in Beijing. Seventy-nine isolates were identified as 22 fungal species in seven genera. The species were Aplosporella hesperidica, A. javeedii, A. prunicola, Botryosphaeria dothidea, Colletotrichum aenigma, Co. euonymi, Co. euonymicola, Co. gloeosporioides, Cytospora ailanthicola, C. albodisca, C. diopuiensis, C. discotoma, C. elaeagni, C. euonymicola, C. euonymina, C. haidianensis, C. leucostoma, C. sophorae, C. zhaitangensis, Diaporthe eres, Dothiorella acericola, and Pestalotiopsis chaoyangensis. On the basis of morphological and phylogenetic analyses, Colletotrichum euonymi, Co. euonymicola, Cytospora zhaitangensis, and Pestalotiopsis chaoyangensis were introduced as novel species. Colletotrichum euonymi, Co. euonymicola, and Pestalotiopsis chaoyangensis were subsequently confirmed as pathogens of E. japonicus leaves by pathogenicity testing. This study provides an important assessment of the fungi associated with diseases of E. japonicus in Beijing, China.
1. Introduction
The family Celastraceae, which includes 96 genera and over 1350 species, is extensively distributed in tropical, subtropical, and temperate climates as evergreen or deciduous trees, shrubs, or vine shrubs [1]. Euonymus japonicus, also called Japanese spindle, is one of the most prevalent and important species in Celastraceae in northern Chinese cities such as Beijing, the capital of China [2]. As a species of evergreen shrub, E. japonicus has strong resistance to the dry and cold conditions in Beijing and can efficiently reduce particulate matter in winter [2]. Furthermore, roots, stems, and leaves of the shrub have a high capacity to enrich heavy metals [3]. However, Japanese spindle in Beijing is frequently extremely ill and even dies because of fungal infestation (Figure 1).
Fungi from many different taxa are associated with diseases of the same host plant species or genus. Raza et al. [4] described one new genus and 32 new species of culturable fungi associated with sugarcane disease in southern China, and Crous et al. [5] described seven new genera and 15 new species as foliar fungal pathogens of eucalypts. Accurate identification of pathogenic fungi provides a good theoretical foundation for the control of plant diseases. However, because fungi associated with fungal diseases of Japanese spindle in Beijing have not been systematically and extensively studied, effective disease prevention is difficult. Therefore, the variety of fungal species associated with Japanese spindle diseases in Beijing was examined in this study. In the study, 79 fungal isolates were classified as seven genera (Aplosporella, Botryosphaeria, Collectotrichum, Cytospora, Diaporthe, Dothiorella, and Pestalotiopsis) in four orders (Amphisphaeriales, Botryosphaeriales, Diaporthales, and Glomerellales).
Aplosporella, Botryosphaeria, and Dothiorella are genera in Botryosphaeriales and Dothideomycetes [6]. Botryosphaeriales is an order that includes many pathogens to various hosts [7,8]. Currently, six families (Aplosporellaceae, Botryosphaeriaceae, Melanopsaceae, Phyllostictaceae, Planistromellaceae, and Saccharataceae) and 33 genera are accepted in Botryosphaeriales [6,9]. Aplosporella is in Aplosporellaceae, and Botryosphaeria and Dothiorella are in Botryosphaeriaceae. The two families can be distinguished by the locules, with Aplosporellaceae multiloculate and Botryosphaeriaceae uniloculate [8]. Conidia of Botryosphaeria are aseptate, whereas those of Dothiorella are septate [10]. Cash [11] recorded A. euonymi on Euonymus atropurpureus. Hanlin [12] and Reid [13] reported that B. dothidea and B. quercuum were associated with E. americanus and E. japonicus, respectively. Dissanayake et al. [14] reported that Do. sarmentorum was associated with E. europaeus.
Colletotrichum is in Glomerellales, Sordariomycetes. The genus comprises 15 species complexes, i.e., the species complexes of Co. acutatum, Co. agaves, Co. boninense, Co. caudatum, Co. dematium, Co. destructivum, Co. dracaenophilum, Co. gigasporum, Co. gloeosporioides, Co. graminicola, Co. magnum, Co. orbiculare, Co. orchidearum, Co. spaethianum, and Co. truncatum [15,16]. Internal transcribed spacer region (ITS) is always used to assign Colletotrichum species to species complexes [16,17]. Liu et al. [16] combined ITS, act, chs-1, gapdh, his3, and tub2 sequences to construct a phylogenetic tree of Colletotrichum species. In addition, ITS, apn2, Mat1/Apn2, and sod2 sequences were combined in phylogenetic analyses of the Co. caudatum species complex, and ITS, act, chs, sod2, and tub2 sequences were combined in phylogenetic analyses of the Co. graminicola species complex [16]. Colletotrichum boninense, Co. gigasporum, Co. gloeosporioides, Co. griseum, Co. karsti, and Co. siamense have been recorded on host Euonymus [18,19,20,21,22].
Cytospora and Diaporthe are genera in Diaporthales, Sordariomycetes. Diaporthales includes many pathogens that cause dieback and canker disease in various hosts [23,24,25,26,27,28,29]. Cytospora euonymi, C. euonymicola, C. euonymina, C. haidianensis, and C. leucostoma have been recorded on host Euonymus [30,31,32,33]. Diaporthe eres, D. euonymi, D. laschii, and D. pardalota have been recorded on host Euonymus [34,35,36,37].
Pestalotiopsis is in Amphisphaeriales, Sordariomycetes. Steyaert [38] distinguished the genus from Pestalotia on the basis of its morphological features (five-celled, clostridial conidia with colored cells in the middle three cells and colorless apical cells, and one to several apical appendages). Pestalotiopsis is a well-known phytopathogenic genera. Jiang et al. [39] introduced 10 new species of Pestalotiopsis from Fagaceae leaves in China. Pestalotiopsis breviseta, P. caroliniana, P. clavata, P. diospyri, P. gracilis, P. neglecta, and P. planimi have been recorded on host Euonymus [40,41,42,43].
During investigations of the diversity of fungal species that cause diseases of E. japonicus, several ascomycetous taxa associated with various disease symptoms were collected in Beijing. The objectives of this study are to (1) investigate fungal diseases on E. japonicus in Beijing, (2) identify the fungal species isolated from E. japonicus, and (3) test the pathogenicity of the novel species identified.
2. Materials and Methods
2.1. Sampling and Isolation
During 2020 to 2021, a survey was conducted in seven districts (Chaoyang, Daxing, Fengtai, Haidian, Mentougou, Shijingshan, and Xicheng) in Beijing, China. A total of 104 specimens (67 branches and 37 leaves) affected with different symptoms were collected. Isolates from leaves were obtained using tissue isolation methods. Leaf spots were cut into small pieces (0.2 × 0.2 cm) and placed on potato dextrose agar (PDA, 200 g potato, 20 g glucose, 20 g agar, and 1000 mL water) plates and incubated at 25 °C after surface sterilisation (1 min in 75% ethanol, 3 min in 1.25% sodium hypochlorite, then rinsed in distilled water and blotted on dry sterile filter paper). Fruiting bodies on diseased branches were shaved off the surface with a sterile blade after surface sterilisation, then the mucoid spore mass from conidiomata was put onto a PDA culture medium and incubated at 25 °C in darkness until spores germinated. Single germinating conidia were transferred onto fresh PDA plates. Hyphal tips were cut and transferred to a new PDA plate twice to obtain a pure culture for further study. Specimens are preserved at the working collection of X.L. Fan (CF) housed at the Beijing Forestry University (BJFC). Cultures of taxonomic novelties are deposited at the China Forestry Culture Collection Centre (CFCC).
2.2. DNA Extraction, PCR Amplification, and Sequencing
The modified CTAB method [44] was used to extract total genomic DNA from mycelium on PDA. The PCR mixture consisted of 10 μL TopTaq™ Master Mix, 7 μL nuclease-free H2O, 1 μL of each primer, and 1 μL DNA samples were made up to the final volume of 20 μL. Partial gene sequences were amplified by polymerase chain reaction (PCR) using the primer sets ITS1/ITS4 for ITS region [45], EF-728F/EF-986R for tef1-α [46], Bt-2a/Bt-2b for tub2 [47], fRPB2-5F/fRPB2-7cR for rpb2 [48], ACT-512F/ACT-783R for act [46], CHS-79F/CHS-345R for chs1 [46], GDF1/GDF2 for gaphd [49], CAL-228F/CAL-737R for cal [46], and CYLH3F/H3-1b [47,50] for his3. The genes used in different genera and the PCR conditions are listed in Table 1.
PCR products were electrophoresed in 1% agarose gel, and the DNA was sequenced by the SinoGenoMax Company Limited (Beijing, China). The forward and reverse reads were assembled by Seqman v. 7.1.0 in the DNASTAR Lasergene core suite software (DNASTAR Inc., Madison, WI, USA). All sequences obtained in this study were submitted to GenBank (Table S1).
2.3. Phylogenetic Analyses
The sequence datasets used in this study were based on Lin et al. [51,52] for Cytospora, Liu et al. [16] for Colletotrichum, Jiang et al. [39] for Pestalotiopsis, and Zhang et al. [6] for Botryosphariales, deleting the overly repetitive isolates of the same species and supplementing them with other sequences obtained from GenBank (Table S1). Sequence alignments of the individual loci were performed in MAFFT v. 6 [53] and adjusted by MEGA v. 6.0 [54]. Ambiguous regions were excluded from alignments. For the genus Colletotrichum, the ITS tree, including 15 species complexes, was first used for inferring delimitation to the species complex level before multi-locus phylogenetic analyses. Maximum Likelihood (ML) and Bayesian Inference (BI) were used for phylogenetic analyses of both each individual loci and the concatenated genes alignments. ML and BI analyses were computed using PhyML v. 3.0 [55] and MrBayes v. 3.1.2 [56], respectively. For ML analyses, GTR + GAMMA model of site substitution with 1000 bootstrap was set. For BI analyses, the best-fit evolutionary models for each partitioned locus were estimated in MrModeltest v. 2.4 [57]. BI analyses with a four simultaneous Markov Chain Monte Carlo (MCMC) were computed from random trees for 1,000,000 generations and sampled every 1000 generations, and the burn-in was set to 0.25. The resulting trees were viewed in Figtree v. 1.3.1 [58]. The multi-locus sequence alignments were deposited in TreeBASE 29991.
2.4. Morphology
For the isolates isolated from diseased branches, the fruiting bodies on the specimens corresponding to the isolates were used for morphological observation. For the isolates isolated from leaf spots, reproductive structures formed on PDA were used for morphological observation. The structure and size of conidiomata were photographed using the Leica stereomicroscope (M205 FA) (Leica Microsystems, Wetzlar, Germany). Over 30 conidiomata were sectioned, and 50 conidia were selected randomly to measure their lengths and widths using a Nikon Eclipse 80 i microscope (Nikon Corporation, Tokyo, Japan) equipped with a Nikon digital sight DS-Ri2 high-definition color camera with differential interference contrast (DIC). Colony color on PDA were described according to the color charts of Rayner [59].
2.5. Pathogenicity Tests
Healthy branches and leaves of E. japonicus collected from Beijing Forestry University (116°20′28″ E, 40°0′8″ N) were used to confirm the pathogenicity of the ex-holotype of the four novel species (isolated from branches: C. zhaitangensis; and isolated from leaves: Co. euonymi, Co. euonymicola, and P. chaoyangensis). Two-year-old branches were inoculated with C. zhaitangensis (CFCC 56227), and one-year-old leaves were inoculated with Co. euonymi (CFCC 55542), Co. euonymicola (CFCC 55486), and P. chaoyangensis (CFCC 55549).
Branches were cut to 20-cm lengths, with the bottoms submerged in water and tips sealed with sealing film. To inoculate branches, a hole punch with 5-mm diameter and approximately 0.5-mm thickness was used to scald branches 10 cm from the tip. A 14-day culture block of the same size was attached to the wounds. Branches were then wet with skimming cotton moistened with sterile water and covered in sealing film. To inoculate leaves, a sterile inoculation needle pierced the leaves five to seven times, and 4-mm-diameter 14-day culture blocks were placed on the wounds. Branch and leaf inoculations were incubated at 25 °C and 70% humidity. Six replicates were prepared for each isolate, and a sterile PDA plug served as the control. Experiments were conducted twice. To fulfil Koch’s postulates, re-isolations were made from lesions to compare the morphological features and DNA sequences with those of the original isolates.
3. Results
3.1. Disease Investigation and Isolation
Symptoms on leaves of infected plants included necrotic patches with tiny, black dot-like infection centers. Branch tips were depressed and dead and spread downward. Black reproductive structures appeared on dead portions of the branches (Figure 2). A total of 79 isolates were obtained, with 26 isolates from Chaoyang, 15 from Haidian, 14 from Mentougou, 10 from Shijingshan, five from Daxing, five from Xicheng, and four from Fengtai districts.
