
Three New Species of Fusicolla (Hypocreales) from China

State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing 100101, China
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(5), 572; https://doi.org/10.3390/jof9050572
Submission received: 8 April 2023
/
Revised: 11 May 2023
/
Accepted: 13 May 2023
/
Published: 15 May 2023
(This article belongs to the Special Issue Ascomycota: Diversity, Taxonomy and Phylogeny 2.0)
Abstract
:To explore the species diversity of the genus Fusicolla, specimens from Henan, Hubei and Jiangsu Provinces in China are examined, and three undescribed taxa are encountered. The morphological characteristics and DNA sequence analyses of the combined acl1, ITS, LSU, rpb2 and tub2 regions support their placement in Fusicolla and their recognition as new species. Fusicolla aeria sp. nov. is distinguished by the formation of abundant aerial mycelia on PDA, falcate, (1–)3-septate macroconidia 16–35 × 1.5–2.8 μm and subcylindrical, aseptate microconidia 7.5–13 × 0.8–1.1 μm. Fusicolla coralloidea sp. nov. has a coralloid colony on PDA, falcate, 2–5-septate macroconidia 38–70 × 2–4.5 μm and rod-shaped to ellipisoidal, aseptate microconidia 2–7 × 1–1.9 μm. Fusicolla filiformis sp. nov. is characterized by filiform, 2–6-septate macroconidia 28–58 × 1.5–2.3 μm and lacking microconidia. Morphological differences between these novel species and their close relatives are compared in detail. The previously recorded species of the genus in China are listed and a key to these taxa is provided.
1. Introduction
The genus Fusicolla Bonord., typified by F. betae (Desm.) Bonord., was established by Bonorden [1] and redefined by Gräfenhan et al. [2], who raised the varieties of Fusarium aquaeductuum (Radlk. & Rabenh.) Lagerh. & Rabenh. to species rank and transferred Fusarium merismoides Corda to Fusicolla. The genus is characterized by scattered to gregarious, yellow, pale buff to orange, globose to pyriform perithecia that are fully or partially immersed in stromata; cylindrical to narrowly clavate asci containing eight ascospores; and the production of falcate, straight to curved, 1–5-septate macroconidia [2,3]. They are mostly saprobes and occur on various substrata, such as rotten twigs, decayed wood, the stromata of other fungi, soil, water, the slime flux of trees, sewage, the bones of wild boar and even air [2,4,5,6,7,8,9]. Currently, there are 22 species accepted in this genus [9,10], of which five are reported from China [9,11,12,13].
Within the scope of our current study on the Chinese Fungus Flora, fresh hypocrealean specimens are examined. Based on the morphology and phylogenetic analyses of the combined sequences of the larger subunit of the ATP citrate lyase (acl1), nuclear ribosomal DNA ITS1-5.8S-ITS2 (ITS), the large subunit of nuclear ribosomal DNA (LSU), the second largest subunit of RNA polymerase II (rpb2) and β-tubulin (tub2), three novel species of Fusicolla are introduced. Comparisons between these taxa and their close relatives are performed. The previously recorded Fusicolla species in China are also listed.
2. Materials and Methods
2.1. Sampling and Morphological Studies
Specimens on wood substrates were collected from Henan, Hubei and Jiangsu Provinces in China and deposited in the Herbarium Mycologicum Academiae Sinicae (HMAS). Lactophenol cotton blue solution was used as a mounting medium for the examination of features and measurements of conidiophores, macroconidia and microconidia. Photographs were taken with a Zeiss AxioCam MRc 5 digital camera (Jena, Germany) attached to a Zeiss Axio Imager A2 microscope (Göttingen, Germany). Cultures were deposited in the China General Microbiological Culture Collection Center (CGMCC). For colony features and growth rates, strains were grown on potato dextrose agar (PDA, 20% (w/v) potato + 2% (w/v) dextrose + 2% (w/v) agar) and synthetic nutrient-poor agar (SNA) [14] in 90 mm plastic Petri dishes at 25 °C for 14 d with alternating periods of light and darkness (12 h/12 h).
2.2. DNA Extraction, PCR Amplification, Sequencing and Phylogenetic Analyses
Genomic DNA was extracted from fresh mycelium following the method of Wang and Zhuang [15]. Five primer pairs, acl1-230up/acl1-1220low [16], ITS5/ITS4 [17], LR0R/LR5 [18,19], RPB2-5f/RPB2-7cR [20] and T1/T22 [21], were used to amplify the sequences of the acl1, ITS, LSU, rpb2 and tub2 regions, respectively. PCR reactions were performed using an ABI 2720 Thermal Cycler (Applied Biosciences, Foster City, CA, USA) with a 25 μL reaction mixture consisting of 12.5 μL Taq MasterMix, 1 μL of each primer (10 μM), 1 μL template DNA and 9.5 μL ddH2O. DNA sequencing was carried out in both directions on an ABI 3730XL DNA Sequencer (Applied Biosciences, Foster City, CA, USA).
