
How Deep Can the Endophytic Mycobiome Go? A Case Study on Six Woody Species from the Brazilian Cerrado

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area
2.2. Sampling and Collection
2.3. Surface Disinfection and Sample Processing
2.4. Isolation, Purification and Deposit of Cultivable Fungi
2.5. DNA Extraction and Molecular Identification of Isolates
2.6. Metabarcoding Sequencing
2.6.1. eDNA Extraction
2.6.2. Library Construction and Sequencing
2.7. Foliar Analysis
2.8. Statistical Analyses
3. Results
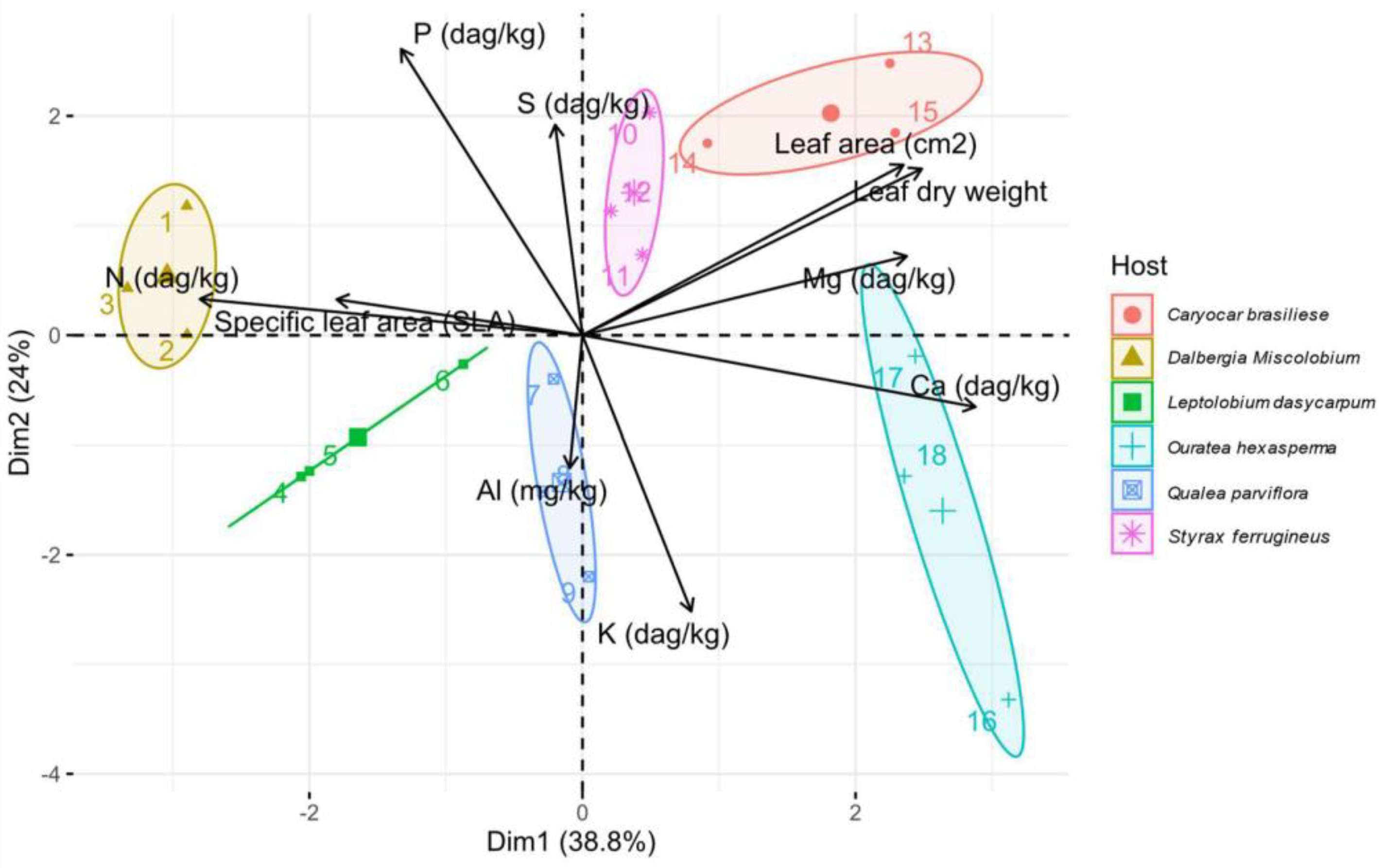
3.1. Analysis of the Nutritional Composition of Leaves
3.2. Cultivation-Dependent Method
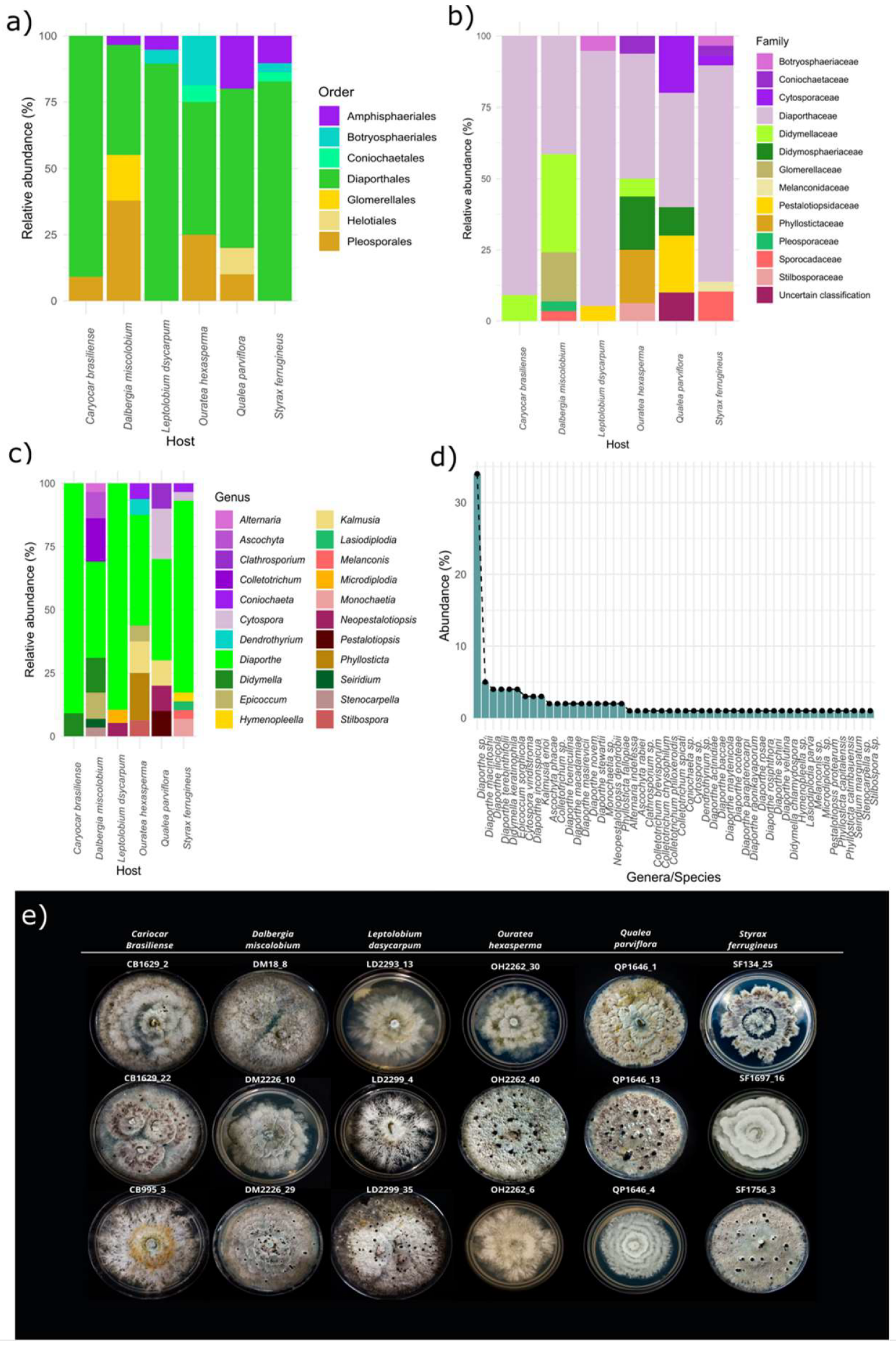
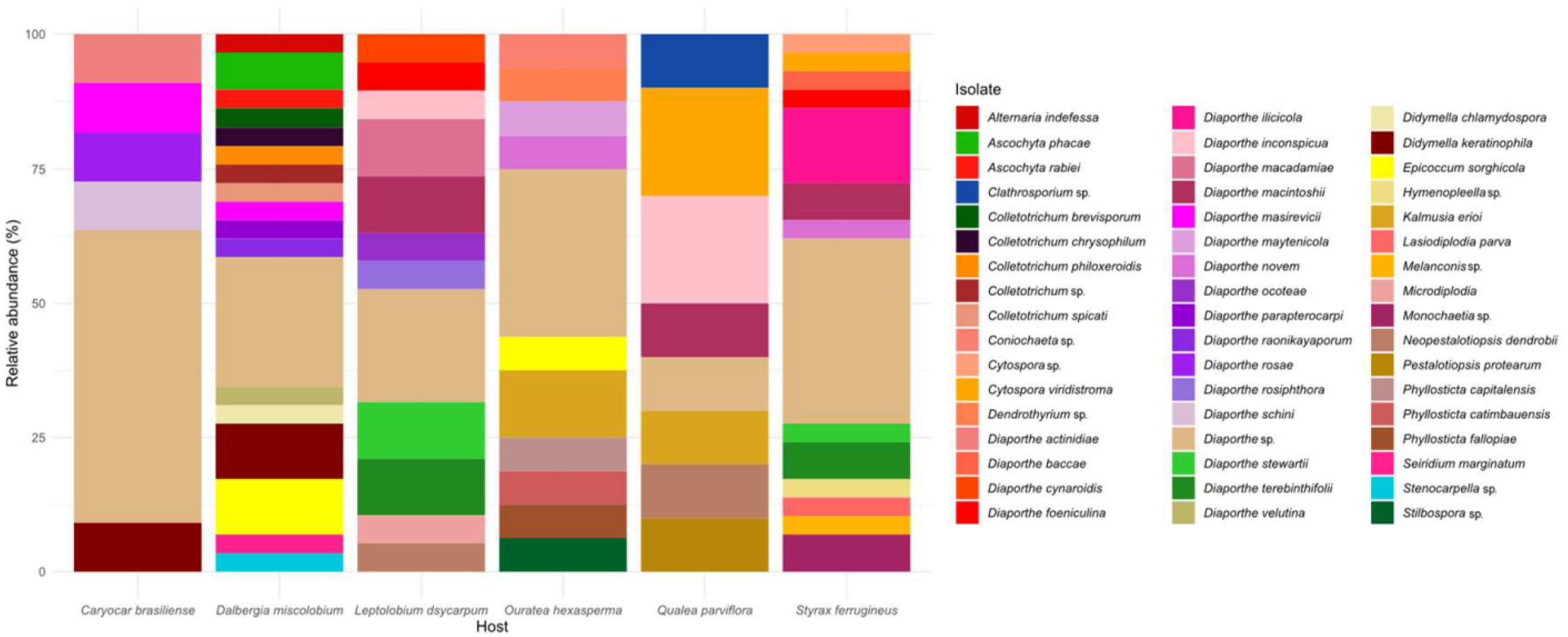
3.2.1. Molecular Identification of Isolates
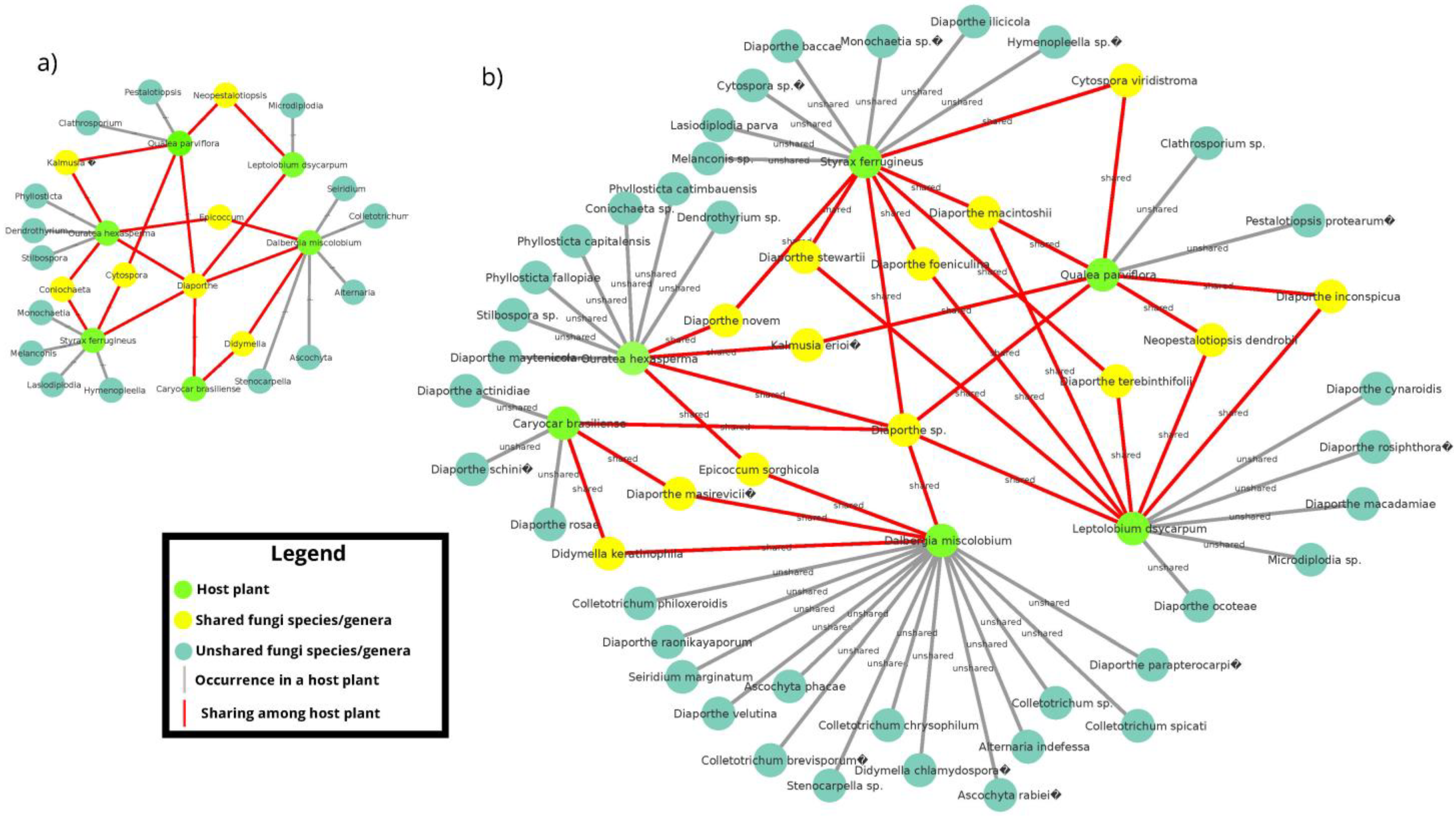
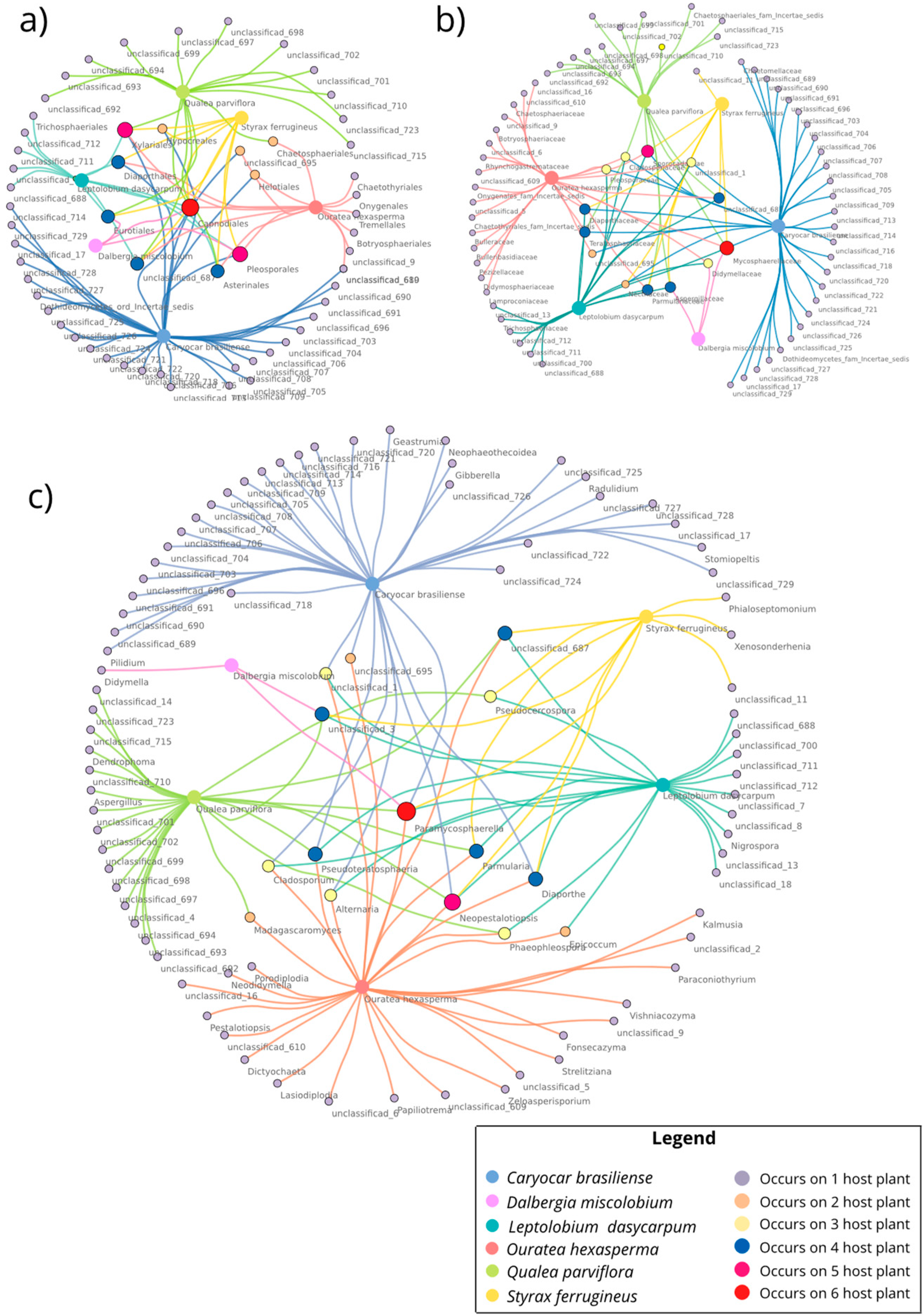
3.2.2. Co-occurrence Networks of Endophytic Fungi from the Cultivation-Dependent Method
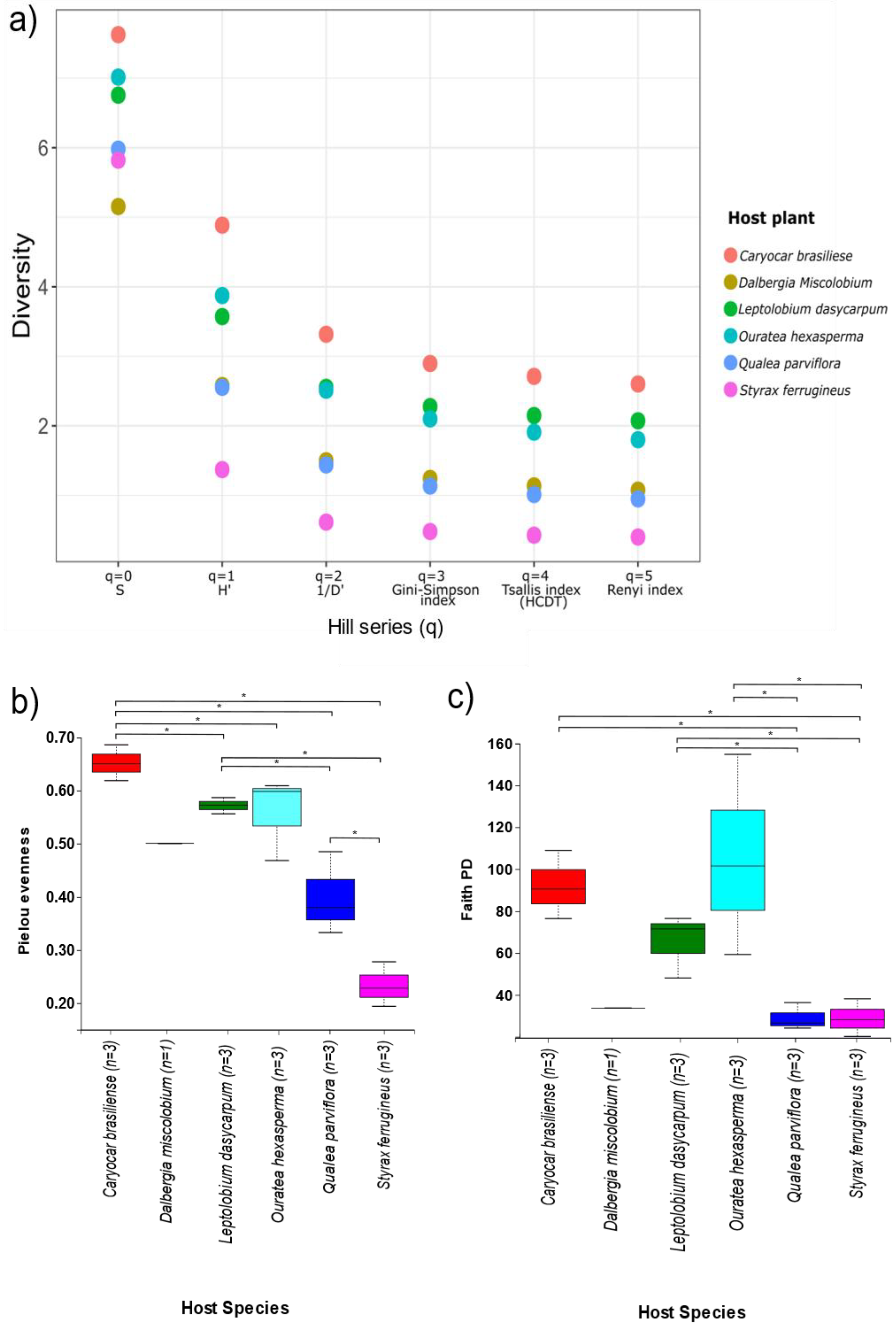
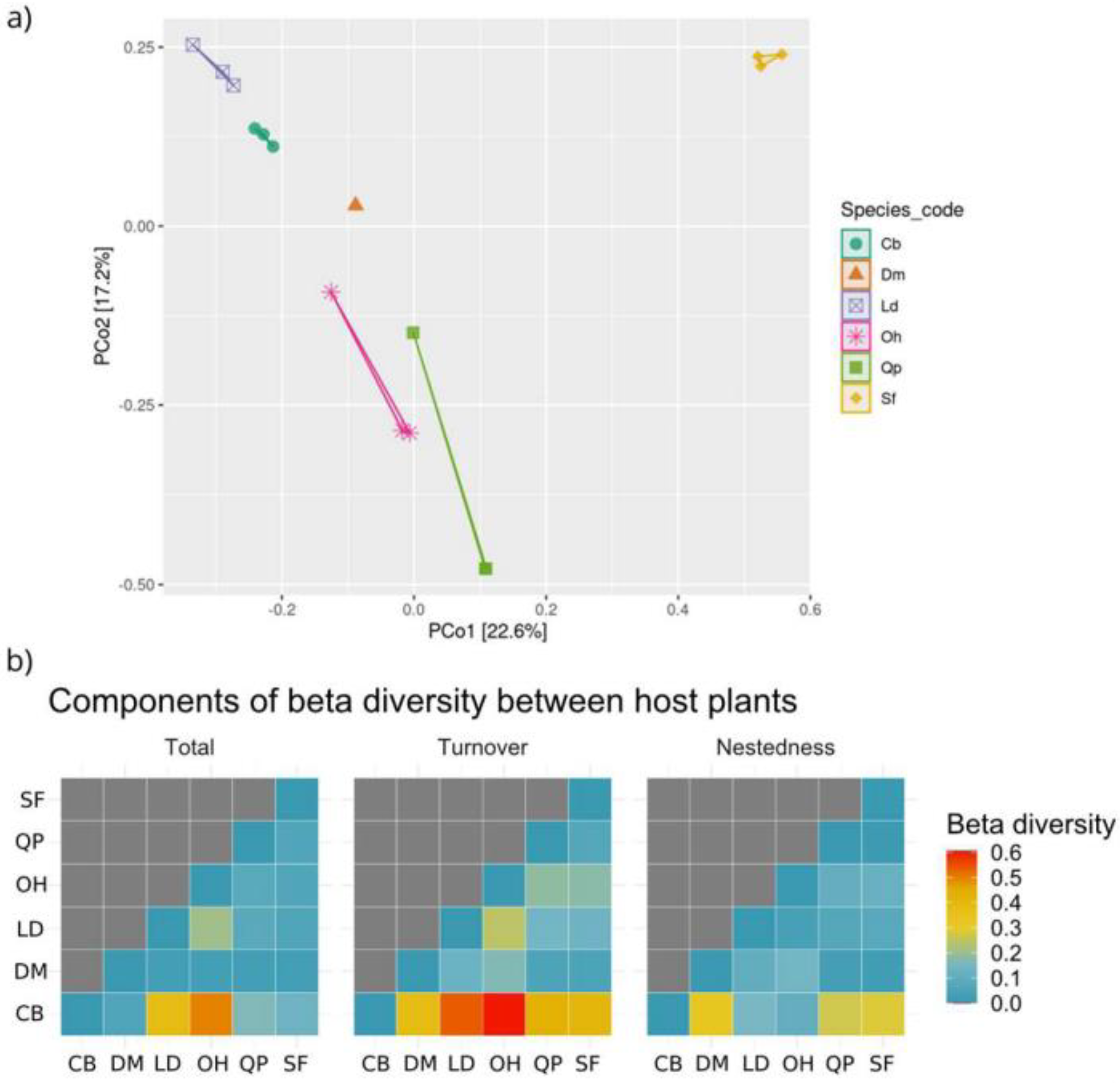
3.2.3. Alpha and Beta Diversity
3.3. Metabarcoding Analysis
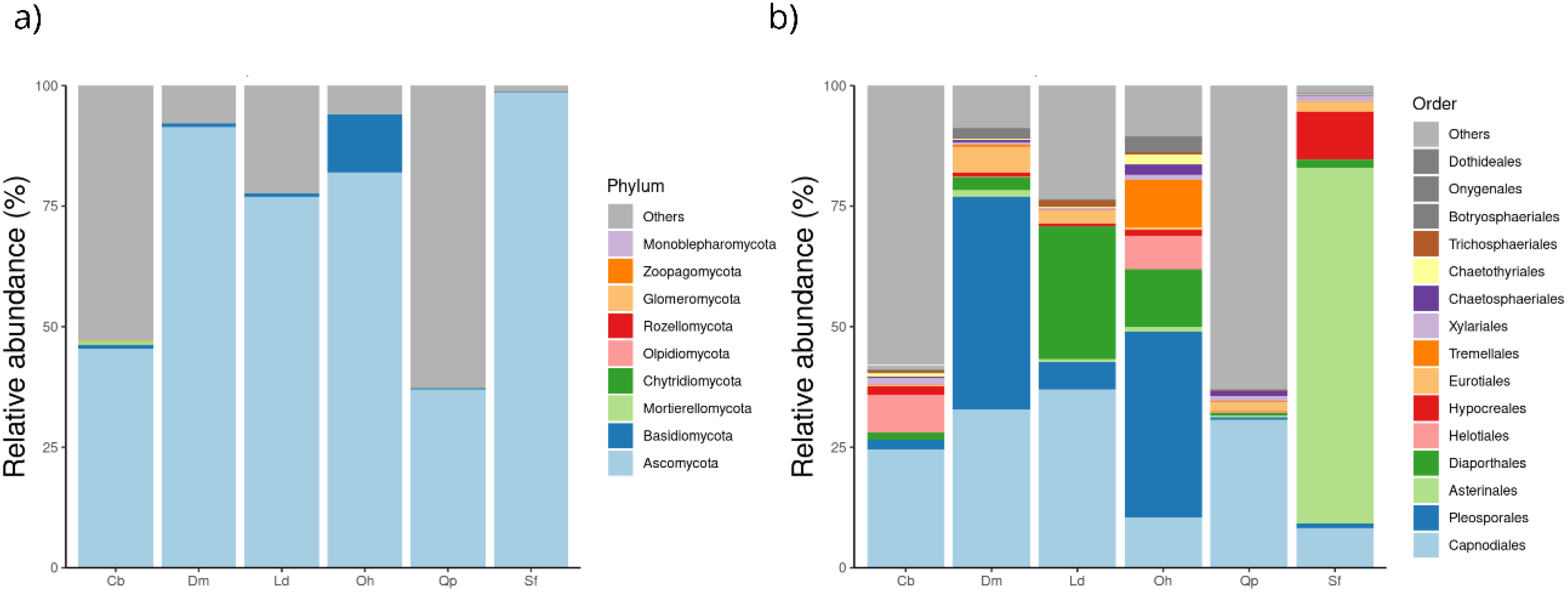
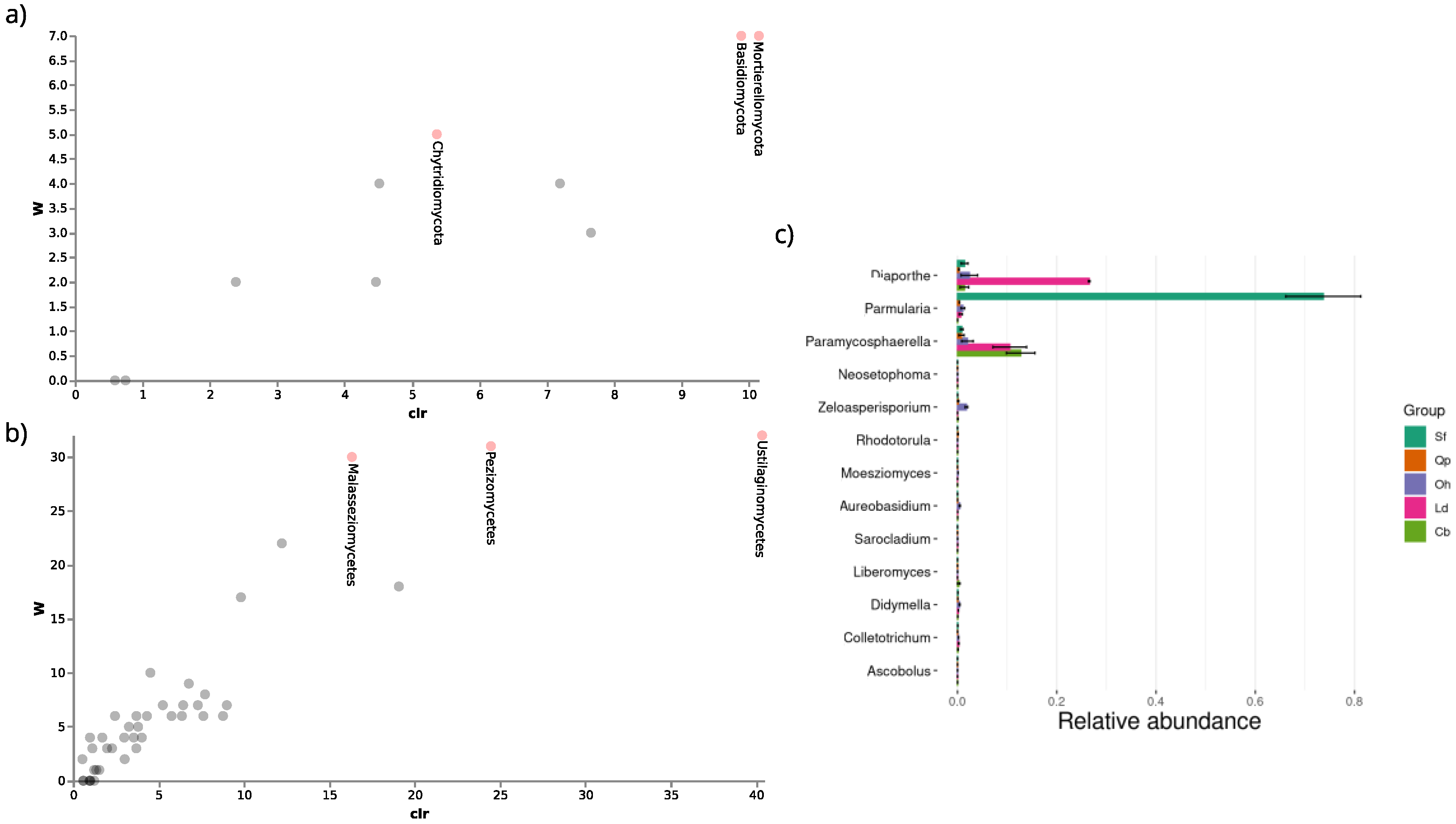
3.3.1. Taxonomic Attributions
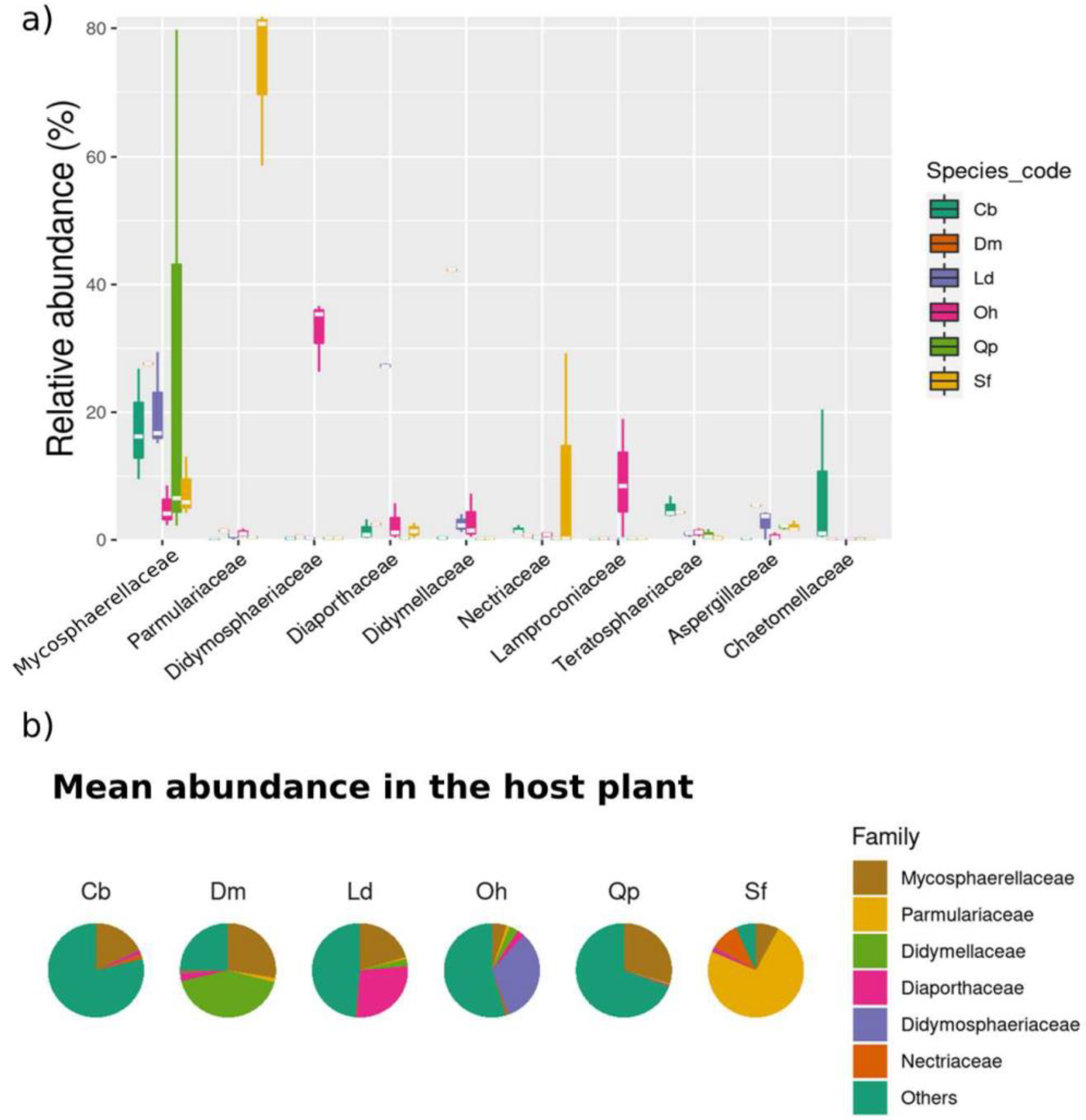
3.3.2. Relative Abundance
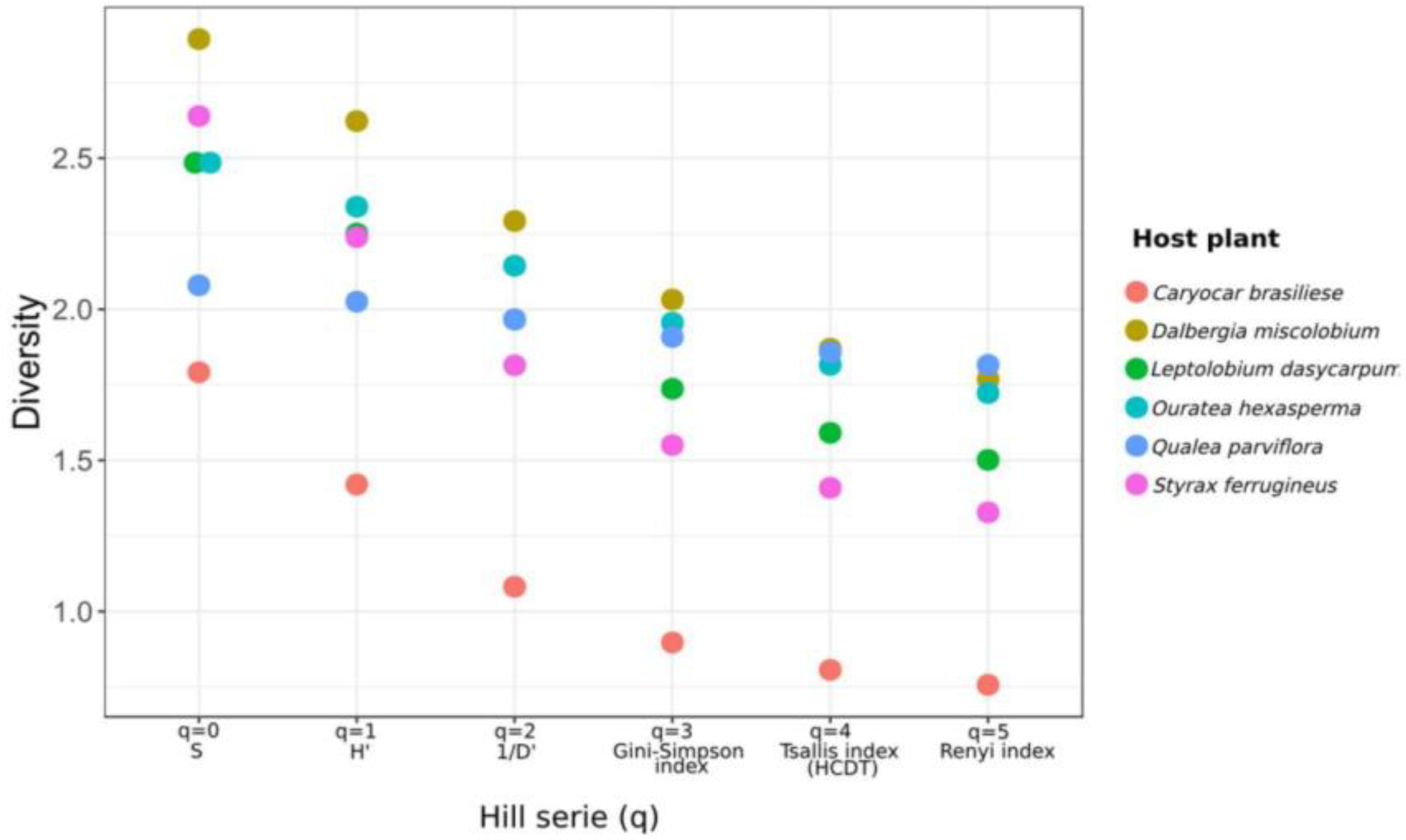
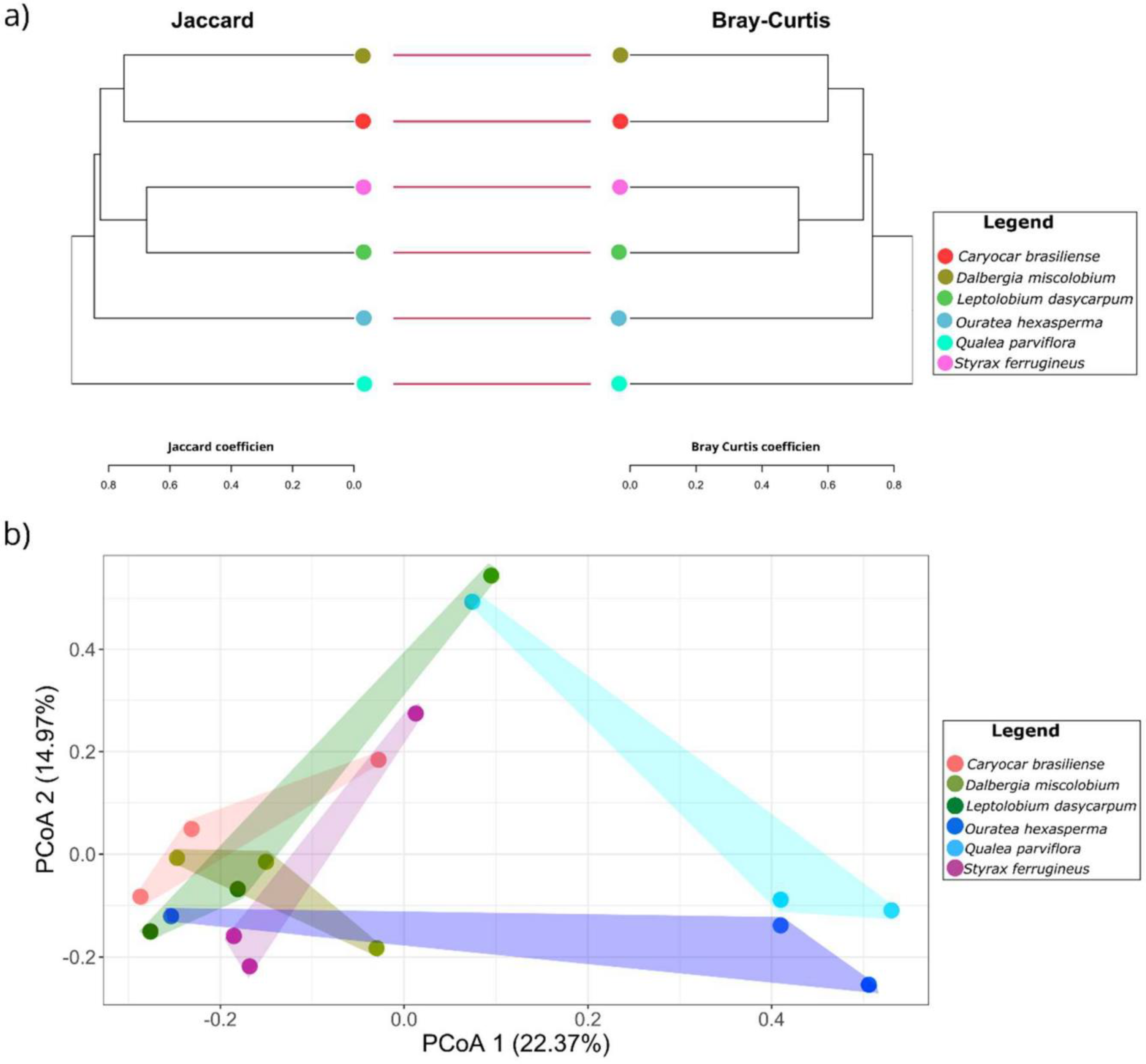
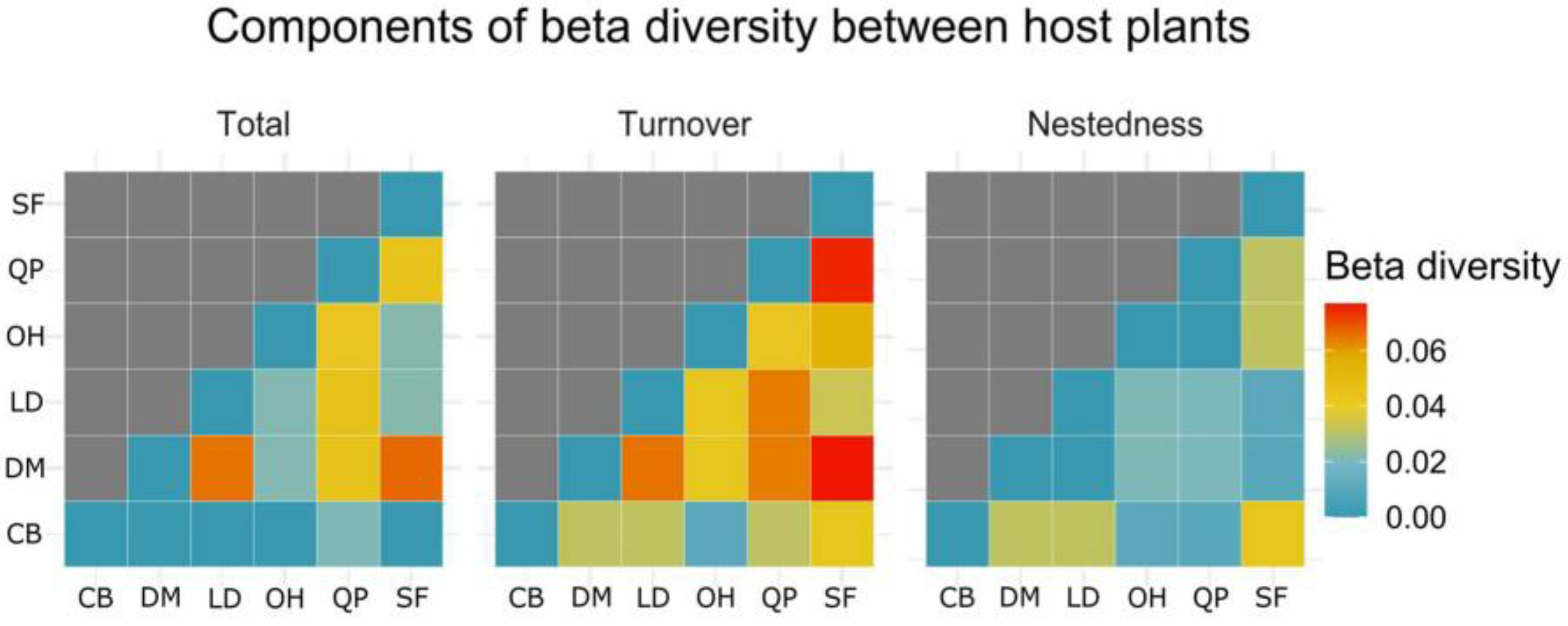
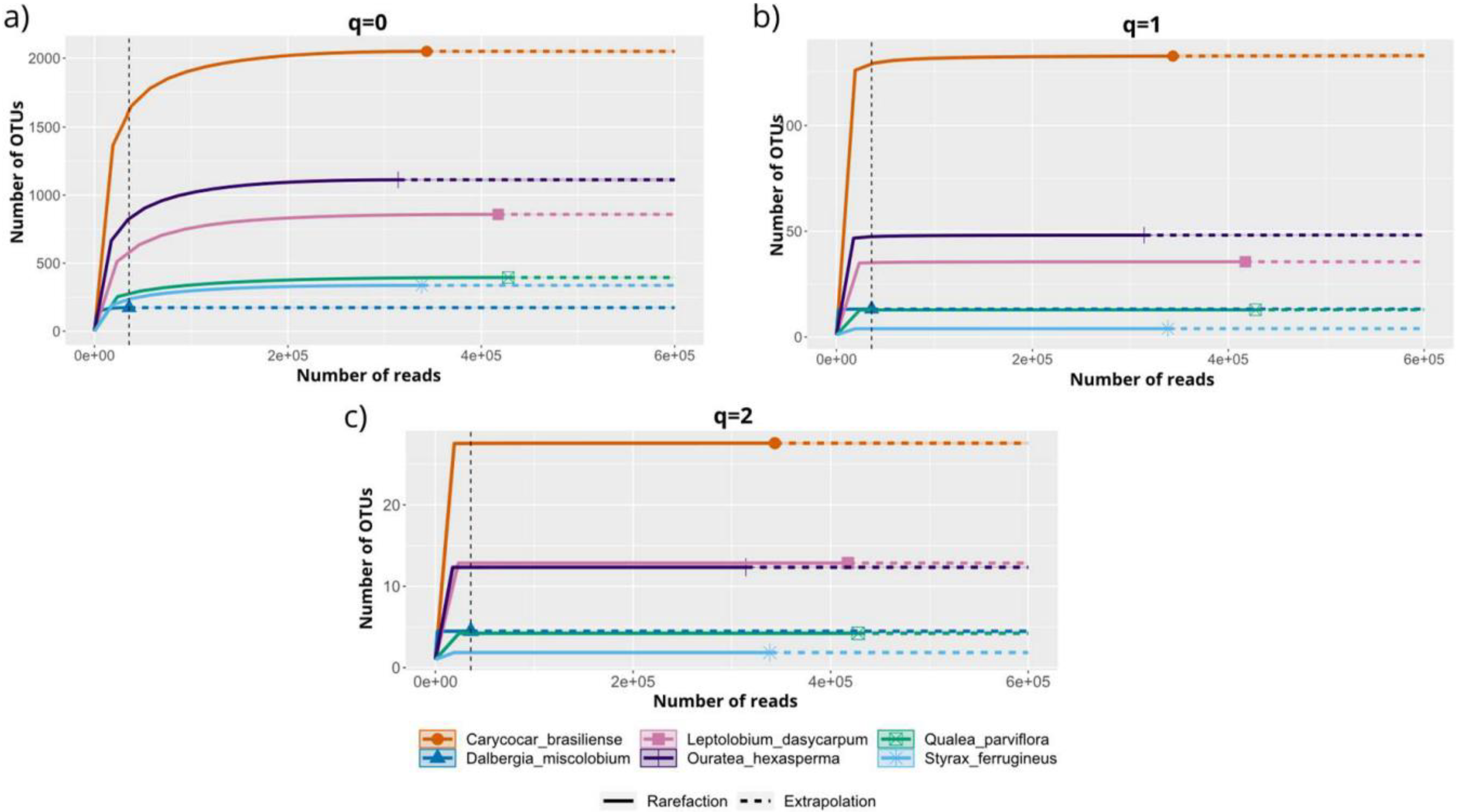
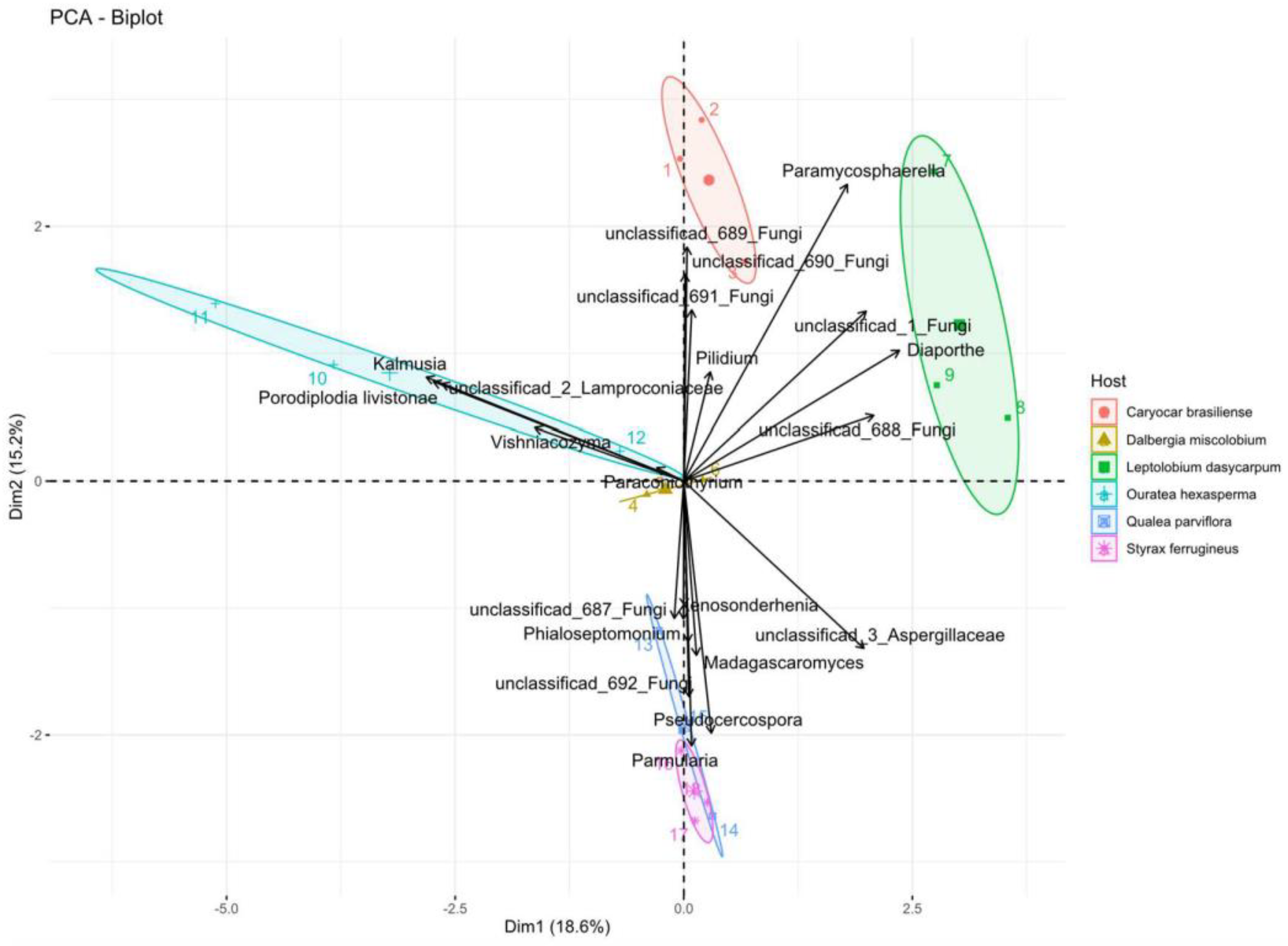
3.3.3. Alpha and Beta Diversity
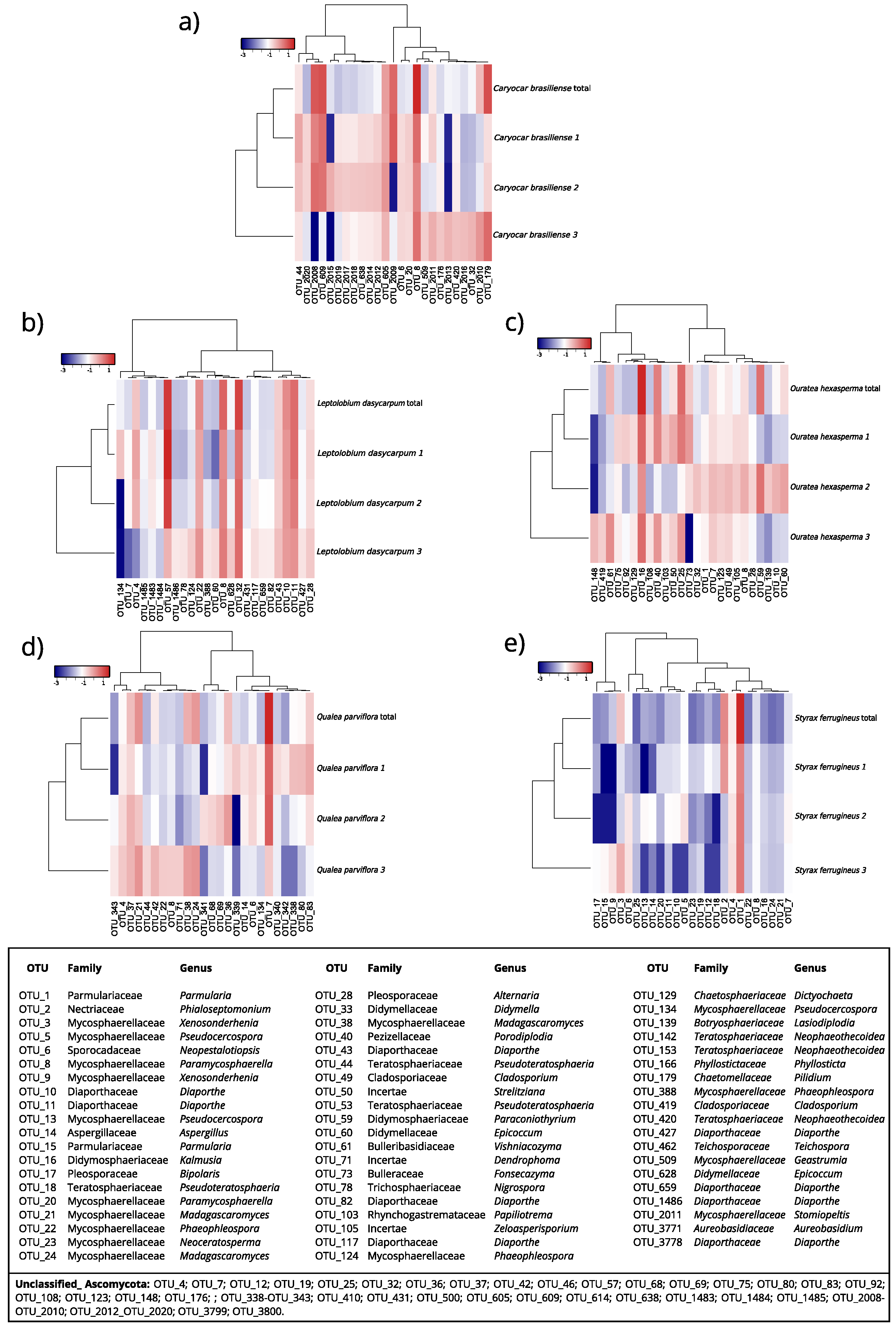
3.3.4. Differential Abundance
3.3.5. Co-occurrence Networks of Endophytic Fungi for Metabarcoding
4. Discussion
4.1. Cultivable Endophytic Fungi