3.2. Phylogenetic Analyses
The best-fit models used in Bayesian analyses and the statistics of ML trees are presented in Table 2 and Table 3, respectively. The ML trees with ML bootstrap support values and posterior probabilities are shown in Figure 3, Figure 4 and Figure 5 and Figures S1–S5. The results of Bayesian analyses did not significantly differ from those of ML trees.
The 79 isolates clustered into seven genera, i.e., Aplosporella, Botryosphaeria, Collectotrichum, Cytospora, Diaporthe, Dothiorella, and Pestalotiopsis. Eight isolates in the genus Aplosporella were clustered in three clades with A. hesperidica, A. javeedii, and A. prunicola (Figure S1). Twenty-one isolates in the genus Botryosphaeria clustered with B. dothidea (Figure S2). The ten isolates in Colletotrichum all clustered in the Co. gloeosporioides complex based on the ITS tree (Figure S3). Multilocus analyses in the Co. gloeosporioides complex showed that the ten isolates clustered in four clades (Figure 3). Isolate CFCC 55535 clustered with Co. aenigma, and three isolates (CFCC 55544, 55545 and 55547) clustered with Co. gloeosporioides. Four isolates (CFCC 55483, 55537, 55540, and 55542) and two isolates (CFCC 55486 and 55539) formed two distinct branches. A total of 31 isolates in the genus Cytospora clustered in 11 clades, with 10 known species (C. ailanthicola, C. albodisca, C. diopuiensis, C. discotoma, C. elaeagni, C. euonymicola, C. euonymina, C. haidianensis, C. leucostoma, and C. sophorae), and one forming a distinct branch (CFCC 56227 and 57537) (Figure 4). Three isolates in Diaporthe were clustered in the D. alnea species complex (Figure S4). Four isolates in Dothiorella clustered with Do. acericola (Figure S5). Two isolates in Pestalotiopsis (CFCC 55549 and 58805) were placed in a distinct clade (Figure 5).
3.3. Taxonomy
Dothideomycetes O.E. Erikss. and Winka, Myconet 1 (1): 5, 1997.
Botryosphaeriales C.L. Schoch, Crous and Shoemaker Mycologia 98 (6): 1050, 2007.
Aplosporella Speg., Anal. Soc. cient. argent. 10 (5–6): 158, 1880.
Aplosporella hesperidica Speg., Anal. Soc. cient. argent. 13 (1): 18, 1882.
Materials examined: China. Beijing City, Haidian District, Shucun Park, 116°17′55″ E, 40°0′53″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (BJFC CF20220104, living culture CFCC 55554).
Notes: Aplosporella hesperidica was first discovered by Spegazzini [63] on Citrus × aurantium in Argentina. The isolate CFCC 55554 clustered with Aplosporella hesperidica CBS 732.79 and A. hesperidica CBS 208.37 in the present phylogenetic analysis and showed 100% similarity in ITS sequence. Therefore, CFCC 55554 is identified as Aplosporella hesperidica according to phylogenetic analyses. A new host record from Euonymus japonicus is provided here.
Aplosporella javeedii Jami, Gryzenh., Slippers and M.J. Wingf., Fungal Biol. 118 (2): 174, 2013.
Materials examined: China. Beijing City, Chaoyang District, Sun Palace South Street, 116°26′57″ E, 39°58′10″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220101, living culture CFCC 55489). Mentougou District, Shitan Road, 116°6′31″ E, 39°55′36″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 25 May 2021 (BJFC CF20220102, living culture CFCC 55553). Haidian District, Shucun Park, 116°17′55″ E, 40°0′53″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 7 June 2021 (BJFC CF20220103, living culture CFCC 55555).
Notes: Aplosporella javeedii was introduced by Jami et al. [64] from a healthy wood section of Celtis Africana. It has been reported on 14 different hosts in 10 families, such as Rhamnaceae, Rosaceae, and Cupressaceae [65,66,67,68,69]. The isolates CFCC 55489, 55553, and 55555 were grouped together with A. javeedii (CFCC 50052 and 50054) with high statistical support (ML/BI = 99/1). The conidia of CFCC 55489 on PDA are aseptate, ellipsoid to oblong, smooth, ends rounded, initial hyaline, becoming brown when mature, 19.5–23.0 × 6.5–8.5 µm, which overlap with the morphological characteristics described by Jami et al. [64]. Therefore, three isolates in this study are identified as A. javeedii based on phylogenetic and morphological analyses.
Aplosporella prunicola Damm and Crous, Fungal Divers. 27 (1): 39, 2007.
Descriptions: See Damm et al. [70].
Material examined: China. Beijing City, Chaoyang District, Sanyuanli, 116°26′59″ E, 39°57′10″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 20 April 2021 (BJFC CF20220105, living culture CFCC 55550). Mentougou District, Lisichen Park, 116°6′33″ E, 39°55′39″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (BJFC CF20220106, living culture CFCC 55551). Fengtai District, West Bureau Yupu Garden, 116°17′32″ E, 39°52′10″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 April 2021 (BJFC CF20220107, living culture CFCC 55552 and 57541).
Notes: In this study, four isolates CFCC 55550–55552 and 57541 were grouped together with Aplosporella prunicola and A. yalgorensis in phylogenetic analyses with 98 ML bootstrap support value and 0.98 posterior probabilities. The four isolates in this study can be distinguished from A. yalgorensis based on ITS and tef loci (for 10–11/520 bp in ITS, 2–6/317 bp in tef). In ML tree, CFCC 55550–55551 clustered with A. prunicola with 83 ML bootstrap support value with 100% repetitive ITS sequences. CFCC 55552 and 57541 are only one base different from A. prunicola in ITS region. Additionally, the conidia size of CFCC 55552 on PDA are 16.5–21.5 × 10.0–10.5 µm, which is consistent with the morphological characteristics of ex-type of A. prunicola for (17) 19–22 (25) × (9) 10–12 (18) [70]. Therefore, these four isolates in this study are identified as Aplosporella prunicola.
Botryosphaeria Ces. and De Not., Comment. Soc. Crittog. Ital. 1: 211. 1863.
Botryosphaeria dothidea (Moug.: Fr.) Ces. and De Not., Comment. Soc. Crittog. Ital. 1: 212. 1863.
Basionym: Sphaeria dothidea Moug., In: Fries, Syst. Mycol. (Lundae) 2 (2): 423. 1823
Description: See Phillips et al. [11].
Material examined: China. Beijing City, Chaoyang District, Olympic Forest Park, 116°23′9″ E, 40°0′2″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 26 April 2021 (BJFC CF20220110, living culture CFCC 55492–55496); 116°23′10″ E, 40°0′7″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 26 April 2021 (BJFC CF20220115, living culture CFCC 55569–55572); 116°23′10″ E, 40°0′2″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (BJFC CF20220119, living culture CFCC 55681). Haidian District, Beijing Forestry University, 116°20′28″ E, 40°0′8″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 April 2021 (BJFC CF20220108, living culture CFCC 55490; BJFC CF20220109, living culture CFCC 55491). Shijingshan District, Sculpture Garden Middle Street, 116°14′40″ E, 39°54′23″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 May 2021 (BJFC CF20220111, living culture CFCC 55564; BJFC CF20220117, living culture CFCC 55576–55577); 116°14′35″ E, 39°54′23″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 May 2021 (BJFC CF20220112, living culture CFCC 55565). Daxing District, Daxing New Town Riverfront Forest Park, 116°17′41″ E, 39°44′31″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 May 2021 (BJFC CF20220113, living culture CFCC 55566); 116°17′51″ E, 39°44′31″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 May 2021 (BJFC CF20220114, living culture CFCC 55567–55568). Mentougou District, Pushan Park, 116°6′35″ E, 39°55′30″ N, from diseased leaves of Euonymus japonicus, H. Gao and X.L. Fan, 14 May 2021 (BJFC CF20220116, living culture CFCC 55575). Xicheng District, Houhai Park, 116°22′48″ E, 39°56′21″ N, from twigs of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (BJFC CF20220118, living culture CFCC 55578).
Notes: Cesati and De Notaris [71] first introduced the genus Botryosphaeria with 12 species described. However, they did not specify a type species of this genus. Barr et al. [72] suggested that Botryosphaeria dothidea (Basionym: Sphaeria dothidea Moug.: Fries [73]) should be a lectotype of this genus. Slippers et al. [74] re-examined that the host of the holotype of Sphaeria dothidea in the Fries herbarium was Rosa sp., which was not consistent with the description of Fries [73] (on Fraxinus sp.). Additionally, the only other specimen identified as S. dothidea on Fraxinus sp. in the Fries herbarium was immature with no spores [74,75]. This specimen was designated as a neotype [74]. Then, Slippers et al. [74] re-collected specimens from a nearby locality and designated an epitype (PREM 57372) on Prunus sp. collected from Crocifisso, Switzerland, with an ex-epitype culture (CBS 115476 = CMW 8000) with phylogenetic data. Zhang et al. [6] reduced four species to synonymy with Botryosphaeria dothidea based on the high sequence similarity values in ITS region. In this study, twenty-one isolates clustered together with B. dothidea in ML and BI trees. Therefore, they are identified as Botryosphaeria dothidea.
Dothiorella Sacc., Michelia 2 (6): 5, 1880/
Dothiorella acericola Phookamsak, Tennakoon and K.D. Hyde, Fungal Divers. 95: 78, 2019/
Descriptions: See Phookamsak et al. [76].
Material examined: China. Beijing City, Chaoyang District, Sun Palace South Street, 116°26′41″ E, 39°58′29″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220164, living culture CFCC 55559). Haidian District, Beijing Forestry University, 116°20′31″ E, 40°0′16″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220165, living culture CFCC 55561). Mentougou District, Shitan Road, 116°6′31″ E, 39°55′36″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220162, living culture CFCC 55556; BJFC CF20220163, living culture CFCC 55558).
Notes: Dothiorella acericola was first discovered on dead hanging twigs of Acer palmatum Thunb. in Yunnan Province, China [76]. In this study, four isolates (CFCC 55556–55558, and 55561) were grouped together with the ex-type of Do. acericola, KUMCC 18-0137, with a high statistical support (ML/BI = 90/1). Therefore, they are identified as Dothiorella acericola.
Sodariomycetes O.E. Erikss. and Winka, Myconet 1 (1): 10, 1997.
Glomerellales Chadef. ex Réblová, W. Gams and Seifert, Stud. Mycol. 68: 170, 2011.
Colletotrichum Corda, in Sturm, Deutschl. Fl., 3 Abt. (Pilze Deutschl.) 3 (12): 41, 1831.
Colletotrichum aenigma B.S. Weir and P.R. Johnst., in Weir, Johnston, and Damm, Stud. Mycol. 73: 135, 2012.
Descriptions: See Weir et al. [77].
Material examined: China. Beijing City, Daxing District, Daxing New Town Riverfront Forest Park, 116°17′40″ E, 39°44′31″ N, from leaf spots of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220154, living culture CFCC 55535).
Notes: Colletotrichum aenigma was first discovered on Persea americana [77]. Over 20 host species of Co. aenigma were recorded (https://nt.ars-grin.gov/fungaldatabases/index.cfm, accessed on 16 January 2023). In this study, one isolate CFCC 55535 clustered into a clade with Co. aenigma ICMP 18608 with high statistical support (ML/BI = 93/0.99). Therefore, CFCC 55535 is identified as Colletotrichum aenigma. The host range of Co. aenigma also expanded to include Euonymus japonicus.
Colletotrichum euonymi L. Lin and X.L. Fan sp. nov. (Figure 6).
MycoBank: MB 846880
Etymology: The name reflects the host genus from it was collected, Euonymus.
Description: Colonies on PDA reaching 75–80 mm diam after 7 d, flat, white to smoke grey, reverse honey in the center, and white towards the margin. Conidiomata 541–1193 µm, formed on the surface of PDA, covered by olivaceous buff mycelium. Conidiophores formed on inoculated Euonymus japonicus, hyaline, septate, unbranched, 24.5–47.5 × 2.5–4.0 µm (av. = 36.15 ± 5.17 × 3.46 ± 0.41 µm, n = 30). Conidiogenous cells hyaline, smooth-walled, cylindrical to subcylindrical, variable in size, 8.0–18.0 × 2.5–4.0 µm (av. = 13.46 ± 2.66 × 3.28 ± 0.32 µm, n = 30). Conidia hyaline, aseptate, smooth-walled, cylindrical to subcylindrical, guttulate, straight, 14.0–22.5 × 4.5–6.0 µm (av. = 16.67 ± 1.83 × 5.35 ± 0.42 µm, n = 50), L/W ratio = 3.13. Appressoria and Setae not observed.