Newly acquired sequences and those retrieved from GenBank are listed in Table 1. The sequences were assembled and aligned, and the primer sequences were trimmed by BioEdit 7.0.5 [22] and converted to NEXUS files by ClustalX 1.83 [23]. The sequences of acl1, ITS, LSU, rpb2 and tub2 were combined and analyzed by Bayesian inference (BI) and maximum likelihood (ML) methods to determine the phylogenetic positions of these strains. The BI analysis was conducted by MrBayes 3.1.2 [24] using a Markov chain Monte Carlo (MCMC) algorithm. Nucleotide substitution models were determined by MrModeltest 2.3 [25]. The ML analysis was performed via IQ-Tree 1.6.12 [26] using the best model for each locus chosen by ModelFinder [27]. Trees were examined by TreeView 1.6.6 [28]. The Bayesian inference posterior probability (BIPP) values greater than 0.9 and maximum likelihood bootstrap (MLBP) values greater than 70% were shown at the nodes.
3. Results
3.1. Phylogeny
The acl1, ITS, LSU, rpb2 and tub2 sequences of 24 Fusicolla species were analyzed. The resulting BI tree is shown in Figure 1. The topology of the ML tree was similar to that of the BI tree. The final matrix was deposited in TreeBASE with accession no. S30023. The isolates CGMCC 3.24907, 3.24908, 3.24909 and 3.24910 grouped with other members of Fusicolla, and the genus received high statistical support values (BIBP/MLBP = 1.0/96%). The isolate CGMCC 3.24910 clustered together with F. gigas Chang Liu, Z.Q. Zeng & W.Y. Zhuang (BIBP/MLBP = 1.0/100%). The isolates CGMCC 3.24908 and 3.24909 were related to F. acetilerea (Tubaki, C. Booth & T. Harada) Gräfenhan & Seifert and F. elongata Decock, Crous & Sand.-Den. but with low support values, and the isolate CGMCC 3.24907 formed a separate lineage.
3.2. Taxonomy
- Fusicolla aeria Z.Q. Zeng & W.Y. Zhuang, sp. nov., Figure 2.
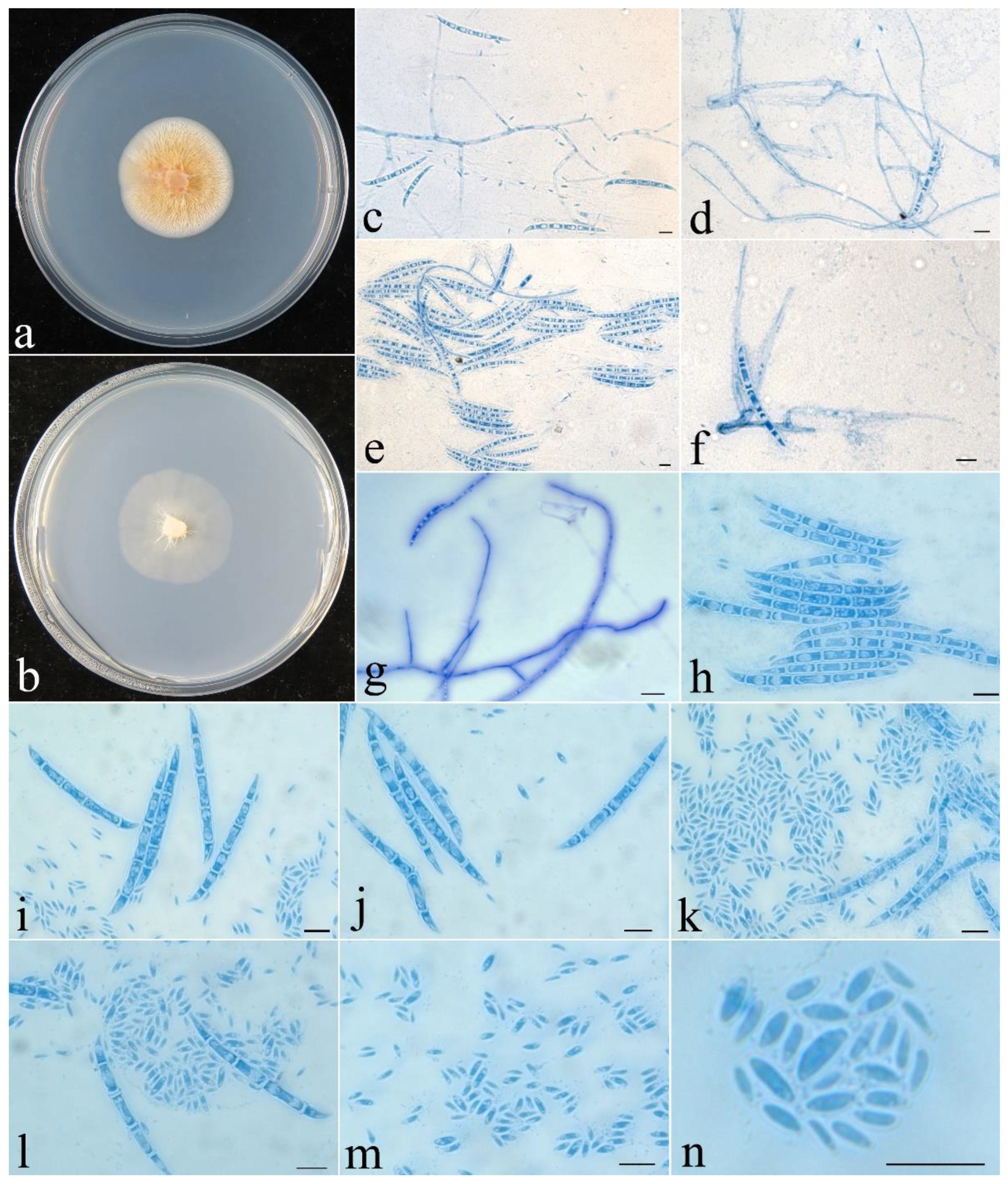
Figure 2.