4.2. Characterization of The Endophytic Mycobiome by eDNA Metabarcoding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richards, T.A.; Leonard, G.; Wideman, J.G. What Defines the “Kingdom” Fungi? Microbiol. Spectr. 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D. The numbers of fungi. Fungal Divers. 2022, 114, 1. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.-R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Taylor, J.W.; Ellison, C.E. Mushrooms: Morphological complexity in the fungi. Proc. Natl. Acad. Sci. USA 2010, 107, 11655–11656. [Google Scholar] [CrossRef]
- de Mattos-Shipley, K.M.; Ford, K.L.; Alberti, F.; Banks, A.M.; Bailey, A.M.; Foster, G.D. The good, the bad and the tasty: The many roles of mushrooms. Stud. Mycol. 2016, 85, 125–157. [Google Scholar] [CrossRef]
- Grube, M.; Wedin, M. Lichenized fungi and the evolution of symbiotic organization. Microbiol. Spectr. 2016, 4, 749–765. [Google Scholar] [CrossRef]
- El-Shafey, N.M.; Marzouk, M.A.; Yasser, M.M.; Shaban, S.A.; Beemster, G.T.S.; AbdElgawad, H. Harnessing endophytic fungi for enhancing growth, tolerance and quality of rose-scented geranium (Pelargonium graveolens (L’Hér) Thunb.) Plants under Cadmium Stress: A Biochemical Study. J. Fungi 2021, 7, 1039. [Google Scholar] [CrossRef]
- Fontana, D.; de Paula, S.; Torres, A.; de Souza, V.; Pascholati, S.; Schmidt, D.; Neto, D.D. Endophytic fungi: Biological control and induced resistance to phytopathogens and abiotic stresses. Pathogens 2021, 10, 570. [Google Scholar] [CrossRef]
- Han, D.; Wang, K.; Long, F.; Zhang, W.; Yao, X.; Chen, S. Effects of endophytic fungi on the secondary metabolites of Hordeum bogdanii under alkaline stress. AMB Express 2022, 12, 73. [Google Scholar] [CrossRef]
- Hosseyni Moghaddam, M.S.; Safaie, N.; Rahimlou, S.; Hagh-Doust, N. Inducing tolerance to abiotic stress in Hordeum vulgare L. by Halotolerant Endophytic fungi associated with salt lake plants. Front. Microbiol. 2022, 13, 906365. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Hyde, K.D.; McKenzie, E.H.C.; Saxena, R.K.; Li, Q. Do all fungi have ancestors with endophytic lifestyles? Fungal Divers. 2023. [Google Scholar] [CrossRef]