Typus: China. Beijing City, Shijingshan District, Sculpture Garden Middle Street, 116°14′14″ E, 39°54′18″ N, from leaf spot of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (holotype BJFC CF20220153, ex-holotype culture CFCC 55542). Haidian District, Beijing Forestry University, 116°20′28″ E, 40°0′8″ N, from leaf spot of Euonymus japonicus, H. Gao and X.L. Fan, 21 May 2021 (paratype BJFC CF20220152, ex-paratype culture CFCC 55483).
Additional materials examined: China. Beijing City, Chaoyang District, Olympic Forest Park, 116°23′13″ E, 40°0′4″ N, from leaf spot of Euonymus japonicus, H. Gao and X.L. Fan, 21 May 2021 (BJFC CF20220150, living culture CFCC 55540). Fengtai District, Lotus Pond Park, 116°18′17″ E, 39°53″ N, from leaf spot of Euonymus japonicus, H. Gao and X.L. Fan, 21 May 2021 (BJFC CF20220151, living culture CFCC 55537).
Notes: In this study, four isolates of Colletotrichum euonymi formed a distinct clade with high statistical support (ML/BI = 100/1). In Colletotrichum gloeosporioides complex, Co. gloeosporioides and Co. siamense have been recorded to host Euonymus [78,79]. However, Co. euonymi can be distinguished from them by larger conidia (14.0–22.5 × 4.5–6.0 µm vs. 12.0–17 × 4.5–6.0 µm for Co. gloeosporioides and 7–18.3 × 3–4.3 µm for Co. siamense) [78,79].
Colletotrichum euonymicola L. Lin and X.L. Fan sp. nov. (Figure 7).
MycoBank: MB 846881
Etymology: The name reflects that the species is a Euonymus-colonizer.
Description: Colonies on PDA 73–82 mm diam in 7 d, flat with undulate edge, mouse grey to dark mouse grey, aerial mycelium short, Conidiomata not developed. Conidiophores formed directly from hyphae. Conidiophores hyaline to brown, septate, branched. Conidiogenous cells hyaline, smooth-walled, cylindrical or slightly tapering towards the apex, 10.5–16.5 × 1.0–3.5 μm. Conidia hyaline, aseptate, smooth-walled, guttulate, cylindrical with obtuse ends, with the base sometimes tapering to a truncate hilum, (9.5) 11.0–23.0 (25.0) × 3.5–6.5 (6.9) µm (av. = 14.3 ± 2.85 × 4.73 ± 0.69 µm, n = 50), L/W ratio = 3.04. Appressoria single, smoke grey to iron grey, terminally at the tip of the hyphae, irregularly shaped, with undulate to lobate margins, 10.0–14.5 × 6.0–8.0 μm. Setae was not observed.
Typus: China. Beijing City, Chaoyang District, 116°23′13″ E, 40°0′4″ N, Sun Palace Middle Street, from leaf spot of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (holotype BJFC CF20220157, ex-holotype culture CFCC 55486; paratype BJFC CF20220158, ex-paratype culture CFCC 55539).
Notes: Colletotrichum euonymicola is phylogenetically related to Co. pseudotheobromicola. However, they show differences of sequence at ITS (13/549), act (4/284), tub2 (1/445), chs-1 (1/296), and gapdh (2/278). Morphologically, Co. euonymicola differs from Co. pseudotheobromicola in that it produces larger-sized appiospora (10.0–14.5 × 6.0–8.0 μm vs. 6–10 × 5–8 μm) [80]. Moreover, Co. euonymicola is different from Co. pseudotheobromicola in terms of host plant species (Euonymus japonicus v.s. Prunus avium). Therefore, Colletotrichum euonymicola is introduced as a novel species here.
Colletotrichum gloeosporioides (Penz.) Penz. and Sacc., Atti Inst. Veneto Sci. lett., ed Arti, Sér. 6 2 (5): 670, 1884.
Descriptions: See Cannon et al. [78].
Materials examined: China. Beijing City, Xicheng District, Houhai Park, 116°22′34″ E, 39°56′25″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (BJFC CF20220155 living culture CFCC 55544–55545); 116°23′7″ E, 39°56′27″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 7 May 2021 (BJFC CF20220156 living culture CFCC 55547).
Notes: The host range of Colletotrichum gloeosporioides includes multiple families. It has been reported that Co. gloeosporioides can affected Euonymus fortunei and Euonymus japonicus [81,82]. In this study, three isolates grouped together with Co. gloeosporioides IMI 356878 with a high statistical support (ML/BI = 95/1). Therefore, they are identified as Colletotrichum gloeosporioides based on the phylogenetic tree.
Diaporthales Nannf., Nova Acta R. Soc. Scient. Upsal., Ser. 48 (2): 53, 1932.
Cytospora Ehrenb., Sylv. mycol. berol. (Berlin): 28, 1818.
Cytospora ailanthicola X.L. Fan and C.M. Tian, Persoonia 45: 13, 2020.
Descriptions: See Fan et al. [32].
Material examined: China. Beijing City, Chaoyang District, Olympic Forest Park, 116°23′9″ E, 40°0′2″ N, from branches of Euonymus japonicus, X.W. Zhu, 28 April 2020 (BJFC CF20220120, living culture CFCC 55529).
Notes: Cytospora ailanthicola was first introduced on branches of Ailanthus altissima [32]. Lin et al. [52] confirmed this species was a pathogen with strong virulence caused by poplar canker disease. In this study, one isolate, CFCC 55529, was isolated from symptomatic branches of Euonymus japonicus in Beijing, which clustered in a well-supported clade with C. ailanthicola ex-holotype CFCC 89970 (ML/BI = 100/1). Therefore, CFCC 55529 is identified as Cytospora ailanthicola.
Cytospora albodisca M. Pan and X.L. Fan, Front. Plant Sci. 12 (636460): 3, 2021.
Descriptions: See Pan et al. [83].
Material examined: China. Beijing City, Mengtougou District, Baihuashan Nature Reserve, 115°33′15″ E, 39°51′52″ N, from branches of Euonymus japonicus, M. Pan and X.L. Fan, 21 August 2021 (BJFC CF20220121, living culture CFCC 56274, 57538).
Notes: Cytospora albodisca was first discovered on Platycladus orientalis, whose ascostroma was surrounded by a black conceptacle [83]. In this study, two isolates (CFCC 56274 and 57538) converged into a separate little branch. However, they only differ from C. albodisca CFCC 53161 and 54373 by 1/778 in act gene and 1/732 in tub2 gene (with gaps). Additionally, the isolates in this study (CFCC 56274 and 57538) grouped together with C. albodisca CFCC 53161 and 54373 with a high statistical support (ML/BI = 94/1). Therefore, these two isolates are identified as Cytospora albodisca.
Cytospora discostoma M. Pan and X.L. Fan, Front. Plant Sci. 12 (636460): 3, 2021.
Descriptions: See Pan et al. [83].
Material examined: China. Beijing City, Mengtougou District, Baihuashan Nature Reserve, 115°33′17″ E, 39°52′52″ N, from branches of Euonymus japonicus, M. Pan and X.L. Fan, 21 August 2021 (BJFC CF20220122, living culture CFCC 56276).
Notes: Cytospora discostoma was first discovered on branches of Platycladus orientalis at Mentougou District in Beijing [83]. In this study, one isolate, CFCC 56276, clustered in a well-support clade (ML/BI = 100/1) with C. discostoma CFCC 53137 and 54368. The specimen BJFC CF20220122 in this study was collected from branches of Euonymus japonicus at Mentougou District in Beijing, where Cytospora discostoma was first discovered.
Cytospora diopuiensis Q.J. Shang, J.K. Liu and K.D. Hyde, Mycosphere, 11 (1): 202, 2020.
Descriptions: See Shang et al. [84].
Material examined: China. Beijing City, Haidian District, Beijing Forestry University, 116°20′28″ E, 40°0′8″ N, from leaves of Euonymus japonicus, H. Gao and X.W. Zhu, 21 October 2020 (BJFC CF20220146, living culture CFCC 54692; BJFC CF20220147, living culture CFCC 55479). Shijingshan District, Sculpture Garden Middle Street, 116°14′11″ E, 39°54′10″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 20 April 2021 (BJFC CF20220148, living culture CFCC 55527; BJFC CF20220149, living culture CFCC 55528).
Notes: Cytospora diopuiensis was discovered on bark of dead wood in Thailand [84]. Jiang et al. [85] reported this species on Kerria japonica f. pleniflora in China. In this study, two isolates from leaves of Euonymus japonicus and two isolates from branches clustered in a well-supported clade with C. diopuiensis (ML/BI = 100/1). Therefore, they were identified as Cytospora diopuiensis.
Cytospora elaeagni Allesch., Hedwigia 36: 162, 1897.
Descriptions: See Fan et al. [86].
Material examined: China. Beijing City, Shijingshan District, Beijing International Sculpture Park, 116°14′11″ E, 39°54′10″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 23 April 2021 (BJFC CF20220123, living culture CFCC 54082). Haidian District, Beijing Forestry University, 116°20′27″ E, 40°0′13″ N, from leaves of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220124, living culture CFCC 55477). The Shucun Park, 116°17′55″ E, 40°0′53″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220125, living culture CFCC 55526). Mengtougou District, Baihuashan Nature Reserve, 115°33′25″ E, 39°51′53″ N, from branches of Euonymus japonicus, M. Pan, Y.K. Bai and X.L. Fan, 21 August 2021 (BJFC CF20220126, living culture CFCC 56273). 115°34′15″ E, 39°51′56″ N, from branches of Euonymus japonicus, M. Pan, Y.K. Bai and X.L. Fan, 21 August 2021 (BJFC CF20220127, living culture CFCC 56287).
Notes: Cytospora elaeagni has been reported from Elaeagnus angustifolia in China, German, and the USA [87,88,89]. Fan et al. [86] provided its morphological descriptions and molecular data. In this study, five isolates are identified as Cytospora elaeagni based on phylogenetic analyses. A new host record from Euonymus japonicus is provided here.
Cytospora euonymicola X.L. Fan and C.M. Tian, Persoonia 45: 13, 2020.
Descriptions: See Fan et al. [32].
Material examined: China. Beijing City, Haidian District, Beijing Forestry University, 116°20′31″ E, 40°0′16″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220128, living culture CFCC 54688). Fengtai District, Lotus Pond Park, 116°18′17″ E, 39°53′34″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220129, living culture CFCC 55530).
Notes: Cytospora euonymicola was first introduced on Euonymus kiautschovicus in Shaanxi Province, China [32]. In this study, two isolates grouped together with C. euonymicola in ML and BI trees (ML/BI = 100/1). Morphologically, the conidia size in this study were similar in C. euonymicola described by Fan et al. [32] (4.5–5.0 × 1.0–1.5 μm vs. 4–5 × 1 μm). Therefore, the two isolates in the current study are identified as Cytospora euonymicola based on phylogeny and morphology.
Cytospora euonymina X.L. Fan and C.M. Tian, Persoonia 45: 13, 2020.
Descriptions: See Fan et al. [32].
Material examined: China. Beijing City, Daxing District, Nanchengzhuang, 116°17′39″ E, 39°44′26″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 11 April 2021 (BJFC CF20220130, living culture CFCC 55524). Xicheng District, Houhai Park, 116°22′49″ E, 39°56′21″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220131, living culture CFCC 55525).
Notes: Cytospora euonymina was first introduced on Euonymus kiautschovicus in Shanxi Province, China [32]. In this study, two isolates grouped together with C. euonymina in ML and BI trees (ML/BI = 100/1). Therefore, they were identified as C. euonymina. Additionally, CFCC 55524 and CFCC 55525 were isolated from leaves of Euonymus japonicus in the current study. The discs of conidioma formed on leaves (100–150 µm in this study) were smaller than the description of Fan et al. (2020) (200–230 µm, on branches).
Cytospora haidianensis X. Zhou and X.L. Fan, Forests, 11 (5): 524, 2020.
Descriptions: See Zhou et al. [33].
Material examined: China. Beijing City, Chaoyang District, Olympic Forest Park, 116°23′9″ E, 40°0′2″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220132, living culture CFCC 55480; BJFC CF20220134, living culture CFCC 55532). 116°23′10″ E, 40°0′2″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220135, living culture CFCC 55533). Shijingshan District, Sculpture Garden Middle Street, 116°14′14″ E, 39°54′18″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220133, living culture CFCC 55531).
Notes: Cytospora haidianensis was first introduced as a pathogen on Euonymus alatus by Zhou et al. [33]. This species has a toruloid locule with a central column of ostiolar tissue [33]. In this study, four isolates grouped together with C. haidianensis in ML and BI trees (ML/BI = 100/1). Therefore, they are identified as Cytospora haidianensis.