Fusicolla aeria (CGMCC 3.24908). (a) Colony after 2 wk at 25 °C on PDA; (b) colony after 2 wk at 25 °C on SNA; (c–e) conidiophores and macroconidia; (f–i) macroconidia; (j) conidiophores and microconidia; (k–n) microconidia. Bars: 10 μm.
Figure 2.
Fusicolla aeria (CGMCC 3.24908). (a) Colony after 2 wk at 25 °C on PDA; (b) colony after 2 wk at 25 °C on SNA; (c–e) conidiophores and macroconidia; (f–i) macroconidia; (j) conidiophores and microconidia; (k–n) microconidia. Bars: 10 μm.

Fungal Names: FN 571312.
Etymology: The specific epithet refers to the abundant aerial mycelium on PDA.
Typification: CHINA, Henan Province, Luoyang, Yushan Forest Park, 34°41′23″ N 112°6′13″ E, on rotten twig, 23 September 2013, H.D. Zheng, Z.Q Zeng & Z.X. Zhu 8875 (holotype HMAS 247866, ex-type strain CGMCC 3.24908). Sequences: acl1 OQ134105, ITS OQ128334, LSU OQ128338, rpb2 OQ134111, tub2 OQ134100.
Other specimen examined: CHINA, Henan Province, Jiaozuo, Yuntaishan, 35°25′53″ N 113°23′30″ E, on twig associated with other fungi, 25 September 2013, H.D. Zheng, Z.Q. Zeng & Z.X. Zhu 8916a (HMAS 247867, strain CGMCC 3.24909). Sequences: acl1 OQ134106, ITS OQ128335, LSU OQ128339, rpb2 OQ134112, tub2 OQ134101.
Colony characteristics: On PDA 35 mm diam. after 2 wk at 25 °C, with abundant, orange aerial mycelium, producing pinkish orange pigment. On SNA 40 mm diam. after 2 wk at 25 °C, with sparse, pale greyish-white aerial mycelium. Conidiophores unbranched or simple branched, hyaline, smooth-walled, septate, bearing terminal and lateral conidiogenous cells. Conidiogenous cells monophialidic, cylindrical to conical, 18−40 × 1.5−3 µm, smooth, thin-walled. Macroconidia falcate, straight to slightly curved, slightly hooked at one end, hyaline, smooth, (1–)3-septate, 16–35 × 1.5–2.8 μm. Microconidia aseptate, subcylindrical, curved to C-shaped, smooth, hyaline, 7.5–13 × 0.8–1.1 μm. Chlamydospores absent. Sexual stage not observed.
Notes: Among the known species of the genus, F. aeria is distinct because of its abundant aerial mycelium on PDA. Morphologically, it resembles F. gigas and F. matuoi (Hosoya & Tubaki) Gräfenhan & Seifert in having C-shaped microconidia in culture. However, F. gigas possesses larger macroconidia (32−80 × 2.3–3.8 µm) with more septa (3–9 septa) [9], while F. matuoi forms longer macroconidia (17–56 μm long) [29]. Phylogenetically, they are remotely related (Figure 1).
- Fusicolla coralloidea Z.Q. Zeng & W.Y. Zhuang, sp. nov., Figure 3.
Figure 3.
Fusicolla coralloidea (CGMCC 3.24907). (a) Colony after 2 wk at 25 °C on PDA; (b) colony after 2 wk at 25 °C on SNA; (c–e) conidiophores, macroconidia and microconidia; (f,g) conidiophores and macroconidia; (h–l) macroconidia and microconidia; (m,n) microconidia. Bars: 10 μm.
Figure 3.
Fusicolla coralloidea (CGMCC 3.24907). (a) Colony after 2 wk at 25 °C on PDA; (b) colony after 2 wk at 25 °C on SNA; (c–e) conidiophores, macroconidia and microconidia; (f,g) conidiophores and macroconidia; (h–l) macroconidia and microconidia; (m,n) microconidia. Bars: 10 μm.

Fungal Names: FN 571313.
Etymology: The specific epithet refers to the coralloid colony on PDA.
Typification: CHINA, Jiangsu Province, Nanjing City, campus of Nanjing Normal University, 32°6′44″ N 118°55′ E, on twig associated with other fungi, 25 July 2011, Z.Q. Zeng & H.D. Zheng 7895 (holotype HMAS 247870, ex-type strain CGMCC 3.24907). Sequences: acl1 OQ134104, ITS OQ128333, LSU OQ128337, rpb2 OQ134110, tub2 OQ134099.