- Tao, G.; Liu, Z.Y.; Hyde, K.D.; Liu, X.Z.; Yu, Z.N. Whole rDNA analysis reveals novel and endophytic fungi in Bletilla ochracea (Orchidaceae). Fungal Divers. 2008, 33, 101–122. [Google Scholar]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Yao, Z.; Li, J.; Sun, C.; Xia, J.; Wang, B.; Shi, D.; Ren, L. Diversity and antimicrobial activity of endophytic fungi isolated from Securinega suffruticosa in the Yellow River Delta. PLoS ONE 2020, 15, e0229589. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Sun, X.; He, C.; Maitra, P.; Li, X.C.; Guo, L.D. Phyllosphere epiphytic and endophytic fungal community and network structures differ in a tropical mangrove ecosystem. Microbiome 2019, 7, 57. [Google Scholar] [CrossRef]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef]
- Apigo, A.; Oono, R. Dimensions of host specificity in foliar fungal endophytes. In Endophytes of Forest Trees; Pirttilä, A., Frank, A., Eds.; Forestry Sciences; Springer: Cham, Switzerland, 2018; Volume 86. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, Z.Y.; Cui, J.L.; Wang, M.L.; Wang, J.H. Biotransformation ability of endophytic fungi: From species evolution to industrial applications. Appl. Microbiol. Biotechnol. 2021, 105, 7095–7113. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Jayawardena, R.S.; Chen, Y.J.; Konta, S.; Tibpromma, S.; Phukhamsakda, C.; Abeywickrama, P.D.; Samarakoon, M.C.; Senwanna, C.; Mapook, A.; et al. Appressorial interactions with host and their evolution. Fungal Divers. 2021, 110, 75–107. [Google Scholar] [CrossRef]
- Zuo, Y.; Li, X.; Yang, J.; Liu, J.; Zhao, L.; He, X. Fungal endophytic community and diversity associated with desert shrubs driven by plant identity and organ differentiation in extremely arid desert ecosystem. J. Fungi. 2021, 7, 578. [Google Scholar] [CrossRef]