Cytospora leucostoma (Pers.) Sacc., Michelia, 2 (7): 264, 1881.
Descriptions: See Fan et al. [32].
Material examined: China. Beijing City, Haidian District, Beijing Forestry University, 116°20′27″ E, 40°0′13″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220136, living culture CFCC 55474). 116°20′32″ E, 40°0′13″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220137, living culture CFCC 55475). 116°20′31″ E, 40°0′16″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220138, living culture CFCC 55476). 116°20′28″ E, 40°0′17″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220139, living culture CFCC 55478). Chaoyang District, Olympic Forest Park, 116°23′10″ E, 40°0′2″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220140, living culture CFCC 55519; BJFC CF20220142, living culture CFCC 55521). Mentougou District, The Shitan Road, 116°6′31″ E, 39°55′36″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220141, living culture CFCC 55520).
Notes: Fan et al. [32] revealed C. donetzica as synonym of C. leucostoma based on DNA data and descriptions. In this study, seven isolates grouped together with C. leucostoma in ML and BI trees (ML/BI = 100/1). Morphologically, the size of conidia in this study was 5.0–6.0 × 1.0–2.0 µm, which overlapped with the description by Fan et al. [32] (4.5–5.5 × 1–1.5 μm). Therefore, these seven isolates are identified as Cytospora leucostoma.
Cytospora sophorae Bres., Fung. trident. 2 (8–10): 44, 1892.
Descriptions: See Fan et al. [90].
Material examined: China. Beijing City, Mentougou District, Lisichen Park, 116°6′31″ E, 39°55′37″ N, from leaves spots of Euonymus japonicus, H. Gao and X.L. Fan, 11 May 2021 (BJFC CF20220143, living culture CFCC 55523).
Notes: Cytospora sophorae has been recorded as a pathogen caused Sophora canker disease in China [89,91]. Fan et al. [90] provided the description and DNA data of this species. In this study, one isolate CFCC 55523 grouped together with C. sophorae in ML and BI trees (ML/BI = 100/1). Therefore, it is identified as Cytospora sophorae.
Cytospora zhaitangensis L. Lin and X.L. Fan sp. nov. (Figure 8).
MycoBank: MB 846882
Etymology: The name reflects the station where the holotype and paratype specimens were collected, next to the Zhaitang reservoir.
Description: Asexual morph: Conidiomata cytosporoid rosette, immersed in bark, erumpent when mature, discoid to conical, 524–936 µm in diam, with multiple locules. Conceptacles were absent. Disc hazel to olivaceous, circular to ovoid, 166–224 µm in diam, with a single ostiole per disc in the center. Ostiole circular to ovoid, olivaceous buff to dark mouse grey, at the same level or above as disc surface, 66–98 µm in diam. Locules are complex with irregular shapes, and do not share common walls. Conidiophores hyaline, unbranched, or branched at the bases or at mid-height, 13.5–27.0 (30.0) × 1.0–2.0 µm (av. = 19.71 ± 4.51 × 1.40 ± 0.18 µm, n = 30). Conidiogenous cells enteroblastic, phialidic, subcylindrical to cylindrical, 6.0–11.0 × 1.0–2.0 µm (av. = 8.05 ± 1.29 × 1.37 ± 0.18 µm, n = 30). Conidia hyaline, unicellular, eguttulate, elongate-allantoid, (3.8) 4.0–6.5 × (1.2) 1.3–1.6 (1.7) µm (av. = 5.14 ± 0.74 × 1.47 ± 0.12 µm, n = 50), L/W ratio = 3.48. Sexual morph: not observed.
Typus: China. Beijing City, Mengtougou District, next to the Zhaitang Reservoir, 115°33′15″ E, 39°51′52″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 August 2021 (holotype BJFC CF20220144, ex-holotype culture CFCC 56227). 115°33′16″ E, 39°51′54″ N, from branches of Euonymus japonicus, M. Pan, Y.K. Bai and X.L. Fan, 21 August 2021 (paratype BJFC CF20220145, ex-paratype culture CFCC 57537).
Notes: Cytospora zhaitangensis is phylogenetically most closely related to C. euonymicola and C. gigalocus. Morphologically, C. zhaitangensis can be differentiated by the wider conidia from C. euonymicola (L/W ratio = 3.48 vs. L/W ratio = 4.5) and C. gigalocus (L/W ratio = 4.36) [32,86]. Additionally, C. zhaitangensis has smaller discs (166–224 µm) than C. euonymicola (240–350 µm) and C. gigalocus (330–620 µm) [32,86]. Cytospora euonymi and C. euonymella were also recorded to host Euonymus [92,93]. Cytospora zhaitangensis can be differentiated by the conidia size from C. euonymella (4.0–6.5 × 1.3–1.6 vs. 2.5 × 0.5 µm) and C. euonymi (8 × 2 µm) [92,93].
Diaporthe Nitschke, Pyrenomyc. Germ. 2: 240, 1870.
Diaporthe eres Nitschke, Pyrenomyc. Germ. 2: 240, 1870.
Descriptions: See Gao et al. [26].
Material examined: China. Beijing City, Chaoyang District, Olympic Forest Park, 116°23′9″ E, 40°0′2″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220159, living culture CFCC 55481). 116°26′52″ E, 39°58′5″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220160, living culture CFCC 55482). Shijingshan District, Sculpture Garden Middle Street, 116°14′14″ E, 39°54′18″ N, from branches of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (BJFC CF20220161, living culture CFCC 55534).
Notes: Norphanphoun et al. [94] introduced 13 species complexes of Diaporthe, with revealing D. eres species complex introduced by Udayanga et al. [95] as D. alnea species complex according to nomenclatural articles. Diaporthe eres strains collected from different hosts were dispersed in the clade [94,95]. In this study, three isolates were dispersed in Diaporthe alnea species complex. To avoid over-classification, we only identified them as Diaporthe eres.
Amphisphaeriales D. Hawksw. and O.E. Erikss., Syst. Ascom. 5 (1): 177, 1986.
Pestalotiopsis Steyaert, Bull. Jard. Bot. État Brux. 19: 300, 1949.
Pestalotiopsis chaoyangensis L. Lin and X.L. Fan sp. Nov. (Figure 9).
MycoBank: MB 846883
Etymology: The name reflects the station where the holotype and paratype specimens were collected, Chaoyang District, Beijing, China.
Description: On PDA, conidiomata was not observed. Conidial masses abundant, black, scattered, or confluent, formed among the mycelia. Conidiophores are indistinct, and are usually reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to subcylindrical. Conidia fusoid, straight or slightly curved, 4-septate, smooth, slightly constricted at the septa, 19.5–25.5 × 4.5–6.5 µm (av. = 22.34 ± 1.17 × 5.52 ± 0.38 µm, n = 50), L/W ratio = 4.05; basal cell obconic with a truncate base, thin-walled, hyaline or pale brown, 3.5–5.5 µm (av. = 4.52 ± 0.44 µm, n = 50); median cells 3, trapezoid or subcylindrical, concolorous, pale brown to brown, thick-walled, the first median cell from base 3.5–6.0 µm (av. = 4.36 ± 0.52 µm, n = 50) mm long, the second cell 4.0–6.5 µm (av. = 4.88 ± 0.39 µm, n = 50) long, the third cell 3.5–5.5 µm (av. = 4.73 ± 0.43 µm, n = 50) long; apical cell conic with an acute apex, thin-walled, hyaline, 2.0–4.5 µm (av. = 3.44 ± 0.43 µm, n = 50) long; basal appendage single, occasionally 2, tubular, centric, straight or slightly bent, 2.0–6.5 µm (av. = 4.02 ± 1.14 µm, n = 50) long; apical appendages 2, unbranched, tubular, centric, straight or bent, 14.0–22.5 µm (av. = 17.56 ± 2.38 µm, n = 50). Sexual morph is unknown.
Typus: China. Beijing City, Chaoyang District, Olympic Forest Park, 116°23′10″ E, 40°0′2″ N, from leaves spots of Euonymus japonicus, H. Gao and X.L. Fan, 21 April 2021 (holotype BJFC CF20220167, ex-holotype culture CFCC 55549; paratype BJFC CF20220168, ex-paratype culture CFCC 58805).
Notes: Two isolates of Pestalotiopsis chaoyangensis (CFCC 55549 and 58805) formed a distinct clade phylogenetically close to P. shaanxiensis (Figure 5). However, P. chaoyangensis can be distinguished from P. shaanxiensis by the number of apical appendages with two for the former and three for the latter [39].
3.4. Pathogenicity Test
In the leaf inoculation assays, fourteen days after inoculation, leaf lesions were caused by all three species isolated from leaves (Co. euonymi CFCC 55542, Co. euonymicola CFCC 55486, and P. chaoyangensis CFCC 55549) (Figure 10 and Figure 11, Table 4). Disease sites initially turned yellow, and as disease progressed, diseased patches enlarged and took on a water-stained appearance, ultimately leading to wilting and consequent death. No symptoms were observed in the non-inoculated controls. All pathogenic species were re-isolated from lesions or conidia masses of inoculated leaves.
In the branch inoculation assay, no symptoms were observed with C. zhaitangensis inoculation or in the non-inoculated controls.
4. Discussion
Euonymus japonicus is an evergreen shrub that often becomes seriously diseased and even dies from fungal infestations in Beijing, China. In the current study, 79 isolates were obtained from 104 specimens collected from seven districts in Beijing City. The isolates included 22 species in seven genera, which were Aplosporella (eight isolates, three species), Botryosphaeria (21 isolates, one species), Colletotrichum (10 isolates, four species), Cytospora (31 isolates, 11 species), Diaporthe (three isolates, one species), Dothiorella (four isolates, one species), and Pestalotiopsis (two isolates, one species). Among the 22 species, Co. euonymi, Co. euonymicola, C. zhaitangensis, and P. chaoyangensis were identified as novel species on the basis of morphological and phylogenetic analyses. Colletotrichum euonymi, Co. euonymicola, and P. chaoyangensis were confirmed as pathogens on leaves of E. japonicus. In this study, A. hesperidica, A. javeedii, A. prunicola, Co. aenigma, C. ailanthicola, C. albodisca, C. diopuiensis, C. discotoma, C. elaeagni, C. sophorae, and Do. acericola were first recorded on the host genus Euonymus.
Cytospora had the highest diversity of species associated with E. japonicus (11 species). The genus includes numerous important pathogens and saprophytic fungi on various hosts [32,33,83,96]. Branch and stem diseases frequently result in skin rot, dryness, and plant death [51,52,57,97]. Among the 10 known species obtained from E. japonicus in the current study, C. ailanthicola and C. haidianensis are confirmed pathogens on Populus and Euonymus, respectively [32,52]. However, C. zhaitangensis, the novel species isolated from branches, was not pathogenic on E. japonicus. The pathogenicity of other Cytospora species on Euonymus needs to be studied further.
Botryosphaeria dothidea was the species with the highest number of isolates (21 isolates), which were distributed in Chaoyang, Daxing, Haidian, Mentougou, Shijingshan, and Xicheng districts. Chaoyang District had the most fungal species occurring on E. japonicus (11 species in seven genera), followed by Haidian and Mentougou districts (nine species in five and four genera, respectively). The differences among districts could be because Chaoyang District is a major industrial district in Beijing, which has more environmental pollution. Schmidt et al. [98] concluded that Sordariomycetous fungi dominated at the polluted site and species diversity of endophytes was higher at the unpolluted site. These changes could weaken plants and increase susceptibility to disease.
Multiple infections can occur on different plant parts. For example, B. dothidea causes apple ring rot of stems, twigs, and fruits [99], and Co. gloeosporioides causes anthracnose of leaves and fruits [100]. Among the seven genera in this study, Aplosporella, Botryosphaeria, Cytospora, Dothiorella, and Diaporthe are pathogenic and cause canker and dieback disease of various hosts [6,29,32,64,69,94,96]. However, eight isolates were obtained from leaves in the current study, i.e., B. dothidea CFCC 55575, C. diopuiensis CFCC 54692 and 55479, C. elaeagni CFCC 55477, C. euonymina CFCC 55524 and 55525, C. sophorae CFCC 55523, and Do. acericola CFCC 55559. The isolates also created reproductive structures on leaves, which indicated that the species may be able to infect branches as well as leaves. The pathogenicity of these species to leaves and stems needs to be studied further. Increased understanding of pathogen diversity is beneficial to E. japonicus production and maintenance because impacts of disease can be minimized, and disease management needs to be improved.