Colony characteristics: On PDA 34 mm diam. after 2 wk at 25 °C, forming coralloid synnema on surface, producing pale orange-yellow pigment. On SNA 32 mm diam. after 2 wk at 25 °C, with very sparse, pale greyish-white aerial mycelium. Conidiophores arising from somatic hyphae, hyaline, smooth-walled, septate, bearing terminal and lateral conidiogenous cells. Conidiogenous cells monophialidic, cylindrical to conical, 18−60 × 2−3 µm, smooth, thin-walled. Macroconidia falcate, straight to slightly curved, acute at both ends, slightly hooked at one end, hyaline, smooth, 2–5-septate, 38–70 × 2–4.5 μm. Microconidia aseptate, rod-shaped to ellipisoidal, straight to slightly curved, hyaline, smooth, 2–7 × 1–1.9 μm. Chlamydospores absent. Sexual stage not observed.
Note: Among the known species of Fusicolla, F. coralloidea is distinguished by the production of coralloid synnemata on the PDA surface. The fungus resembles F. epistroma (Höhn.) Gräfenhan & Seifert in having rod-shaped to ellipisoidal microconidia [2]. However, the microconidia of the latter are much longer (3.5–8 μm long). Phylogenetically, they were recognized as distinct lineages (Figure 1). Both morphology and DNA sequence analyses support the independent status of these species.
- Fusicolla filiformis Z.Q. Zeng & W.Y. Zhuang, sp. nov., Figure 4.
Figure 4.
Fusicolla filiformis (CGMCC 3.24910). (a) Colony after 2 wk at 25 °C on PDA; (b) colony after 2 wk at 25 °C on SNA; (c,d) conidiophores and macroconidia; (e–p) macroconidia. Bars: 10 μm.
Figure 4.
Fusicolla filiformis (CGMCC 3.24910). (a) Colony after 2 wk at 25 °C on PDA; (b) colony after 2 wk at 25 °C on SNA; (c,d) conidiophores and macroconidia; (e–p) macroconidia. Bars: 10 μm.

Fungal Names: FN 571314.
Etymology: The specific epithet refers to its filiform macroconidia.
Typification: CHINA, Hubei Province, Shennongjia Forestry District, Muyu Town, 31°24′55″ N 110°28′55″ E, on rotten twig, 25 October 2021, Z.Q. Zeng, Z.H. Yu & J.X. Deng 12994b (holotype HMAS 247871, ex-type strain CGMCC 3.24910). Sequences: acl1 OQ134103, ITS OQ128332, LSU OQ128336, rpb2 OQ134109, tub2 OQ134098.
Colony characteristics: On PDA 20 mm diam. after 2 wk at 25 °C, with very sparse, orange aerial mycelium, producing orange pigment. On SNA 17 mm diam. after 2 wk at 25 °C, with very sparse, pale yellowish-white aerial mycelium. Conidiophores arising from somatic hyphae, unbranched, hyaline, smooth-walled, septate, bearing terminal and lateral conidiogenous cells. Conidiogenous cells monophialidic, cylindrical to conical, 25–62 × 1.8–2.5 μm, smooth, thin-walled. Macroconidia filiform to falcate, straight to slightly curved, acute at both ends, with hooked cell at one end, hyaline, smooth, 2–6-septate, 28–58 × 1.5–2.3 μm. Microconidia and chlamydospores absent. Sexual stage not observed.
Note: Phylogenetically, F. filiformis clustered with F. gigas, receiving full support (Figure 1). However, between their type cultures, there are 25 bp, 7 bp, 9 bp, 39 bp and 21 bp divergences detected for acl1, ITS, LSU, rpb2 and tub2 regions, respectively. Morphologically, F. gigas differs in having C-shaped microconidia and wider macroconidia (2.5–3.5 μm wide) with more septa (up to nine septa) [9].
- Other Fusicolla Species Recorded in China
- Fusicolla aquaeductuum (Radlk. & Rabenh.) Gräfenhan, Seifert & Schroers, in Gräfenhan, Schroers, Nirenberg & Seifert, Stud. Mycol. 68: 100, 2011.
≡ Selenosporium aquaeductuum Radlk. & Rabenh., in Rabenhorst, Hedwigia 2: 73, 1862.
≡ Fusarium aquaeductuum (Radlk. & Rabenh.) Lagerh. & Rabenh., Centbl. Bakt. ParasitKde, Abt. I 9: 655. 1891.
- Fusicolla gigas Chang Liu, Z.Q. Zeng & W.Y. Zhuang, in Crous et al., Fungal Systematics and Evolution 9: 192, 2022.
Specimen examined: CHINA, Chongqing City, Wushan County, Hongchiba National Forest Park, in soil, 30 October 2020, Z.Q. Zeng, X.C. Wang, H.D. Zheng & C. Liu CGMCC 3.20680 (HMAS 247872).
- Fusicolla guangxiensis Z.Q. Zeng, Chang Liu & W.Y. Zhuang, in Crous et al., Fungal Systematics and Evolution 9: 192, 2022.
Specimen examined: CHINA, Guangxi Zhuang Autonomous Region, Fangchenggang City, Shiwandashan National Forest Park, on rotten twig, 10 December 2019, Z.Q. Zeng & H.D. Zheng CGMCC 3.20679 (HMAS 247873).