- Purushotham, N.; Jones, E.; Monk, J.; Ridgway, H. Fungal communities in the native new zealand medicinal plant Pseudowintera colorata (Horopito) are determined by plant organ type and host maturity with key members promoting plant growth. Microorganisms 2021, 9, 2576. [Google Scholar] [CrossRef]
- Li, J.L.; Sun, X.; Zheng, Y.; Lü, P.P.; Wang, Y.L.; Guo, L.D. Diversity and community of culturable endophytic fungi from stems and roots of desert halophytes in northwest China. MycoKeys 2020, 62, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Li, P.; Meng, L.; Xv, K.; Dong, F.; Qiu, Y.; He, L.; Lin, L. Diversity and communities of culturable endophytic fungi from different tree peonies (geoherbs and non-geoherbs), and their biosynthetic potential analysis. Braz. J. Microbiol. 2018, 49, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Silva, J.M.C.; Bates, J.M. Biogeographic patterns and conservation in the South American Cerrado: A tropical Savanna hotspot. BioScience 2002, 52, 225–233. [Google Scholar] [CrossRef]
- Lewis, K.; Barros, F.D.V.; Cure, M.B.; Davies, C.A.; Furtado, M.N.; Hill, T.C.; Hirota, M.; Martins, D.L.; Mazzochini, G.G.; Mitchard, E.T.A.; et al. Mapping native and non-native vegetation in the Brazilian Cerrado using freely available satellite products. Sci. Rep. 2022, 12, 1588. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, O.S.; Marquis, R.J. The Cerrados of Brazil: Ecology and Natural History of a Neotropical Savanna; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]
- Ratter, J.A.; Bridgewater, S.; Ribeiro, J.F. Analysis of the floristic composition of the brazilian Cerrado vegetation III: Comparison of the woody vegetation of 376 areas. J. Bot. 2003, 60, 57–109. [Google Scholar] [CrossRef]
- Souza, R.C.; Mendes, I.C.; Reis-Junior, F.B.; Carvalho, F.M.; Nogueira, M.A.; Vasconcelos, A.T.; Vicente, V.A.; Hungria, M. Shifts in taxonomic and functional microbial diversity with agriculture: How fragile is the Brazilian Cerrado? BMC Microbiol. 2016, 16, 42. [Google Scholar] [CrossRef]