Over 90 species of fungi occurring on E. japonicus are recorded in the Fungal database (https://nt.ars-grin.gov/fungaldatabases/index.cfm; accessed on 16 January 2022). There are numerous cases of Erysiphe and Phytophthora infesting Euonymus in China, in addition to the taxa identified in the current study [88,89,101,102]. Erysiphe alphitoides, E. euonymi, E. euonymicola, E. lianyungangensis, E. mayumi, and E. pseudopusilla have been recorded on host Euonymus [102,103,104,105]. Phytophthora citrophthora, P. meadii, and P. palmivora have been reported on host E. japonicus [88,89,101]. Therefore, the two genera also need to be considered in disease control.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof9020271/s1, Figure S1. Phylogram of Aplosporella based on maximum likelihood (ML) analysis of the dataset of combined ITS and tef1-α genes; Figure S2. Phylogram of Botryosphaeria based on maximum likelihood (ML) analysis of the dataset of combined ITS, tef1-α, and tub2 genes; Figure S3. Phylogram of Colletotrichum based on maximum likelihood (ML) analysis of the dataset of ITS gene; Figure S4. Phylogram of Diaporthe based on maximum likelihood (ML) analysis of the dataset of combined ITS, cal, his3, tef1-α, and tub2 genes; Figure S5. Phylogram of Dothiorella based on maximum likelihood (ML) analysis of the dataset of combined ITS, tef1-α, and tub2 genes; Table S1: Strains used in the molecular analyses in this study.
Author Contributions
Conceptualization, L.L. and X.F.; methodology, L.L. and M.P.; software, L.L. and X.F.; validation, L.L. and M.P.; formal analysis, L.L. and M.P.; investigation, H.G., M.P. and X.F.; resources, L.L., X.F. and C.T.; writing—original draft preparation, L.L.; writing—review and editing, H.G., M.P. and X.F.; visualization, L.L.; supervision, X.F.; funding acquisition, X.F. and C.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32101533), National Science and Technology Fundamental Resources Investigation Program of China (2021FY100900).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Alignments generated during the current study are available in Tree-BASE (accession http://purl.org/phylo/treebase/phylows/study/TB2:S29991 (accessed on 15 February 2023)). All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Acknowledgments
We are grateful to Yingmei Liang (the Museum of Beijing Forestry University) for providing relevant specimens, also to Xiaohong Liang, Jing Han (the Experimental Teaching Centre, College of Forestry, Beijing Forestry University) for providing installed scientific equipment in the whole process. And we are grateful for the assistance of Xinwei Zhu, Yukun Bai, Haoyu Zhou, and Zhihui Bian (Beijing Forestry University) during this study. Xinlei Fan would like to acknowledge the supporting of strain preservation of Chungen Piao and Minwei Guo (China Forestry Culture Collection Centre, Chinese Academy of Forestry, Beijing).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Bai, Y.; Hong, X.; Sun, L.; Liu, Y. Particulate matter and heavy metal deposition on the leaves of Euonymus japonicus during the East Asian monsoon in Beijing, China. PLoS ONE 2017, 12, e0179840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.W.; Yang, L.F.; Ding, Z.Q.; Tu, J.H. Characteristics of absorption and enrichment of heavy metal in Photinia serrulata and Euonymus japonicus planted in sewage sludge substrates. North. Hortic. 2010, 3, 70–74. (In Chinese) [Google Scholar]
- Raza, M.; Zhang, Z.F.; Hyde, K.D.; Diao, Y.Z.; Cai, L. Culturable plant pathogenic fungi associated with sugarcane in southern China. Fungal Divers. 2019, 99, 1–104. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Edwards, J.; Taylor, P.W.J.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Pers.-Mol. Phylogeny Evol. Fungi 2021, 46, 63–115. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Phillips, A.J.L.; Li, X.H.; Hyde, K.D. Botryosphaeriaceae, Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef]
- Phillips, A.J.; Hyde, K.D.; Alves, A.; Liu, J.K. Families in Botryosphaeriales, a phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae, genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Cash, E.K. A Record of the Fungi Named by JB Ellis; Division of Mycology and Disease Survey, Bureau of Plant Industry, Soils, and Agricultural Engineering, Agricultural Research Administration, US Department of Agriculture: Washington, DC, USA, 1952.
- Hanlin, R.T. A Revision of the Ascomycetes of Georgia; Georgia Agricultural Experiment Stations, University of Georgia, College of Agriculture: Athens, GA, USA, 1963. [Google Scholar]
- Reid, D. An annotated list of some fungi from the Channel Islands, mostly from Jersey. Trans. Br. Mycol. Soc. 1985, 84, 709–714. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Phillips, A.J.L.; Fu, C.Y.; Yan, J.Y.; Li, X.H. Dothiorella species associated with woody hosts in Italy. Mycosphere 2016, 7, 51–63. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Liu, F.; Ma, Z.Y.; Hou, L.W.; Diao, Y.Z.; Wu, W.P.; Damm, U.; Song, S.; Cai, L. Updating species diversity of Colletotrichum, with a phylogenomic overview. Stud. Mycol. 2022, 101, 1–86. [Google Scholar] [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [Green Version]
- Blain, W.L. A list of diseases of economic plants in Alabama. Mycologia 1931, 23, 300–304. [Google Scholar] [CrossRef]
- Lee, H.B.; Park, J.Y.; Jung, H.S. First report of leaf anthracnose caused by Colletotrichum boninense on spindle trees. Plant Pathol. 2005, 54, 254. [Google Scholar] [CrossRef]
- Alizadeh, A.; Javan-Nikkhah, M.; Zare, R.; Fotouhifar, K.B.; Damm, U.; Stukenbrock, E.H. New records of Colletotrichum species for the mycobiota of Iran. Mycol. Iran. 2015, 2, 95–109. [Google Scholar]
- Wang, T.T.; Li, P.L.; Li, H.W.; Li, J.; Gao, F.; Liu, D.; Yan, J.M.; Gong, G.S. First report of Colletotrichum gloeosporioides sensu stricto causing anthracnose on Nandina domestica in Sichuan Province of China. Plant Dis. 2018, 102, 822. [Google Scholar] [CrossRef]
- Wu, M. First report of Colletotrichum siamense causing Anthracnose on Euonymus japonicus in China. Plant Dis. 2020, 104, 587. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.; Jeewon, R.; Phillips, A.J.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, N.; Tian, C.M. Three new Diaporthe species from Shaanxi province, China. MycoKeys 2020, 67, 1–18. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, N.; Tian, C.M. New species and records of Diaporthe from Jiangxi Province, China. MycoKeys 2021, 77, 41–64. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Pan, M.; Tian, C.M.; Fan, X.L. Cytospora and Diaporthe species associated with hazelnut canker and dieback in Beijing, China. Front. Cell. Infect. Microbiol. 2021, 11, 664366. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from Fagaceae leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Piao, C.G.; Li, Y. Two new species of Diaporthe (Diaporthaceae, Diaporthales) associated with tree cankers in The Netherlands. MycoKeys 2021, 85, 31. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.K.; Pan, M.; Gao, H.; Lin, L.; Tian, C.M.; Fan, X.L. Studies of Diaporthe species causing hazelnut canker disease in Beijing, China, with two new species described. Plant Pathol. 2022, 71, 1980–1991. [Google Scholar] [CrossRef]
- Simonyan, S.A. Mycoflora of Botanical Gardens and Arboretums of the Armenian SSR (Translated from Russian); Publishing House of the Academy of Sciences of the Armenian SSR: Yerevan, Armenia, 1981. [Google Scholar]
- Ahmad, S.; Iqbal, S.H.; Khalid, A.N. Fungi of Pakistan; Sultan Ahmad Mycological Society of Pakistan: Lahore, Pakistan, 1997. [Google Scholar]
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Pers.-Mol. Phylogeny Evol. Fungi 2020, 45, 1–45. [Google Scholar] [CrossRef]
- Zhou, X.; Pan, M.; Li, H.; Tian, C.M.; Fan, X.L. Dieback of Euonymus alatus (Celastraceae) Caused by Cytospora haidianensis sp. nov. in China. Forests 2020, 11, 524. [Google Scholar] [CrossRef]
- Wehmeyer, L.E. The Genus Diaporthe Nitschke and Its Segregates; University of Michigan Press: Ann Arbor, MI, USA, 1933. [Google Scholar]
- Otani, Y. Mycological Flora of Japan; Yokendo Ltd.: Tokyo, Japan, 1995. [Google Scholar]
- Mulenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A preliminary checklist of Micromycetes in Poland. W. Szafer Institute of Botany. Pol. Acad. Sci. 2008, 9, 752. [Google Scholar]
- Gajanayake, A.J.; Abeywickrama, P.D.; Jayawardena, R.S.; Camporesi, E.; Bundhun, D. Pathogenic Diaporthe from Italy and the first report of D. foeniculina associated with Chenopodium sp. Plant Pathol. Quar. 2020, 10, 172–197. [Google Scholar] [CrossRef]
- Steyaert, R.L. Contribution à l’étude monographique de Pestalotia de Not. et Monochaetia Sacc. (Truncatella gen. nov. et Pestalotiopsis gen. nov.). Bull. Jard. Bot. Brux. 1949, 19, 285–354. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Xue, H.; Piao, C.G.; Li, Y. Morphology and phylogeny of Pestalotiopsis (Sporocadaceae, Amphisphaeriales) from Fagaceae leaves in China. Microbiol. Spectr. 2022, 10, e03272-22. [Google Scholar] [CrossRef] [PubMed]
- Nag Raj, T.R. Coelomycetous Anamorphs with Appendage-Bearing Conidia; Mycologue Publications: Waterloo, ON, Canada, 1993. [Google Scholar]
- Kobayashi, T.; Watanabe, K.; Ono, Y.; Furukawa, T. Index of fungi inhabiting woody plants in Japan-host, distribution and literature. Mycoscience 2010, 51, 379–386. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Guo, L.D.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167–187. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Crous, P.W.; Bhat, D.J.; McKenzie, E.H.C.; Bahkali, A.H.; et al. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes, evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerber, J.C.; Liu, B.; Correll, J.C.; Johnston, P.R. Characterization of diversity in Colletotrichum acutatum sensu lato by sequence analysis of two gene introns, mtDNA and intron RFLPs, and mating compatibility. Mycologia 2003, 95, 872–895. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Groenewald, J.Z.; Risède, J.M.; Simoneau, P.; Hywel-Jones, N.L. Calonectria species and their Cylindrocladium anamorphs, species with sphaeropedunculate vesicles. Stud. Mycol. 2004, 50, 415–430. [Google Scholar]
- Lin, L.; Pan, M.; Tian, C.M.; Fan, X.L. Fungal Richness of Cytospora species associated with willow canker disease in China. J. Fungi 2022, 8, 377. [Google Scholar] [CrossRef]
- Lin, L.; Pan, M.; Bezerra, J.; Tian, C.M.; Fan, X.L. Re-evaluation of the fungal diversity and pathogenicity of Cytospora species from Populus in China. Plant Dis. 2022; in press. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA 6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, H.O. New algorithms and methods to estimate maximum-likelihood phylogenies, assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3, Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nouri, M.T.; Lawrence, D.P.; Holland, L.A.; Doll, D.A.; Kallsen, C.E.; Culumber, C.M.; Nylander, J.A.A. MrModeltest v2. Distributed by the Author; Evolutionary Biology Center, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A.; Drummond, A. FigTree v. 1.3.1. Institute of Evolutionary Biology; University of Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: Kew, UK, 1970. [Google Scholar]
- Wickham, H. ggplot2, Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. 2018. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 15 February 2023).