- Fusicolla matuoi (Hosoya & Tubaki) Gräfenhan & Seifert, in Gräfenhan, Schroers, Nirenberg & Seifert, Stud. Mycol. 68: 101, 2011.
≡ Fusarium matuoi Hosoya & Tubaki, Mycoscience 45: 264, 2004.
- Fusicolla violacea Gräfenhan & Seifert, in Gräfenhan, Schroers, Nirenberg & Seifert, Stud. Mycol. 68: 101, 2011.
= Fusarium merismoides var. violaceum Gerlach, Phytopath. Z. 90(1): 34, 1977. Nom. inval., Art. 37.
- Key to the Known Species of Fusicolla in China
| 1. Forming macroconidia and microconidia on PDA | 2 |
| 1. Only forming macroconidia on PDA | 6 |
| 2. Microconidia ellipiosoid, rod-shaped to falcate | 3 |
| 2. Microconidia subcylindrical, curved to C-shaped | 4 |
| 3. Producing pale orange-yellow pigment on PDA | F. coralloidea |
| 3. Producing purple pigment on PDA | F. violacea |
| 4. Aerial mycelium abundant on PDA | F. aeria |
| 4. Aerial mycelium absent to spare on PDA | 5 |
| 5. Colony on PDA light yellow to deep orange | F. matuoi |
| 5. Colony on PDA pinkish orange | F. gigas |
| 6. Macroconidia filiform | F. filiformis |
| 6. Macroconidia falcate | 7 |
| 7. Producing orange-yellow pigment on PDA | F. guangxiensis |
| 7. Producing pink pigment on PDA | F. aquaeductuum |
4. Discussion
Since the establishment of Fusarium Link in 1809, many fusarioid species have been assigned to the genus and the generic boundary has become obscure. The accumulated morphological and phylogenetic data suggested that the genus was heterogeneous [30]. Efforts were made toward the construction of a monophyletic Fusarium as well as its allies [31,32]. The previously recognized members classified in Fusarium sensu lato are now treated as separate genera, i.e., Albonectria Rossman & Samuels, Atractium Link, Bisifusarium L. Lombard, Crous & W. Gams, Cosmosporella S.K. Huang, R. Jeewon & K.D. Hyde, Cyanonectria Samuels & P. Chaverri, Dialonectria (Sacc.) Cooke, Fusicolla, Geejayessia Schroers, Gräfenhan & Seifert, Macroconia (Wollenw.) Gräfenhan, Seifert & Schroers, Microcera Desm., Neocosmospora E.F. Sm., Pseudofusicolla D. Triest, Rectifusarium (L. Lombard, Crous & W. Gams) and Stylonectria Höhn. [2,33,34].
Several studies have shown that members of Fusicolla are economically important in the fields of human health [11,35,36,37,38], fermentation [39,40], ecology [41,42] and agriculture [13,43,44,45]. For example, Fusicolla species were related to gastric cancer and disorganized lipid metabolism in patients with nonalcoholic fatty liver disease [37,38]. Fusicolla merismoides (Corda) Gräfenhan, Seifert & Schroers (as Fusarium merismoides Corda) was reported as an important source of anticancer agents [35], and F. violacea can produce secondary bioactive metabolites that may be potential biological agents [13,43]. Thus, studies on the biodiversity of Fusicolla are of theoretical and practical importance and should be continuously and extensively carried out.
The phylogenetic overview of Fusicolla based on multilocus sequence analyses showed that the genus is monophyletic [2]. The present phylogeny, including the newly added taxa, inferred from sequences of the acl1, ITS, LSU, rpb2 and tub2 regions, resulted in a similar tree topology to that demonstrated in the previous studies [8,9,46,47]. The result indicated that the four Chinese strains (CGMCC 3.24907, 3.24908, 3.24909 and 3.24910) grouped with the known species of Fusicolla (BIBP/MLBP = 1.0/96%), which confirmed their taxonomic placements. Fusicolla filiformis is associated with, but clearly separated from, F. gigas (BIBP/MLBP = 1.0/100%) and is characterized by filiform macroconidia. Fusicolla aeria is grouped with F. acetilerea and F. elongata, all three species forming abundant aerial mycelia on PDA. Fusicolla coralloidea, representing an independent linage, can be easily distinguished by its coralloid synnemata in culture and rod-shaped to ellipsoidal microconidia.
Among the known species of Fusicolla, F. aquaeductuum, F. betae, F. bharatavarshae Devadatha, V.V. Sarma & E.B.G. Jones, F. epistroma, F. melogrammae Lechat & Aplin, F. ossicola Lechat & Rossman and F. siamensis R.H. Perera, E.B.G. Jones & K.D. Hyde were described with both sexual and asexual stages [2,4,5,12,47,48]. However, F. cassiae-fistulae R.H. Perera, E.B.G. Jones & K.D. Hyde, F. gigantispora Dayar. & K.D. Hyde and F. reyesiana (Sacc.) Forin & Vizzini are only known from their sexual stages, and the remaining taxa are reported solely with their asexual stages [2,5,8,10,34,46,47,49], as well as the newly described species. Large-scale surveys covering different ecosystems and substrates in unexplored regions will further improve our knowledge of the species diversity of the genus and establish connections between the sexual and asexual stages of Fusicolla species, which will permit a better understanding of the whole fungus.