- Dos Santos Lucas, L.; Neto, A.R.; de Moura, J.B.; de Souza, R.F.; Santos, M.E.F.; de Moura, L.F.; Xavier, E.G.; Dos Santos, J.M.; Nehring, R.; Dutra, E.S. Mycorrhizal fungi arbuscular in forage grasses cultivated in Cerrado soil. Sci. Rep. 2022, 12, 3103. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Aluizio, R.; Palácio-Cortes, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and structure of fungal endophyte communities found in the brazilian biomes, Pantanal, and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef]
- Morais, E.M.; Silva, A.A.R.; Sousa, F.W.A.; Azevedo, I.M.B.; Silva, H.F.; Santos, A.M.G.; Beserra Júnior, J.E.A.; Carvalho, C.P.; Eberlin, M.N.; Porcari, A.M.; et al. Endophytic Trichoderma strains isolated from forest species of the Cerrado-Caatinga ecotone are potential biocontrol agents against crop pathogenic fungi. PLoS ONE 2022, 17, e0265824. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D. Endophytic fungal diversity: Review of traditional and molecular techniques. Mycology 2012, 3, 65–76. [Google Scholar]
- Chi, W.C.; Chen, W.; He, C.C.; Guo, S.Y.; Cha, H.J.; Tsang, L.M.; Ho, T.W.; Pang, K.L. A highly diverse fungal community associated with leaves of the mangrove plant Acanthus ilicifolius var. xiamenensis revealed by isolation and metabarcoding analyses. PeerJ 2019, 7, e7293. [Google Scholar] [CrossRef] [PubMed]
- Gautam, A.K.; Verma, R.K.; Avasthi, S.; Sushma; Bohra, Y.; Devadatha, B.; Niranjan, M.; Suwannarach, N. Current insight into traditional and modern methods in fungal diversity estimates. J. Fungi 2022, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.W.K.; Stephenson, S.L.; Bahkali, A.H.; Hyde, K.D. From morphology to molecular biology: Can we use sequence data to identify fungal endophytes? Fungal Divers. 2011, 50, 113–120. [Google Scholar] [CrossRef]