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.; Jones, E.B.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Milan, C.; Samarakoon, M.C.; Malaithong, M.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Spegazzini, C. Fungi argentini additis nonnullis brasiliensibus montevideensibusque. Anal. Soc. Cient. Argent. 1882, 12, 97–117. [Google Scholar]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Botryosphaeriaceae species overlap on four unrelated, native South African hosts. Fungal Biol. 2014, 118, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Yang, Q.; Cao, B.; Liang, Y.M.; Tian, C.M. New records of Aplosporella javeedii on five hosts in China based on multi-gene analysis and morphology. Mycotaxon 2015, 130, 749–756. [Google Scholar] [CrossRef]
- Jami, F.; Wingfield, M.J.; Gryzenhout, M.; Slippers, B. Diversity of tree-infecting Botryosphaeriales on native and non-native trees in South Africa and Namibia. Australas. Plant Pathol. 2017, 46, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Li, J.; Piao, C.G.; Guo, M.W.; Tian, C.M. Identification and characterization of chestnut branch-inhabiting melanocratic fungi in China. Mycosphere 2018, 9, 1268–1289. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Tian, C.M.; Fan, X.L. Studies of botryosphaerialean fungi associated with canker and dieback of tree hosts in Dongling Mountain of China. Phytotaxa 2018, 348, 63–76. [Google Scholar] [CrossRef]
- Pan, M.; Zhu, H.Y.; Bezerra, J.D.P.; Bonthond, G.; Tian, C.M.; Fan, X.L. Botryosphaerialean fungi causing canker and dieback of tree hosts from Mount Yudu in China. Mycol. Prog. 2019, 18, 1341–1361. [Google Scholar] [CrossRef]
- Damm, U.; Fourie, P.H.; Crous, P.W. Aplosporella prunicola, a novel species of anamorphic Botryosphaeriaceae. Fungal Divers. 2007, 27, 35–43. [Google Scholar]
- Cesati, V.; De Notaris, G. Schema di classificazione degle sferiacei italici aschigeri piu’ o meno appartenenti al genere Sphaeria nell’antico significato attribuitoglide Persono. Comment. Della Soc. Crittogamolog. Ital. 1863, 1, 177–420. [Google Scholar]
- Barr, M.E. Preliminary studies on the Dothideales in temperate North America. Mich. Univ. Herb Contrib. 1972, 9, 523–638. [Google Scholar]
- Fries, E.M. Systema Mycologicum; Sumtibus Ernesti Mauritii: Gryphiswald, Germany, 1823; Volume 2. [Google Scholar]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.L.; Lucas, M.T. The taxonomic status of Macrophoma flaccida and Macrophoma reniformis and their relationship to Botryosphaeria dothidea. Sydowia 1997, 49, 150–159. [Google Scholar]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, P.F.; Buddie, A.G.; Bridge, P.D. The typification of Colletotrichum gloeosporioides. Mycotaxon 2008, 104, 189–204. [Google Scholar]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Chethana, K.W.T.; Jayawardene, R.S.; Zhang, W.; Zhou, Y.Y.; Liu, M.; Hyde, K.D.; Li, X.H.; Wang, J.; Zhang, K.C.; Yan, J.Y. Molecular characterization and pathogenicity of fungal taxa associated with cherry leaf spot disease. Mycosphere 2019, 10, 490–530. [Google Scholar] [CrossRef]
- Mahoney, M.J.; Tattar, T.A. Identification, etiology, and control of Euonymus fortunei anthracnose caused by Colletotrichum gloeosporiodes. Plant Dis. 1980, 64, 854–856. [Google Scholar] [CrossRef]
- Li, P.L.; Li, J.; Zhan, X.Y.; Liu, D.; Gong, G.S.; Chen, H.B.; Zhang, M.; Huang, Y.; Yan, J.M. First report of Colletotrichum gloeosporioides sensu stricto causing anthracnose on Euonymus japonicus in Sichuan Province of China. Plant Dis. 2017, 101, 1318. [Google Scholar] [CrossRef]
- Pan, M.; Zhu, H.Y.; Tian, C.M.; Huang, M.; Fan, X.L. Assessment of Cytospora isolates from conifer cankers in China, with the descriptions of four new Cytospora species. Front. Plant Sci. 2021, 12, 636460. [Google Scholar] [CrossRef]
- Shang, Q.J.; Hyde, K.D.; Camporesi, E.; Maharachchikumbura, S.S.N.; Norphanphoun, C.; Brooks, S.; Liu, J.K. Additions to the genus Cytospora with sexual morph in Cytosporaceae. Mycosphere 2020, 11, 189–224. [Google Scholar] [CrossRef]
- Jiang, N.; Li, X.W.; Xue, H.; Piao, C.G.; Li, Y. Identification of Kerria japonica f. pleniflora branch canker related fungi based on morphological and molecular methods. J. Terr. Ecosyst. Conserv. 2022, 2, 50–56. (In Chinese) [Google Scholar]
- Fan, X.L.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Ma, R.; Tian, C.M. Cytospora species associated with canker disease of three anti-desertification plants in northwestern China. Phytotaxa 2015, 197, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Saccardo, P.A. Sylloge Fungorum; Patavii: Sumptibus auctoris. Typis seminari; Berlin: Iterum impressum apud R. Friedländer & Sohn: Berlin, Germany, 1888. [Google Scholar]
- Chen, M.M. Forest Fungi Phytogeography, Forest Fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; University and Jepson Herbaria: Sacramento, CA, USA, 2002. [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China. Mycotaxon: Ithaca, NY, USA, 2005. [Google Scholar]
- Fan, X.L.; Liang, Y.M.; Ma, R.; Tian, C.M. Morphological and phylogenetic studies of Cytospora (Valsaceae, Diaporthales) isolates from Chinese scholar tree, with description of a new species. Mycoscience 2014, 55, 252–259. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. (In Chinese) [Google Scholar]
- Cooke, M.C. New British Fungi. Grevillea 1885, 14, 1–7. [Google Scholar]
- Saccardo, P.A. Sylloge Fungorum; Typis Seminarii: Berlin/Heidelberg, Germany, 1892; Volume 10. [Google Scholar]
- Norphanphoun, C.; Gentekaki, E.; Hongsana, S.; Jayawardena, R.; Senanayake, I.C.; Manawasinghe, I.S.; Abeywickrama, P.D.; Bhunjun, C.S.; Hyde, K.D. Diaporthe, formalizing the species-group concept. Mycosphere 2022, 13, 752–819. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe, phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.C.; Wingfield, M.J.; Common, R.; Roux, J. Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Stud. Mycol. 2005, 52, 1–144. [Google Scholar]
- Moyo, P.; Mostert, L.; Spies, C.F.; Damm, U.; Halleen, F. Diversity of Diatrypaceae species associated with dieback of Grapevines in South Africa, with the description of Eutypa cremea sp. nov. Plant Dis. 2018, 102, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.S.; Lovecká, P.; Mrnka, L.; Vychodilová, A.; Strejček, M.; Fenclová, M.; Demnerová, K.J.M.E. Distinct communities of poplar endophytes on an unpolluted and a risk element-polluted site and their plant growth-promoting potential in vitro. Microb. Ecol. 2018, 75, 955–969. [Google Scholar] [CrossRef]
- Tang, W.; Ding, Z.; Zhou, Z.Q.; Wang, Y.Z.; Guo, L.Y. Phylogenetic and pathogenic analyses show that the causal agent of apple ring rot in China is Botryosphaeria dothidea. Plant Dis. 2012, 96, 486–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.Z.; Xu, T. Cytological characteristics of the infection in different species, varieties and organs of persimmon by Colletotrichum gloeosporioides. Mycosystema 2005, 24, 116–122. (In Chinese) [Google Scholar]
- Wang, Q.; Liu, Z.; He, W.; Zhang, Y. Pseudocercospora spp. from leaf spots of Euonymus japonicus in China. Mycosystema 2019, 38, 159–170. [Google Scholar]
- Huang, J.G.; Chi, M.Y.; Sun, X.M.; Qian, H.W.; Liang, W.X.; Zhou, X.F. First Report of Powdery Mildew Caused by Erysiphe alphitoides on Euonymus japonicas and Its Natural Teleomorph Stage in China. Plant Dis. 2017, 101, 387. [Google Scholar] [CrossRef]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); CBS Biodiversity Series; CBS: Utrecht, The Netherlands, 2012; Volume 11. [Google Scholar]
- Li, C.W.; Zhang, Y.; Liu, Y.; Kang, J.M.; Ma, X.M.; Fu, L.L. First report of powdery mildew caused by Erysiphe euonymi-japonici on Euonymus japonicus in central China. Plant Dis. 2011, 95, 611. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Pan, Y.; Di, R.; He, Q.; Rajaofera, M.J.N.; Liu, W.; Zheng, F.; Miao, W. Molecular identification of the powdery mildew fungus infecting rubber trees in China. For. Pathol. 2019, 49, e12519. [Google Scholar] [CrossRef]
Figure 1.
Diseased Euonymus japonicus in Beijing: (a–e) Symptoms of branches dieback; (f) Symptoms of leaf spots.
Figure 1.
Diseased Euonymus japonicus in Beijing: (a–e) Symptoms of branches dieback; (f) Symptoms of leaf spots.

Figure 2.
Different symptoms of the diseases of Euonymus japonicus: (a–j) Leaves spots with different symptoms; (k–q) Different morphological characteristics of reproductive structure on diseased branches.
Figure 2.
Different symptoms of the diseases of Euonymus japonicus: (a–j) Leaves spots with different symptoms; (k–q) Different morphological characteristics of reproductive structure on diseased branches.

Figure 3.
Phylogram of Colletotrichum gloeosporioides complex based on maximum likelihood (ML) analysis of the dataset of combined ITS, act, tub2, chs1, and gaphd genes. ML bootstrap support values above 70% are shown near nodes. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type isolates are in bold. Isolates highlighted with blue colours were obtained in this study.
Figure 3.
Phylogram of Colletotrichum gloeosporioides complex based on maximum likelihood (ML) analysis of the dataset of combined ITS, act, tub2, chs1, and gaphd genes. ML bootstrap support values above 70% are shown near nodes. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type isolates are in bold. Isolates highlighted with blue colours were obtained in this study.

Figure 4.
Phylogram of Cytospora based on maximum likelihood (ML) analysis of the dataset of combined ITS, act, rpb2, tef1-α, and tub2 genes. ML bootstrap support values above 70% are shown near nodes. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type isolates are in bold. Isolates highlighted with blue colours were obtained in this study.
Figure 4.
Phylogram of Cytospora based on maximum likelihood (ML) analysis of the dataset of combined ITS, act, rpb2, tef1-α, and tub2 genes. ML bootstrap support values above 70% are shown near nodes. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type isolates are in bold. Isolates highlighted with blue colours were obtained in this study.

Figure 5.
Phylogram of Pestalotiopsis based on maximum likelihood (ML) analysis of the dataset of combined ITS, tef1-α, and tub2 genes. ML bootstrap support values above 70% are shown near nodes. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type isolates are in bold. Isolates highlighted with blue colours were obtained in this study.
Figure 5.
Phylogram of Pestalotiopsis based on maximum likelihood (ML) analysis of the dataset of combined ITS, tef1-α, and tub2 genes. ML bootstrap support values above 70% are shown near nodes. Thickened branches represent posterior probabilities above 0.95 from BI. Ex-type isolates are in bold. Isolates highlighted with blue colours were obtained in this study.

Figure 6.
Colletotrichum euonymi (ex-holotype culture CFCC 55542): (a,b) Symptoms caused by Colletotrichum euonymi; (c) Front colony on PDA (10 d); (d) Conidiama on PDA (30 d); (e–g) Conidiophores and conidiogenous cells formed on inoculated Euonymus japonicus; (h,i) Conidia formed on inoculated Euonymus japonicus. Scale bars = 10 μm.
Figure 6.
Colletotrichum euonymi (ex-holotype culture CFCC 55542): (a,b) Symptoms caused by Colletotrichum euonymi; (c) Front colony on PDA (10 d); (d) Conidiama on PDA (30 d); (e–g) Conidiophores and conidiogenous cells formed on inoculated Euonymus japonicus; (h,i) Conidia formed on inoculated Euonymus japonicus. Scale bars = 10 μm.

Figure 7.
Colletotrichum euonymicola (ex-holotype culture CFCC 55486): (a,b) Symptoms caused by Colletotrichum euonymicola; (c) Front colony on PDA (10 d); (d–h) Conidiogenous cells formed on PDA; (i–k) Appiospora formed on PDA; (l) Conidia formed on PDA. Scale bars = 10 μm.
Figure 7.
Colletotrichum euonymicola (ex-holotype culture CFCC 55486): (a,b) Symptoms caused by Colletotrichum euonymicola; (c) Front colony on PDA (10 d); (d–h) Conidiogenous cells formed on PDA; (i–k) Appiospora formed on PDA; (l) Conidia formed on PDA. Scale bars = 10 μm.

Figure 8.
Cytospora zhaitangensis (holotype BJFC CF20220144): (a) Diseased Euonymus japonicus branches caused by Cytospora zhaitangensis in the field; (b,c) Habit of conidiomata on twig; (d) Transverse section of a conidioma; (e) Longitudinal section through a conidioma; (f) Conidiophores and conidiogenous cells; (g) Conidia. Scale bars: 500 µm (b–e), 10 µm (f,g).
Figure 8.
Cytospora zhaitangensis (holotype BJFC CF20220144): (a) Diseased Euonymus japonicus branches caused by Cytospora zhaitangensis in the field; (b,c) Habit of conidiomata on twig; (d) Transverse section of a conidioma; (e) Longitudinal section through a conidioma; (f) Conidiophores and conidiogenous cells; (g) Conidia. Scale bars: 500 µm (b–e), 10 µm (f,g).

Figure 9.