Author Contributions
Conceptualization, Z.-Q.Z. and W.-Y.Z.; methodology, software, validation, formal analysis, investigation, Z.-Q.Z.; resources, W.-Y.Z. and Z.-Q.Z.; data curation, Z.-Q.Z.; writing—original draft preparation, Z.-Q.Z.; writing—review and editing, W.-Y.Z. and Z.-Q.Z.; visualization, Z.-Q.Z.; supervision, Z.-Q.Z. and W.-Y.Z.; project administration, W.-Y.Z.; funding acquisition, W.-Y.Z. and Z.-Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (32270009, 31750001, 31870012); the Biological Resources Programme, Chinese Academy of Sciences (KFJ-BRP-017-082); and the Frontier Key Program of the Chinese Academy of Sciences (QYZDY-SSW-SMC029).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The names of the new species were formally registered in the database Fungal Names (https://nmdc.cn/fungalnames (accessed on 20 February 2023)). Specimens were deposited in the Herbarium Mycologicum Academiae Sinicae (https://nmdc.cn/fungarium/ (accessed on 18 February 2023)). Cultures were deposited in the China General Microbiological Culture Collection Center (https://cgmcc.net/ (accessed on 4 April 2023)). The newly generated sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank (accessed on 29 December 2022)).
Acknowledgments
The authors would like to thank Z.H. Yu, H.D. Zheng, J.X. Deng and Z.X. Zhu for collecting samples jointly for this study and to T. Huang for the technical help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bonorden, H.F. Handbuch der Allgemeinen Mykologie; Schweizerbart: Stuttgart, Germany, 1851; pp. 1–336. [Google Scholar]
- Gräfenhan, T.; Schroers, H.J.; Nirenberg, H.I.; Seifert, K.A. An overview of the taxonomy, phylogeny, and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella, and Volutella. Stud. Mycol. 2011, 68, 79–113. [Google Scholar] [CrossRef] [PubMed]
- Lombard, L.; Van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed]
- Lechat, C.; Rossman, A.R. A new species of Fusicolla (Hypocreales), F. ossicola, from Belgium. Ascomycete.org 2017, 9, 225–228. [Google Scholar]
- Jones, E.B.G.; Devadatha, B.; Abdel-Wahab, M.A.; Dayarathne, M.C.; Zhang, S.N.; Hyde, K.D.; Liu, J.K.; Bahkali, A.H.; Sarma, V.V.; Tibell, S.; et al. Phylogeny of new marine Dothideomycetes and Sordariomycetes from mangroves and deep-sea sediments. Bot. Mar. 2019, 63, 155–181. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morphomolecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Jaeduk, G.; Hye, Y.M. Diversity of fungi in brackish water in Korea. Kor. J. Mycol. 2020, 48, 457–473. [Google Scholar]
- Singh, S.K.; Rana, S.; Bhat, J.D.; Singh, P.N. Morphology and phylogeny of a novel species of Fusicolla (Hypocreales, Nectriaceae), isolated from the air in the Western Ghats, India. J. Fungal Res. 2020, 4, 258–265. [Google Scholar]
- Liu, C.; Zhuang, W.Y.; Yu, Z.H.; Zeng, Z.Q. Two new species of Fusicolla (Hypocreales) from China. Phytotaxa 2022, 536, 165–174. [Google Scholar] [CrossRef]
- Crous, P.W.; Sandoval-Denis, M.; Costa, M.M.; Groenewald, J.Z.; van Iperen, A.L.; Starink-Willemse, M.; Hernández-Restrepo, M.; Kandemir, H.; Ulaszewski, B.; de Boer, W.; et al. Fusarium and allied fusarioid taxa (FUSA). 1. Fungal Syst. Evol. 2022, 9, 161–200. [Google Scholar] [CrossRef]
- Bai, X.; Zhang, T.; Qu, Z.; Li, H.; Yang, Z. Contribution of filamentous fungi to the musty odorant 2,4,6-trichloroanisole in water supply reservoirs and associated drinking water treatment plants. Chemosphere 2017, 182, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.K.; Jeewon, R.; Hyde, K.D.; Bhat, D.; Wen, T.C. Novel taxa within Nectriaceae: Cosmosporella gen. nov. and Aquanectria sp. nov. from freshwater habitats in China. Cryptogam. Mycol. 2018, 39, 169–192. [Google Scholar] [CrossRef]
- Li, W.; Long, Y.; Mo, F.; Shu, R.; Yin, X.; Wu, X.; Zhang, R.; Zhang, Z.; He, L.; Chen, T.; et al. Antifungal activity and biocontrol mechanism of Fusicolla violacea J-1 against soft rot in Kiwifruit caused by Alternaria alternata. J. Fungi 2021, 7, 937. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H.I. Studies on the morphologic and biologic differentiation in Fusarium section Liseola. Mitt. Biol. Bundesanst. Land-Forstw. 1976, 169, 1–117. [Google Scholar]
- Wang, L.; Zhuang, W.Y. Designing primer sets for amplification of partial calmodulin genes from penicillia. Mycosystema 2004, 23, 466–473. [Google Scholar]
- Nowrousian, M.; Kück, U.; Loser, K.; Weltring, K.M. The fungal acl1 and acl2 genes encode two polypeptides with homology to the N- and C-terminal parts of the animal ATP citrate lyase polypeptide. Curr. Genet. 2000, 37, 189–193. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analyzed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA Polymerase II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgin, D.G. The ClustalX windows interface: Flexible strategies for multiple sequences alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest v2, Program distributed by the author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004.