- Durán, M.; San Emeterio, L.; Canals, R.M. comparison of culturing and metabarcoding methods to describe the fungal endophytic assemblage of Brachypodium rupestre growing in a range of anthropized disturbance regimes. Biology 2021, 10, 1246. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.R.D.; Marcuzzo, F.F.N.; Barros, J.R. Climatic classification of Köppen-Geiger for the state of Goiás and the Federal District. ACTA Geog. 2014, 8, 40–55. [Google Scholar] [CrossRef]
- Cavararo, R. Reserva Ecológica do IBGE: Ambiente e plantas vasculares. EstudosandPesquisas. Inf. Geográfica 2004, 3, 1–71. [Google Scholar]
- Eiten, G. Vegetação do Cerrado. In Cerrado: Caracterização, Ocupação Eperspectivas; Universidade de Brasília: Brasília, Brazil, 1994; pp. 17–73. [Google Scholar]
- Eiten, G. The Cerrado vegetation of Brazil. Bot. Rev. 1972, 38, 201–341. [Google Scholar] [CrossRef]
- Silva, F.D.A.; Liotti, R.G.; Boleti, A.P.D.A.; Reis, D.M.; Passos, M.B.S.; dos Santos, E.L.; Sampaio, O.M.; Januário, A.H.; Branco, C.L.B.; da Silva, G.F.; et al. Diversity of cultivable fungal endophytes in Paullinia cupana (Mart.) Ducke and bioactivity of their secondary metabolites. PLoS ONE 2018, 13, e0195874. [Google Scholar] [CrossRef]
- Khan, A.L.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Farsi, Z.; Al-Mamari, A.; Waqas, M.; Asaf, S.; Elyassi, A.; Mabood, F.; Shin, J.-H.; et al. Frankincense tree endophytic fungi enhance host growth and produce extracellular enzymes and indole acetic acid. PLoS ONE 2016, 11, e0158207. [Google Scholar] [CrossRef]
- Dhayanithy, G.; Subban, K.; Chelliah, J. Diversity and biological activities of endophytic fungi associated with Catharanthus roseus. BMC Microbiol. 2019, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Hoog, G.S.; Gerrits van den Ende, A.H. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- DNA Sequence Assembler v4 Heracle BioSoft. 2013. Available online: www.DnaBaser.com (accessed on 7 May 2021).