Pestalotiopsis chaoyangensis (ex-holotype culture CFCC 55549): (a) Diseased Euonymus japonicus leaves caused by Pestalotiopsis chaoyangensis; (b) Colony on PDA at 30 days; (c) Conidial masses formed on PDA; (d) Conidiogenous cells; (e) Conidia. Scale bars: 10 µm.
Figure 9.
Pestalotiopsis chaoyangensis (ex-holotype culture CFCC 55549): (a) Diseased Euonymus japonicus leaves caused by Pestalotiopsis chaoyangensis; (b) Colony on PDA at 30 days; (c) Conidial masses formed on PDA; (d) Conidiogenous cells; (e) Conidia. Scale bars: 10 µm.

Figure 10.
Lesions (1, 14 d; 2, 17d) resulting from inoculation of three novel species isolated from leaves onto Euonymus japonicus. Disease symptoms inoculated with (a) Colletotrichum euonymi (CFCC 55542); (b) Colletotrichum euonymicola (CFCC 55486); (c) Pestalotiopsis chaoyangensis (CFCC 55549); (d) Blank control.
Figure 10.
Lesions (1, 14 d; 2, 17d) resulting from inoculation of three novel species isolated from leaves onto Euonymus japonicus. Disease symptoms inoculated with (a) Colletotrichum euonymi (CFCC 55542); (b) Colletotrichum euonymicola (CFCC 55486); (c) Pestalotiopsis chaoyangensis (CFCC 55549); (d) Blank control.

Figure 11.
Average lesion diameter (mm) resulting from inoculation with Euonymus japonicus. Vertical bars represent standard error of means. Different letters above the bars indicate treatments that were significantly different (α = 0.05).
Figure 11.
Average lesion diameter (mm) resulting from inoculation with Euonymus japonicus. Vertical bars represent standard error of means. Different letters above the bars indicate treatments that were significantly different (α = 0.05).


{kind=link}
{kind=link}
{kind=link}
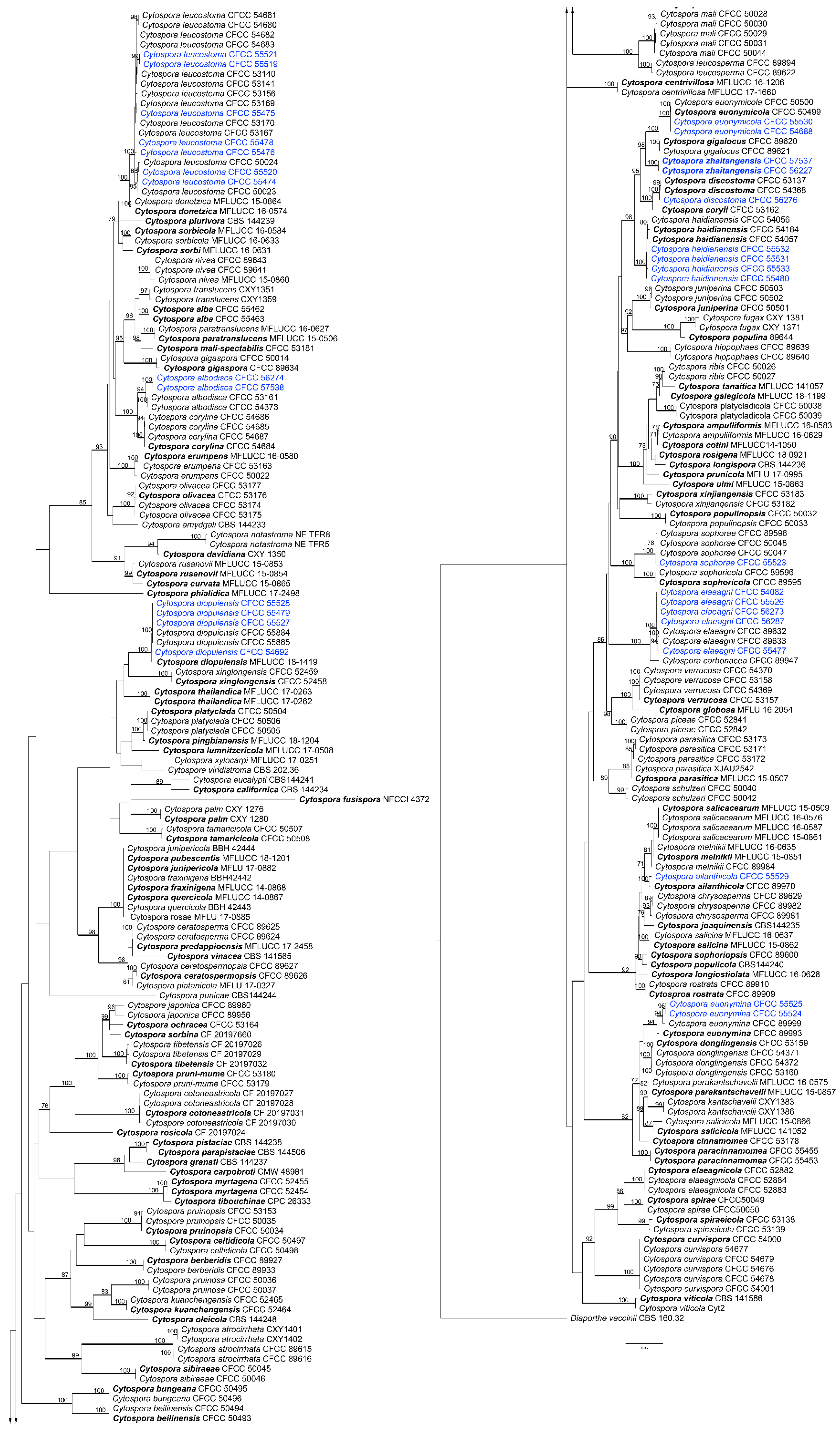{kind=link}
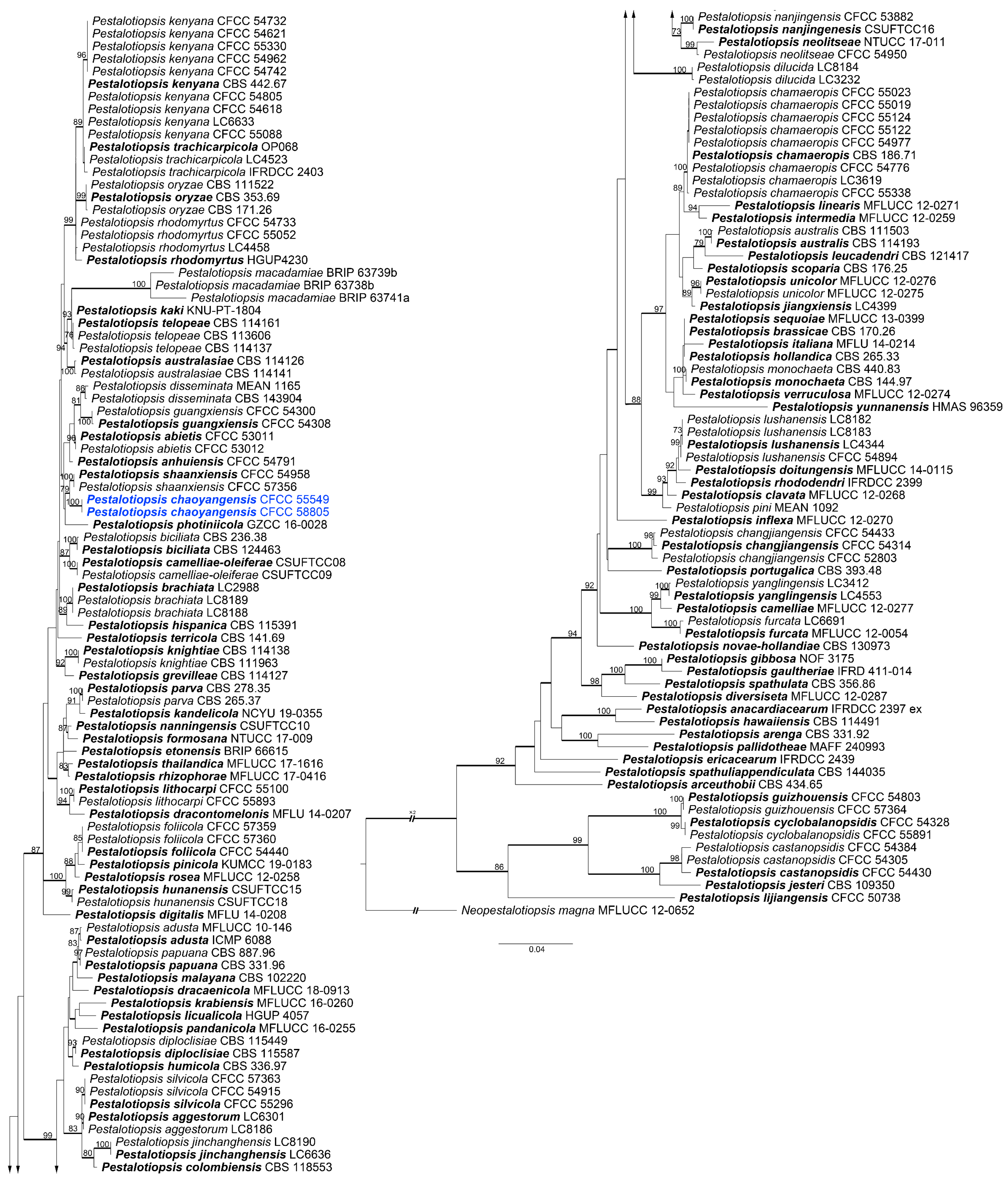{kind=link}
{kind=link}
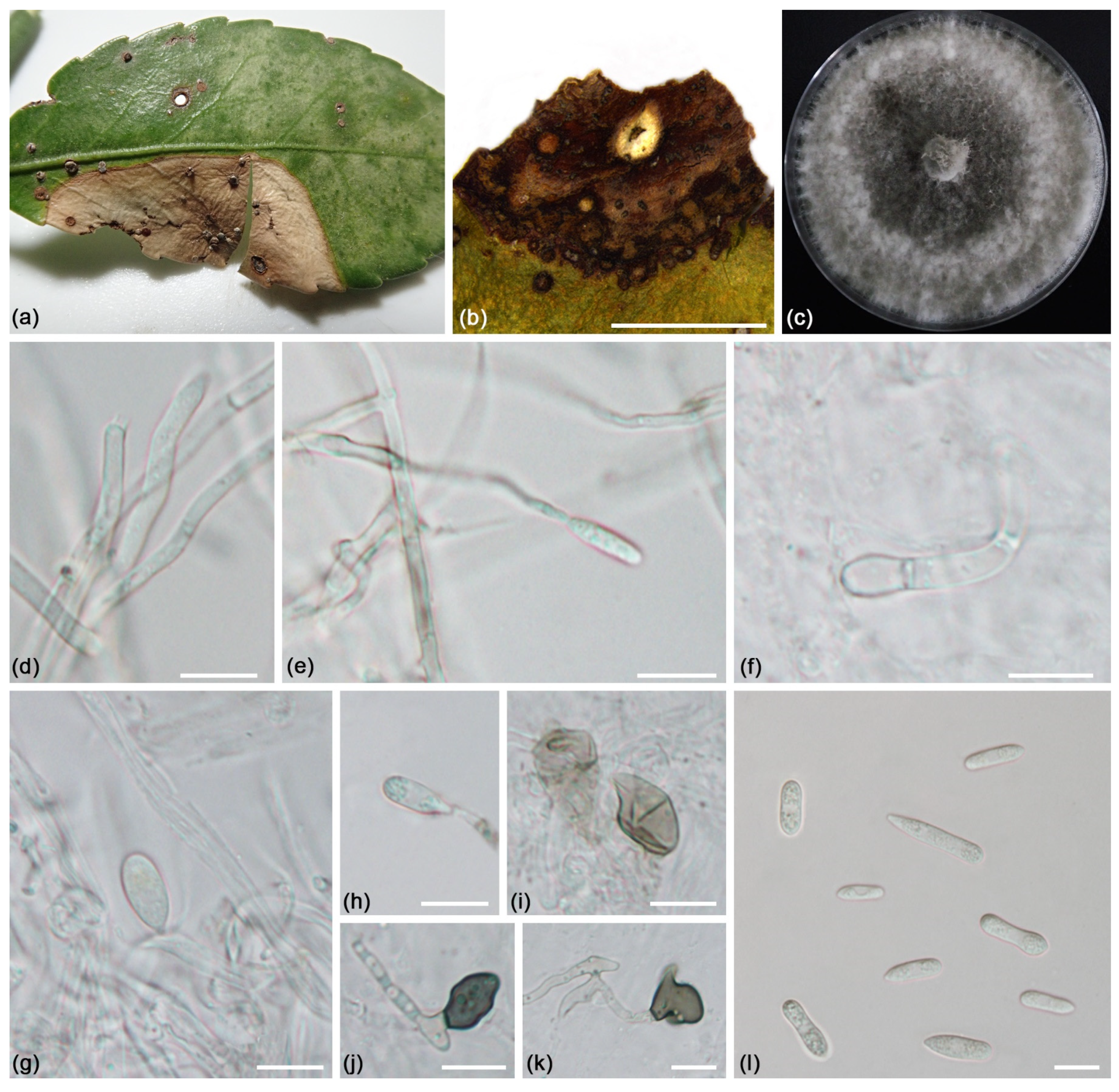{kind=link}
{kind=link}
{kind=link}
{kind=link}
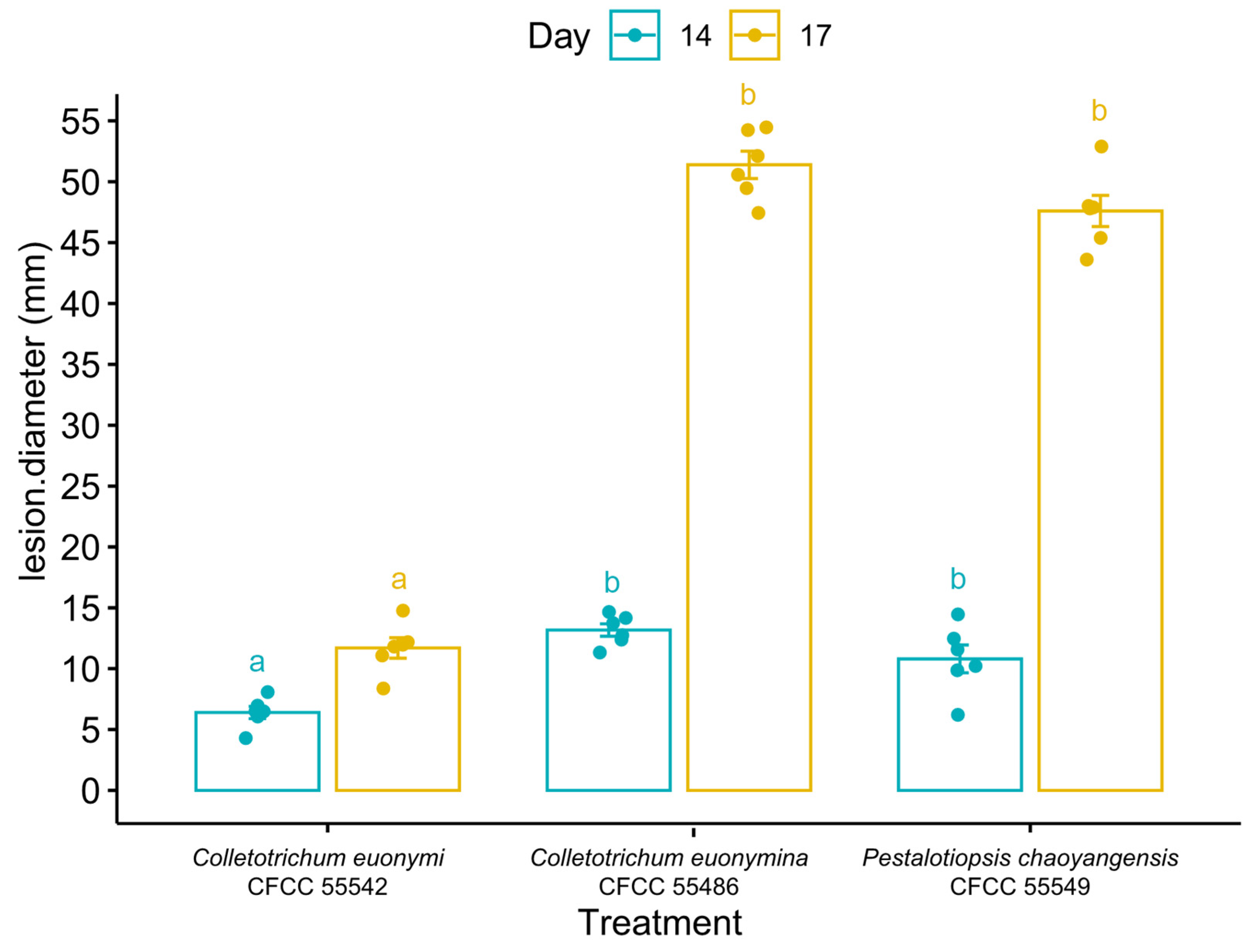{kind=link}
Table 1.
Genes used in this study with PCR primers and optimal annealing temperature.
| Locus | PCR Primers | PCR: Thermal Cycles: (Annealing Temp. in Bold) | Reference | Genera |
|---|---|---|---|---|
| ITS | ITS1/ITS4 | (95 °C: 30 s, 51 °C: 30 s, 72 °C: 1 min) × 35 cycles | [45] | Aplosporella, Botryosphaeria, Cytospora, Colletotrichum, Diaporthe, Dothiorella, and Pestalotiopsis |
| tef1-α | EF1-728F/EF1-986R | (95 °C: 15 s, 55 °C: 20 s, 72 °C: 1 min) × 35 cycles | [46] | Aplosporella, Botryosphaeria, Cytospora, Diaporthe, Dothiorella, and Pestalotiopsis |
| tub2 | Bt2a/Bt2b | (95 °C: 30 s, 55 °C: 30 s, 72 °C: 1 min) × 35 cycles | [47] | Botryosphaeria, Cytospora, Colletotrichum, Diaporthe, Dothiorella, and Pestalotiopsis |
| act | ACT-512F/ACT-783R | (95 °C: 45 s, 55 °C: 45 s, 72 °C: 1 min) × 35 cycles | [46] | Cytospora and Colletotrichum |
| rpb2 | RPB2-5F/RPB2-7cR | (95 °C: 30 s, 52 °C: 1 min, 72 °C: 1 min) × 35 cycles | [46] | Cytospora |
| chs1 | CHS-79F/CHS-345R | (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 35 cycles | [46] | Colletotrichum |
| gaphd | GDF1/GDF2 | (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 35 cycles | [49] | Colletotrichum |
| cal | CAL228F/CAL737R | (95 °C: 15 s, 55 °C: 20 s, 72 °C: 1 min) × 35 cycles | [46] | Diaporthe |
| his3 | CYLH3F/H3-1b | (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 35 cycles | [47,50] | Diaporthe |
Table 2.
Substitution models used for Bayesian analyses in this study.
| Analysis | Number of Ingroup Sequences | Outgroup | Substitution Models Used for Bayesian Analyses/Number of Characters with Gaps | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ITS | tef1 | tub2 | act | ITS | chs | gaphd | cal | ITS | |||
| Aplosporella 2-genes | 27 | Alanomyces indica CBS 134264 | SYM + G/558 | HKY + G/317 | – | – | – | – | – | – | – |
| Botryosphaeria 3-genes | 71 | Cophinforma eucalypti CBS 134651 | GTR + I/448 | GTR + G/266 | HKY + G/415 | – | – | – | – | – | – |