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Mol. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed]
- Hosoya, T.; Tubaki, K. Fusarium matuoi sp. nov. and its teleomorph Cosmospora matuoi sp. nov. Mycoscience 2004, 45, 261–270. [Google Scholar] [CrossRef]
- O’Donnell, K. Fusarium and its near relatives. In The Fungal Holomorph: Mitotic, Meiotic and Pleomorphic Speciation in Fungal Systematic; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 224–233. [Google Scholar]
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–248. [Google Scholar]
- Summerbell, R.C.; Schroers, H.J. Analysis of phylogenetic relationship of Cylindrocarpon lichenicola and Acremonium falciforme to the Fusarium solani species complex and a review of similarities in the spectrum of opportunistic infections caused by these fungi. J. Clin. Microbiol. 2002, 40, 2866–2875. [Google Scholar] [CrossRef] [PubMed]
- Schroers, H.J.; Gräfenhan, T.; Nirenberg, H.I.; Seifert, K.A. A revision of Cyanonectria and Geejayessia gen. nov., and related species with Fusarium-like anamorphs. Stud. Mycol. 2011, 68, 115–138. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, S.; Yanagisawa, M.; Katoh, A.; Fujii, T.; Sano, T.; Matsukuma, S.; Furumai, T.; Fujiu, M.; Watanabe, K.; Yokose, K.; et al. Fusarium merismoides CORDA NR 6356, the source of the protein kinase C inhibitor, azepinostatin. taxonomy, yield improvement, fermentation and biological activity. J. Antibiot. 1994, 47, 639–647. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, R.; Koss, M.; Ziegler, D.; De Respinis, S.; Petrini, O. Fungi in water samples of a full-scale water work. Mycol. Prog. 2018, 17, 467–478. [Google Scholar] [CrossRef]
- You, N.; Xu, J.; Wang, L.; Zhuo, L.; Zhou, J.; Song, Y.; Ali, A.; Luo, Y.; Yang, J.; Yang, W.; et al. Fecal fungi dysbiosis in nonalcoholic fatty liver disease. Obesity 2021, 29, 350–358. [Google Scholar] [CrossRef]
- Zhong, M.Y.; Xiong, Y.B.; Zhao, J.B.; Gao, Z.; Ma, J.S.; Wu, Z.X.; Song, Y.X.; Hong, X.H. Candida albicans disorder is associated with gastric carcinogenesis. Theranostics 2021, 11, 4945–4956. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Yu, D.; Gao, P.; Jiang, Q.; Yang, F. Dynamics and diversity of microbial community succession during fermentation of Suan yu, a Chinese traditional fermented fish, determined by high throughput sequencing. Food Res. Int. 2018, 111, 565–573. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Huang, Y.G. Structure and diversity analysis of mold community in main Maotai-flavor baijiu brewing areas of Maotai town using high-throughput sequencing. Food Sci. 2021, 42, 150–156. [Google Scholar]
- Clocchiatti, A.; Hannula, S.E.; van den Berg, M.; Hundscheid, M.P.J.; de Boer, W. Evaluation of phenolic root exudates as stimulants of saptrophic fungi in the rhizosphere. Front. Microbiol. 2021, 12, 644046. [Google Scholar] [CrossRef]
- Zhu, Q.; Wang, N.; Duan, B.; Wang, Q.; Wang, Y. Rhizosphere bacterial and fungal communities succession patterns related to growth of poplar fine roots. Sci. Total Environ. 2021, 756, 143839. [Google Scholar] [CrossRef]
- Hoch, H.C.; Abawi, G.S. Mycoparasitism of oospores of Pythium ultimum by Fusarium merismoides. Mycologia 1979, 71, 621–625. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, Y.; Zhou, T.; Akkaya, M.S.; Wang, L.; Li, S.; Li, X. Biocontrol of Fusarium wilt disease in strawberries using bioorganic fertilizer fortified with Bacillus licheniformis X-1 and Bacillus methylotrophicus Z-1. 3 Biotech 2020, 10, 80. [Google Scholar] [CrossRef]
- Wang, M.; Xue, J.; Ma, J.; Feng, X.; Ying, H.; Xu, H. Streptomyces lydicus M01 regulates soil microbial community and alleviates foliar disease caused by Alternaria alternata on cucumbers. Front. Microbiol. 2020, 11, 942. [Google Scholar] [CrossRef] [PubMed]
- Forin, N.; Vizzini, A.; Nigris, S.; Ercole, E.; Voyron, S.; Girlanda, M.; Baldan, B. Illuminating type collections of nectriaceous fungi in Saccardo’s fungarium. Persoonia 2020, 45, 221–249. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; Mckenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef]
Figure 1.