- Inglis, P.W.; Pappas, M.C.R.; Resende, L.V.; Grattapaglia, D. Fast and inexpensive protocols for consistent extraction of high quality DNA and RNA from challenging plant and fungal samples for high-throughput SNP genotyping and sequencing applications. PLoS ONE 2018, 13, e0206085. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. J. Bioinf. 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. J. Bioinf. 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Otasek, D.; Morris, J.H.; Bouças, J.; Pico, A.R.; Demchak, B. Cytoscape Automation: Empowering workflow-based network analysis. Genome Biol. 2019, 2, 185. [Google Scholar] [CrossRef]
- Philip, D. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005. [Google Scholar]
- Baselga, A.; Orme, C.D.L. Betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Stevens, A.J.; Purcell, R.V.; Darling, K.A.; Eggleston, M.J.F.; Kennedy, M.A.; Rucklidge, J.J. Human gut microbiome changes during a 10 week Randomised Control Trial for micronutrient supplementation in children with attention deficit hyperactivity disorder. Sci. Rep. 2019, 9, 10128. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic. samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Fonseca, P.L.C.; Skaltsas, D.; da Silva, F.F.; Kato, R.B.; de Castro, G.M. An integrative view of the phyllosphere mycobiome of native rubber trees in the brazilian amazon. J. Fungi. 2022, 8, 373. [Google Scholar] [CrossRef]
- Fonseca, P.L.C.; Skaltsas, D.; da Silva, F.F.; Kato, R.B.; de Castro, G.M.; García, G.J.Y.; Quintanilha-Peixoto, G.; Mendes-Pereira, T.; Carmo, A.O.D.; Aguiar, E.R.G.R.; et al. The diversity, antimicrobial and anticancer activity of endophytic fungi associated with the medicinal plant Stryphnodendron adstringens (Mart.) Coville (Fabaceae) from the Brazilian savannah. Symbiosis 2012, 57, 95–107. [Google Scholar]
- Carvalho, C.R.; Gonçalves, V.N.; Pereira, C.B.; Johann, S.; Galliza, I.V.; Alves, T.M.A.; Rabello, A.; Sobral, M.E.G.; Zani, C.; Rosa, C.A.; et al. Diversity of the endophytic fungi associated with the ancient and narrowly endemic neotropical plant Vellozia gigantea from the endangered Brazilian rupestrian grasslands. Biochem. Syst. Ecol. 2017, 71, 163–169. [Google Scholar]
- Li, P.; Wu, Z.; Liu, T.; Wang, Y. Biodiversity, phylogeny, and antifungal functions of endophytic fungi associated with Zanthoxylum bungeanum. Int. J. Mol. Sci. 2016, 17, 1541. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Chen, X.; Luo, X.; Zhang, H.; Liu, Y.; Zhang, Y.; Wu, J.; Zhao, C.; Zhao, P. Diversity of endophytic fungi from the leaves of Vaccinium dunalianum. Lett. Appl. Microbiol. 2020, 71, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [PubMed]
- Yahr, R.; Schoch, C.L.; Dentinger, B.T. Scaling up discovery of hidden diversity in fungi: Impacts of barcoding approaches. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150336. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef]
- Dos Reis, J.B.A.; Lorenzi, A.S.; do Vale, H.M.M. Methods used for the study of endophytic fungi: A review on methodologies and challenges, and associated tips. Arch. Microbiol. 2022, 204, 675. [Google Scholar] [CrossRef]
- Dos Reis, J.B.A.; do Vale, H.M.M.; Lorenzi, A.S. Insights into taxonomic diversity and bioprospecting potential of Cerrado endophytic fungi: A review exploring an unique Brazilian biome and methodological limitations. World J. Microbiol. Biotechnol. 2022, 38, 202. [Google Scholar] [CrossRef]
- Yang, J.H.; Oh, S.Y.; Kim, W.; Hur, J.S. Endolichenic fungal community analysis by pure culture isolation and metabarcoding: A case study of Parmotrema tinctorum. Mycobiology 2022, 50, 55–65. [Google Scholar] [CrossRef]
- Nischitha, R.; Shivanna, M.B. Antimicrobial activity and metabolite profiling of endophytic fungi in Digitaria bicornis (Lam) Roem and Schult. and Paspalidium flavidum (Retz.) A. Camus. 3 Biotech 2021, 11, 53. [Google Scholar] [CrossRef]
- Parungao, M.M.; Fryar, S.C.; Hyde, K.D. Diversity of fungi on rainforest litter in North Queensland, Australia. Biodivers. Conserv. 2002, 11, 1185–1194. [Google Scholar] [CrossRef]
- Liu, J.; Ridgway, H.J.; Jones, E.E. Apple endophyte community is shaped by tissue type, cultivar and site and has members with biocontrol potential against Neonectria ditissima. J. Appl. Microbiol. 2020, 128, 1735–1753. [Google Scholar] [CrossRef] [PubMed]
- Daghino, S.; Martino, E.; Voyron, S.; Perotto, S. Metabarcoding of fungal assemblages in Vaccinium myrtillus endosphere suggests colonization of above-ground organs by some ericoid mycorrhizal and DSE fungi. Sci. Rep. 2022, 12, 11013. [Google Scholar] [CrossRef] [PubMed]
- Marczylo, E.L.; Macchiarulo, S.; Gant, T.W. Metabarcoding of soil fungi from different urban greenspaces around Bournemouth in the UK. Ecohealth 2021, 18, 315–330. [Google Scholar] [CrossRef]
- Alem, D.; Dejene, T.; Geml, J.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Metabarcoding analysis of the soil fungal community to aid the conservation of underexplored church forests in Ethiopia. Sci. Rep. 2022, 12, 4817. [Google Scholar] [CrossRef] [PubMed]
- Cowan, D.; Lebre, P.; Amon, C.; Becker, R.; Boga, H.; Boulangé, A.; Chiyaka, T.; Coetzee, T.; de Jager, P.; Dikinya, O.; et al. Biogeographical survey of soil microbiomes across sub-Saharan Africa: Structure, drivers, and predicted climate-driven changes. Microbiome 2022, 10, 131. [Google Scholar] [CrossRef]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Stürmer, S.L.; Kemmelmeier, K. The Glomeromycota in the neotropics. Front. Microbiol. 2021, 12, 553679. [Google Scholar] [CrossRef]
- Burgdorf, R.J.; Laing, M.D.; Morris, C.D.; Jamal-Ally, S.F. A procedure to evaluate the efficiency of surface sterilization methods in culture-independent fungal endophyte studies. Braz. J. Microbiol. 2014, 45, 977–983. [Google Scholar] [CrossRef]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. Fungal Traits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, T.P.; Li, L.Z.; Luo, M.Y.; Jia, W.Y.; Zeng, Y.; Jiang, Y.L.; Tao, G.C. Multigene phylogeny, diversity and antimicrobial potential of endophytic sordariomycetes from Rosa roxburghii. Front Microbiol. 2021, 12, 755919. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 53, 1–221. [Google Scholar] [CrossRef] [PubMed]
- Guatimosim, E.; Firmino, A.L.; Bezerra, J.L.; Pereira, O.L.; Barreto, R.W.; Crous, P.W. Towards a phylogenetic reappraisal of Parmulariaceae and Asterinaceae (Dothideomycetes). Persoonia 2015, 35, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, I.C.; Crous, P.W.; Groenewald, J.Z.; Maharachchikumbura, S.S.N.; Jeewon, R.; Phillips, A.J.L.; Bhat, J.D.; Perera, R.H.; Li, Q.R.; Li, W.J.; et al. Families of Diaporthales based on morphological and phylogenetic evidence. Stud. Mycol. 2017, 86, 217–296. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee, M.P.A.; Wingfield, M.J.; Crous, P.W. Evolution of lifestyles in Capnodiales. Stud. Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef] [PubMed]
- Martorelli, I.; Helwerda, L.S.; Kerkvliet, J.; Gomes, S.I.F.; Nuytinck, J.; van der Werff, C.R.A.; Ramackers, G.J.; Gultyaev, A.P.; Merckx, V.S.F.T.; Verbeek, F.J. Fungal metabarcoding data integration framework for the MycoDiversity DataBase (MDDB). J. Integr. Bioinform. 2020, 17, 20190046. [Google Scholar] [CrossRef]
- Vargas-Gastélum, L.; Riquelme, M. The Mycobiota of the deep sea: What omics can offer. Life 2020, 10, 292. [Google Scholar] [CrossRef]
- Pang, B.; Yin, D.; Zhai, Y.; He, A.; Qiu, L.; Liu, Q.; Ma, N.; Shen, H.; Jia, Q.; Liang, Z.; et al. Diversity of endophytic fungal community in Huperzia serrata from different ecological areas and their correlation with Hup A content. BMC Microbiol. 2022, 22, 191. [Google Scholar] [CrossRef]
- Pang, K.L.; Guo, S.Y.; Chen, I.A.; Burgaud, G.; Luo, Z.H.; Dahms, H.U.; Hwang, J.S.; Lin, Y.L.; Huang, J.S.; Ho, T.W.; et al. Insights into fungal diversity of a shallow-water hydrothermal vent field at Kueishan Island, Taiwan by culture-based and metabarcoding analyses. PLoS ONE 2019, 14, e0226616. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N (dag/Kg) | P (dag/Kg) | K (dag/Kg) | Ca (dag/Kg) | Mg (dag/Kg) | S (dag/Kg) | Al (dag/Kg) | SLA (cm2/g) |
|---|---|---|---|---|---|---|---|---|