| Colletotrichum 1-gene | 487 | Monilochaetes infuscans CBS 869.96 | GTR + I + G/611 | – | – | – | – | – | – | – | – |
| Colletotrichum gloeosporioides complex 5-genes | 67 | Colletotrichum vietnamense CBS 125477 | GTR + I + G/555 | – | HKY + G/451 | HKY + G/290 | – | K80 + I + G/275 | HKY + G/278 | – | – |
| Colletotrichum vietnamense CBS 125478 | |||||||||||
| Cytospora 5-genes | 277 | Diaporthe vaccinii CBS 160.32 | GTR + I + G/626 | HKY + I + G/788 | HKY + I + G/680 | GTR + I + G/347 | GTR + I + G/732 | – | – | – | – |
| Diaporthe 5-genes | 307 | Diaporthella corylina CBS 121124 | GTR + I + G/551 | HKY + I + G/678 | GTR + I + G/533 | – | – | – | – | HKY + I + G/625 | GTR + I + G/625 |
| Aplosporella 2-genes | 27 | Alanomyces indica CBS 134264 | SYM + G/558 | HKY + G/317 | – | – | – | – | – | – | – |
| Botryosphaeria 3-genes | 71 | Cophinforma eucalypti CBS 134651 | GTR + I/448 | GTR + G/266 | HKY + G/415 | – | – | – | – | – | – |
Table 3.
The statistics of ML trees in this study.
| Analysis | Figure Number | Estimated Base Frequencies | Substitution Rates | Gamma Distribution Shape Parameter α A | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | T | C | G | AC | A | T | C | CT | GT | |||
| Aplosporella | Figure S1 | 0.212988 | 0.257488 | 0.274402 | 0.255122 | 1.887152 | 3.178340 | 1.573282 | 1.399408 | 2.980772 | 1.000000 | 0.382287 |
| Botryosphaeria | Figure S2 | 0.215054 | 0.222293 | 0.312418 | 0.250235 | 0.441342 | 2.413944 | 0.639018 | 0.521572 | 3.599492 | 1.000000 | 0.240522 |
| Colletotrichum | Figure S3 | 0.227417 | 0.235495 | 0.284682 | 0.252405 | 1.597190 | 2.099216 | 1.489264 | 1.500977 | 5.675789 | 1.000000 | 0.260939 |
| Colletotrichum gloeosporioides complex | Figure 3 | 0.229013 | 0.228092 | 0.298051 | 0.244843 | 0.821304 | 2.465133 | 0.850090 | 0.790048 | 4.448447 | 1.000000 | 0.393050 |
| Cytospora | Figure 4 | 0.244498 | 0.230131 | 0.286014 | 0.239357 | 1.288489 | 3.541841 | 1.487837 | 0.962637 | 6.197985 | 1.000000 | 0.360802 |
| Diaporthe | Figure S4 | 0.213713 | 0.223461 | 0.326805 | 0.236021 | 1.074776 | 3.154257 | 1.134578 | 0.879474 | 4.526775 | 1.000000 | 0.509636 |
| Dothiorella | Figure S5 | 0.204842 | 0.229767 | 0.320640 | 0.244750 | 0.762653 | 1.883624 | 1.132143 | 0.814926 | 3.755328 | 1.000000 | 0.228281 |
| Pestalotiopsis | Figure 5 | 0.232209 | 0.258050 | 0.294948 | 0.214793 | 0.941298 | 2.870198 | 0.998145 | 0.973783 | 3.778432 | 1.000000 | 0.294632 |
Table 4.
Lesions diameter (mm) of inoculated leaves.
| Treatment | Days after Inoculation | Lesion Diameter (mm) | Average (mm) | Sd. (mm) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Colletotrichum euonymi CFCC 55542 | 14 | 6.5 | 7.0 | 8.1 | 4.3 | 6.5 | 6.1 | 6.4 | 1.24 |
| 17 | 11.8 | 12.0 | 14.8 | 8.4 | 12.2 | 11.1 | 11.7 | 2.06 | |
| Colletotrichum euonymicola CFCC 55486 | 14 | 12.4 | 13.7 | 14.2 | 11.3 | 12.7 | 14.7 | 13.2 | 1.24 |
| 17 | 49.5 | 52.1 | 54.2 | 47.4 | 50.6 | 54.5 | 51.4 | 2.76 | |
| Pestalotiopsis chaoyangensis CFCC 55549 | 14 | 9.9 | 10.2 | 14.5 | 6.2 | 12.5 | 11.6 | 10.8 | 2.79 |
| 17 | 45.4 | 47.8 | 52.9 | 43.6 | 48.0 | 47.9 | 47.6 | 3.14 | |
| Blank control | 14 | 4 | 4 | 4 | 4 | 4 | 4 | 4.0 | 0.0 |
| 17 | 4 | 4 | 4 | 4 | 4 | 4 | 4.0 | 0.0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, L.; Pan, M.; Gao, H.; Tian, C.; Fan, X. The Potential Fungal Pathogens of Euonymus japonicus in Beijing, China. J. Fungi 2023, 9, 271. https://doi.org/10.3390/jof9020271
AMA Style
Lin L, Pan M, Gao H, Tian C, Fan X. The Potential Fungal Pathogens of Euonymus japonicus in Beijing, China. Journal of Fungi. 2023; 9(2):271. https://doi.org/10.3390/jof9020271
Chicago/Turabian StyleLin, Lu, Meng Pan, Hong Gao, Chengming Tian, and Xinlei Fan. 2023. "The Potential Fungal Pathogens of Euonymus japonicus in Beijing, China" Journal of Fungi 9, no. 2: 271. https://doi.org/10.3390/jof9020271
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.