The BI tree generated based on the combined datasets of acl1, ITS, LSU, rpb2 and tub2 sequences of Fusicolla species. BIPP (left) values greater than 0.9 and MLBP (right) values greater than 70% are shown at the nodes. Macroconia leptosphaeriae and Microcera larvarum were chosen as outgroup taxa.
Figure 1.
The BI tree generated based on the combined datasets of acl1, ITS, LSU, rpb2 and tub2 sequences of Fusicolla species. BIPP (left) values greater than 0.9 and MLBP (right) values greater than 70% are shown at the nodes. Macroconia leptosphaeriae and Microcera larvarum were chosen as outgroup taxa.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of Fusicolla species, herbarium/strain numbers and GenBank accession numbers of materials used in this study.
Table 1.
List of Fusicolla species, herbarium/strain numbers and GenBank accession numbers of materials used in this study.
| Species | Herbarium/Strain Numbers | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| acl1 | ITS | LSU | rpb2 | tub2 | ||
| F. acetilerea | BBA 63789 T | HQ897839 | HQ897790 | U88108 | HQ897701 | − |
| F. aeria | CGMCC 3.24908 T | OQ134105 a | OQ128334 a | OQ128338 a | OQ134111 a | OQ134100 a |
| CGMCC 3.24909 | OQ134106 a | OQ128335 a | OQ128339 a | OQ134112 a | OQ134101 a | |
| F. aquaeductuum | CBS 837.85 T | HQ897880 | KM231823 | KM231699 | HQ897744 | − |
| F. betae | BBA 64317 T | HQ897917 | − | − | HQ897781 | − |
| F. bharatavarshae | NFCCI 4423 T | − | MK152510 | MK152511 | MK157022 | MK376462 |
| F. cassiae-fistulae | MFLUCC 19-0318 T | − | NR171299 | NG073862 | − | − |
| F. coralloidea | CGMCC 3.24907 T | OQ134104 a | OQ128333 a | OQ128337 a | OQ134110 a | OQ134099 a |
| F. elongata | CBS 148934 T | ON759286 | ON763203 | ON763200 | ON759297 | ON745628 |
| F. epistroma | BBA 62201 T | HQ897901 | − | AF228352 | HQ897765 | − |
| F. filiformis | CGMCC 3.24910 T | OQ134103 a | OQ128332 a | OQ128336 a | OQ134109 a | OQ134098 a |
| F. gigantispora | MFLU 16-1206 T | − | MN047104 | MN017869 | − | − |
| F. gigas | CGMCC 3.20680 T | OQ134107 a | OK465362 | OK465449 | OQ134113 a | OQ134102 a |
| F. guangxiensis | CGMCC 3.20679 T | OQ134108 a | OK465363 | OK465450 | OQ134114 a | − |
| F. hughesii | NFCCI 4234 T | − | MG779450 | MG779452 | − | − |
| F. matuoi | CBS 581.78 T | HQ897858 | KM231822 | KM231698 | HQ897720 | KM232093 |
| F. melogrammae | CBS 141092 T | − | KX897140 | NG058275 | − | MW834305 |
| F. meniscoidea | CBS 110189 T | MW834043 | MW827613 | MW827654 | MW834010 | MW834306 |
| F. merismoides | CBS 186.34 T | − | MH855482 | MH866963 | − | − |
| F. ossicola | CBS 140161 T | − | NR161034 | MF628021 | MW834011 | MW834307 |
| F. quarantenae | URM 8367 T | − | MW553789 | MW553788 | MW556626 | MW556624 |
| F. septimanifiniscientiae | CBS 144935 T | − | MK069422 | MK069418 | − | MK069408 |
| F. siamensis | MFLUCC 172577 T | − | NR171300 | NG073863 | − | − |
| F. sporellula | CBS 110191 T | MW834044 | MW827614 | MW827655 | MW834012 | MW834308 |
| F. violacea | CBS 634.76 T | KM231059 | KM231824 | KM231700 | HQ897696 | KM232095 |
| Macroconia leptosphaeriae | CBS 100001 | HQ897891 | HQ897810 | KC291787 | HQ728164 | KM232097 |
| Microcera larvarum | CBS 738.79/AR 4580 | KM231060 | KM231825 | KM231701 | KM232387 | KC291935 |
T indicates the ex-type culture. a indicates the newly provided sequences.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zeng, Z.-Q.; Zhuang, W.-Y. Three New Species of Fusicolla (Hypocreales) from China. J. Fungi 2023, 9, 572. https://doi.org/10.3390/jof9050572
AMA Style
Zeng Z-Q, Zhuang W-Y. Three New Species of Fusicolla (Hypocreales) from China. Journal of Fungi. 2023; 9(5):572. https://doi.org/10.3390/jof9050572
Chicago/Turabian StyleZeng, Zhao-Qing, and Wen-Ying Zhuang. 2023. "Three New Species of Fusicolla (Hypocreales) from China" Journal of Fungi 9, no. 5: 572. https://doi.org/10.3390/jof9050572
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.