| Caryocar brasiliense | 1.25 ± 0.06 b | 0.06 ± 0.00 a | 0.30 ± 0.04 ab | 0.36 ± 0.08 c | 0.18 ± 0.04 bd | 0.08 ± 0.00 ab | 280.34 ± 44.11 a | 66.49 ± 5.84 bc |
| Dalbergia miscolobium | 1.95 ± 0.09 a | 0.06 ± 0.00 a | 0.29 ± 0.02 a | 0.18 ± 0.03 a | 0.14 ± 0.02 ab | 0.08 ± 0.00 ab | 113.00 ± 27.15 a | 94.04 ± 15.43 a |
| Leptolobium dasycarpum | 1.78 ± 0.14 a | 0.06 ± 0.00 a | 0.32 ± 0.05 ab | 0.17 ± 0.04 a | 0.08 ± 0.03 a | 0.07 ± 0.01 a | 172.66 ± 41.66 a | 63.26 ± 12.48 bc |
| Qualea parviflora | 1.27 ± 0.10 b | 0.06 ± 0.00 ab | 0.34 ± 0.05 ab | 0.32 ± 0.02 ac | 0.17 ± 0.02 bd | 0.08 ± 0.01 ab | 12,345.41 ± 1001.30 b | 85.06 ± 14.09 ac |
| Ouratea hexasperma | 1.17 ± 0.15 b | 0.05 ± 0.00 b | 0.38 ± 0.06 b | 0.66 ± 0.14 b | 0.25 ± 0.03 c | 0.08 ± 0.01 ab | 106.08 ± 27.07 a | 56.34 ± 10.52 b |
| Styrax ferrugineus | 1.33 ± 0.09 b | 0.06 ± 0.00 a | 0.27 ± 0.02 a | 0.40 ± 0.12 c | 0.21 ± 0.06 cd | 0.09 ± 0.00 b | 242.27 ± 61.44 a | 63.16 ± 10.26 bc |
| Host Species | Isolate ID | Closest GenBank Match | Match | Identity (%) | Query Coverage (%) |
|---|---|---|---|---|---|
| Caryocar brasiliense | CB1629_2 CB1629_22 CB1629_25 CB1629_3 CB1629_31 CB995_15 CB995_2 CB995_22 CB995_3 CB995_6 CB995_7 | Diaporthe sp. Diaporthe schini Diaporthe sp. Diaporthe sp. Didymella keratinophila Diaporthe rosae Diaporthe sp. Diaporthe sp. Diaporthe sp. Diaporthe actinidiae Diaporthe masirevicii | NR_111849.1 NR_111861.1 NR_147537.1 NR_145303.1 NR_158275.1 MG828894.1 EU552122.1 MH171064.1 NR_137105.1 KC145886.1 NR_147534.1 | 95% 99% 95% 94% 99% 99% 95% 96% 92% 99% 98% | 88% 100% 86% 95% 100% 100% 100% 99% 100% 100% 100% |
| Dalbergia miscolobium | DM1044_2 DM1044_23 DM1044_3 DM1044_6 DM1044_16 DM1044_8 DM1044_19 DM1044_9 DM1044_5 DM18_1 DM18_12 DM18_2 DM18_8 DM2226_1 DM2226_10 DM2226_11 DM2226_11_1 DM2226_13 DM2226_14 DM2226_15 DM2226_19 DM2226_22 DM2226_29 DM2226_3 DM2226_4 DM2226_17 DM2226_5 DM2226_6 DM18_6 | Colletotrichum spicati Didymella chlamydospora Diaporthe sp. Alternaria indefessa Diaporthe masirevicii Didymella keratinophila Diaporthe sp. Diaporthe raonikayaporum Didymella keratinophila Epicoccum sorghicola Diaporthe sp. Epicoccum sorghicola Diaporthe sp. Diaporthe sp. Diaporthe parapterocarpi Ascochyta phacae Diaporthe velutina Colletotrichum sp. Seiridium marginatum Colletotrichum chrysophilum Colletotrichum philoxeroidis Stenocarpella sp. Diaporthe sp. Ascochyta rabiei Diaporthe sp. Ascochyta phacae Colletotrichum brevisporum Epicoccum sorghicola Didymella keratinophila | OL842171.1 MK836111.1 MH855768.1 MH861641.1 KJ197276.1 NR_158275.1 EU552122.1 NR_111860.1 NR_158275.1 OK442368.1 MH864501.1 OK442368.1 MN152927.1 MH855768.1 NR_168152.1 MH857437.1 NR_152470.1 MK541032.1 KT949914.1 NR_160821.1 OL842188.1 NR_173403.1 NR_168240.1 EU167600.1 EU552122.1 MH857437.1 KC790943.1 OK442368.1 NR_158275.1 | 99% 98% 95% 99% 98% 99% 93% 99% 99% 98% 93% 98% 93% 95% 99% 98% 97% 92% 98% 99% 97% 93% 94% 99% 90% 98% 98% 99% 99% | 96% 100% 100% 100% 100% 100% 100% 100% 100% 100% 100% 100% 99% 100% 100% 99% 100% 100% 99% 100% 96% 100% 92% 99% 100% 99% 95% 100% 100% |
| Leptolobium dsycarpum | LD2293_12 LD2293_13 LD2293_23 LD2293_3 LD2293_4 LD2293_7 LD2299_1 LD2299_2 LD2299_3 LD2299_35 LD2299_4 LD2299_5 LD2299_6 LD2304_1 LD2304_2 LD2304_30 LD2304_34 LD2304_35 LD2304_50 | Diaporthe stewartii Diaporthe stewartii Diaporthe cynaroidis Diaporthe terebinthifolii Diaporthe sp. Microdiplodia sp. Diaporthe macadamiae Diaporthe terebinthifolii Diaporthe ocoteae Diaporthe macadamiae Diaporthe sp. Diaporthe rosiphthora Diaporthe sp. Diaporthe sp. Diaporthe foeniculina Neopestalotiopsis dendrobii Diaporthe macintoshii Diaporthe macintoshii Diaporthe inconspicua | MH855768.1 MH855768.1 EU552122.1 NR_111862.1 NR_147596.1 DQ885897.1 NR_168240.1 NR_111862.1 NR_147596.1 NR_168240.1 NR_111849.1 MT311197.1 MH171064.1 NR_168240.1 NR_145303.1 MK993572.1 NR_147539.1 NR_147539.1 NR_111849.1 | 99% 96% 96% 99% 93% 96% 98% 99% 98% 98% 90% 99% 96% 95% 97% 99% 98% 98% 99% | 100% 100% 100% 99% 100% 100% 98% 100% 100% 100% 99% 100% 100% 96% 100% 100% 99% 100% 99% |
| Ouratea hexasperma | OH1078_22 OH1078_12 OH1078_2 OH1078_3 OH1078_31 OH1078_24 OH2206_31 OH2208_3 OH2208_4 OH2262_2 OH2262_30 OH2262_31 OH2262_4 OH2262_40 OH2262_5 OH2262_6 | Kalmusia erioi Phyllosticta catimbauensis Diaporthe novem Diaporthe sp. Kalmusia erioi Dendrothyrium sp. Phyllosticta fallopiae Stilbospora sp. Phyllosticta capitalensis Diaporthe maytenicola Diaporthe sp. Epicoccum sorghicola Coniochaeta sp. Diaporthe sp. Diaporthe sp. Diaporthe sp. | MN473058.1 NR_156631.1 MH864503.1 NR_120138.1 MN473058.1 JX496097.1 AB454307.1 KF570166.1 OL957169.1 NR_137826.1 EU552122.1 OK442368.1 NR_173009.1 NR_147596.1 NR_158416.1 MH855768.1 | 99% 99% 98% 93% 97% 93% 100% 88% 100% 97% 92% 98% 90% 93% 93% 95% | 100% 88% 100% 97% 97% 98% 99% 100% 100% 100% 100% 100% 99% 100% 100% 100% |
| Qualea parviflora | QP1131_11 QP1646_1 QP1646_11 QP1646_13 QP1646_3 QP1646_4 QP65_0 QP65_30 QP65_4 QP971_2 | Pestalotiopsis protearum Diaporthe macintoshii Neopestalotiopsis dendrobii Diaporthe sp. Diaporthe inconspicua Diaporthe inconspicua Kalmusia erioi Cytospora viridistroma Cytospora viridistroma Clathrosporium sp. | JN712498.1 NR_147539.1 MK993572.1 NR_147539.1 NR_111849.1 NR_111849.1 MN473058.1 MN172408.1 MN172408.1 NR_153908.1 | 99% 98% 99% 96% 99% 99% 97% 99% 99% 88% | 100% 100% 100% 100% 100% 100% 97% 100% 100% 86% |
| Styrax ferrugineus | SF1034_3 SF1034_4 SF134_2 SF134_22 SF134_25 SF134_3 SF134_3_1 SF134_4 SF134_6 SF134_7 SF1697_10 SF1697_16 SF1697_17 SF1697_3 SF1697_13 SF1697_7 SF1697_8_1 SF1697_8 SF1697_15 SF1756_12 SF1756_13_1 SF1756_18 SF1756_2 SF1756_3 SF1756_4 SF1756_5 SF1756_6 SF1756_8 SF1756_9 | Cytospora viridistroma Diaporthe sp. Diaporthe macintoshii Diaporthe macintoshii Diaporthe foeniculina Hymenopleella sp. Diaporthe sp. Diaporthe ilicicola Monochaetia sp. Monochaetia sp. Diaporthe novem Diaporthe sp. Diaporthe stewartii Lasiodiplodia parva Cytospora sp. Melanconis sp. Diaporthe sp. Diaporthe terebinthifolii Diaporthe sp. Diaporthe baccae Diaporthe ilicicola Diaporthe sp. Diaporthe sp. Diaporthe ilicicola Diaporthe sp. Diaporthe sp. Diaporthe ilicicola Diaporthe sp. Diaporthe terebinthifolii | MN172408.1 NR_111857.1 NR_147539.1 NR_147539.1 NR_145303.1 KT949901.1 NR_137825.1 MH171064.1 LC146750.1 LC146750.1 MH864503.1 EU552122.1 MH855768.1 MH861166.1 MK912135.1 MN784964.1 MH855768.1 NR_111862.1 MH855768.1 NR_152458.1 MH171064.1 NR_172435.1 MH171064.1 MH171064.1 MH171064.1 NR_137126.1 MH171064.1 NR_111849.1 NR_111862.1 | 99% 93% 98% 98% 97% 90% 96% 98% 95% 95% 99% 90% 98% 99% 98% 90% 95% 99% 96% 97% 97% 96% 95% 97% 95% 96% 97% 95% 99% | 100% 100% 100% 100% 100% 100% 100% 99% 90% 90% 100% 100% 100% 100% 100% 100% 100% 100% 100% 100% 92% 100% 89% 99% 100% 100% 99% 90% 100% |
| Plant Host | Read | OTUs | Phyla | Classes | Orders | Families | Genera |
|---|---|---|---|---|---|---|---|
| Caryocar brasiliense | 343,535 | 2050 | 8 | 27 | 64 | 162 | 309 |
| Dalbergia miscolobium | 35,782 | 173 | 3 | 13 | 30 | 52 | 68 |
| Leptolobium dasycarpum | 417,491 | 859 | 7 | 25 | 64 | 131 | 169 |
| Ouratea hexasperma | 314,104 | 1113 | 9 | 22 | 58 | 142 | 243 |
| Qualea parviflora | 427,863 | 395 | 5 | 11 | 32 | 75 | 116 |
| Styrax ferrugineus | 338,354 | 337 | 4 | 14 | 42 | 87 | 114 |
| Total | 1,877,129 | 3821 | 9 | 35 | 86 | 214 | 435 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Reis, J.B.A.; Pappas Junior, G.J.; Lorenzi, A.S.; Pinho, D.B.; Costa, A.M.; Bustamante, M.M.d.C.; Vale, H.M.M.d. How Deep Can the Endophytic Mycobiome Go? A Case Study on Six Woody Species from the Brazilian Cerrado. J. Fungi 2023, 9, 508. https://doi.org/10.3390/jof9050508
dos Reis JBA, Pappas Junior GJ, Lorenzi AS, Pinho DB, Costa AM, Bustamante MMdC, Vale HMMd. How Deep Can the Endophytic Mycobiome Go? A Case Study on Six Woody Species from the Brazilian Cerrado. Journal of Fungi. 2023; 9(5):508. https://doi.org/10.3390/jof9050508
Chicago/Turabian Styledos Reis, Jefferson Brendon Almeida, Georgios Joannis Pappas Junior, Adriana Sturion Lorenzi, Danilo Batista Pinho, Alexandra Martins Costa, Mercedes Maria da Cunha Bustamante, and Helson Mario Martins do Vale. 2023. "How Deep Can the Endophytic Mycobiome Go? A Case Study on Six Woody Species from the Brazilian Cerrado" Journal of Fungi 9, no. 5: 508. https://doi.org/10.3390/jof9050508