
Multi-Locus Phylogenetic Analysis Revealed the Association of Six Colletotrichum Species with Anthracnose Disease of Coffee (Coffea arabica L.) in Saudi Arabia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. Molecular Characterization
2.2.1. DNA Extraction, PCR Amplification, and Sequencing
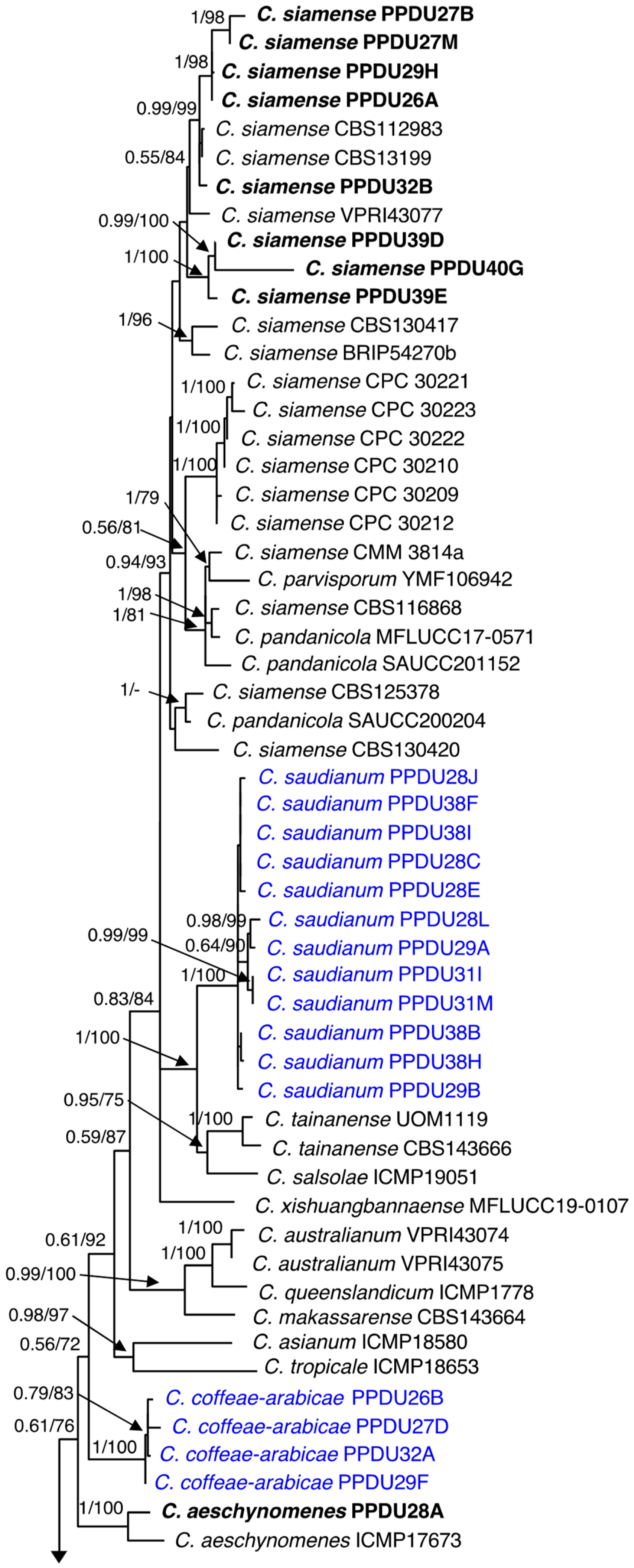
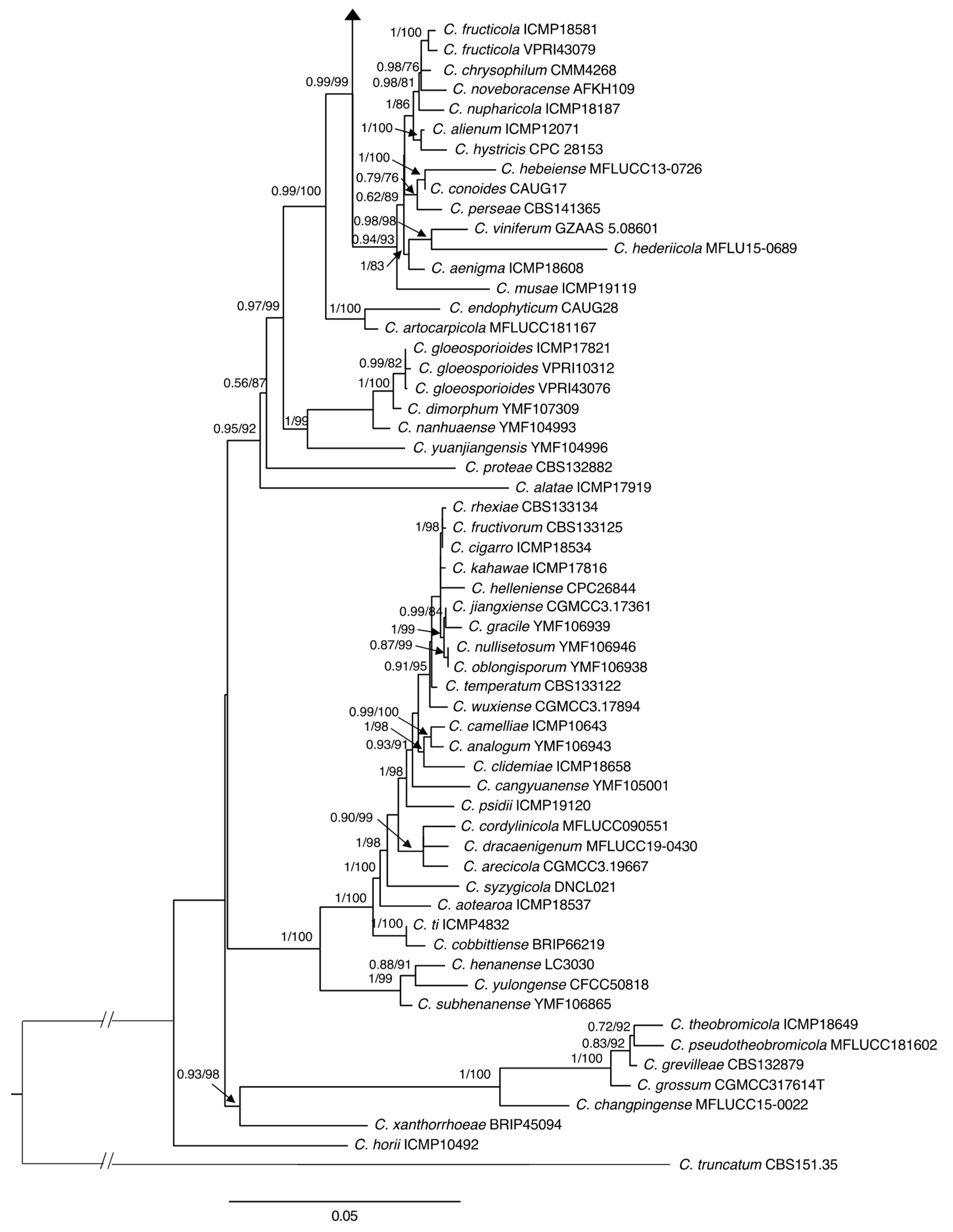
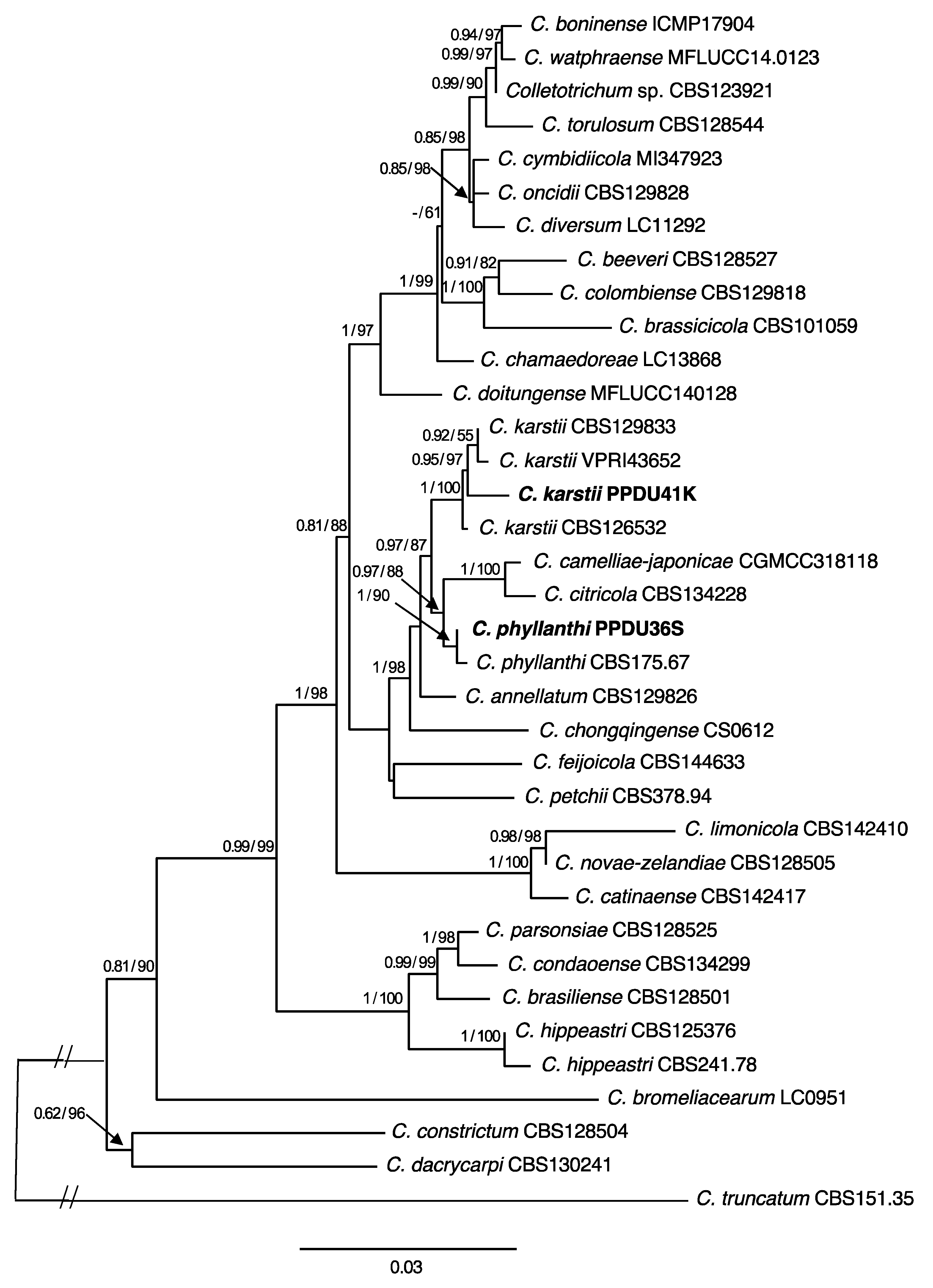
2.2.2. Phylogenetic Analyses
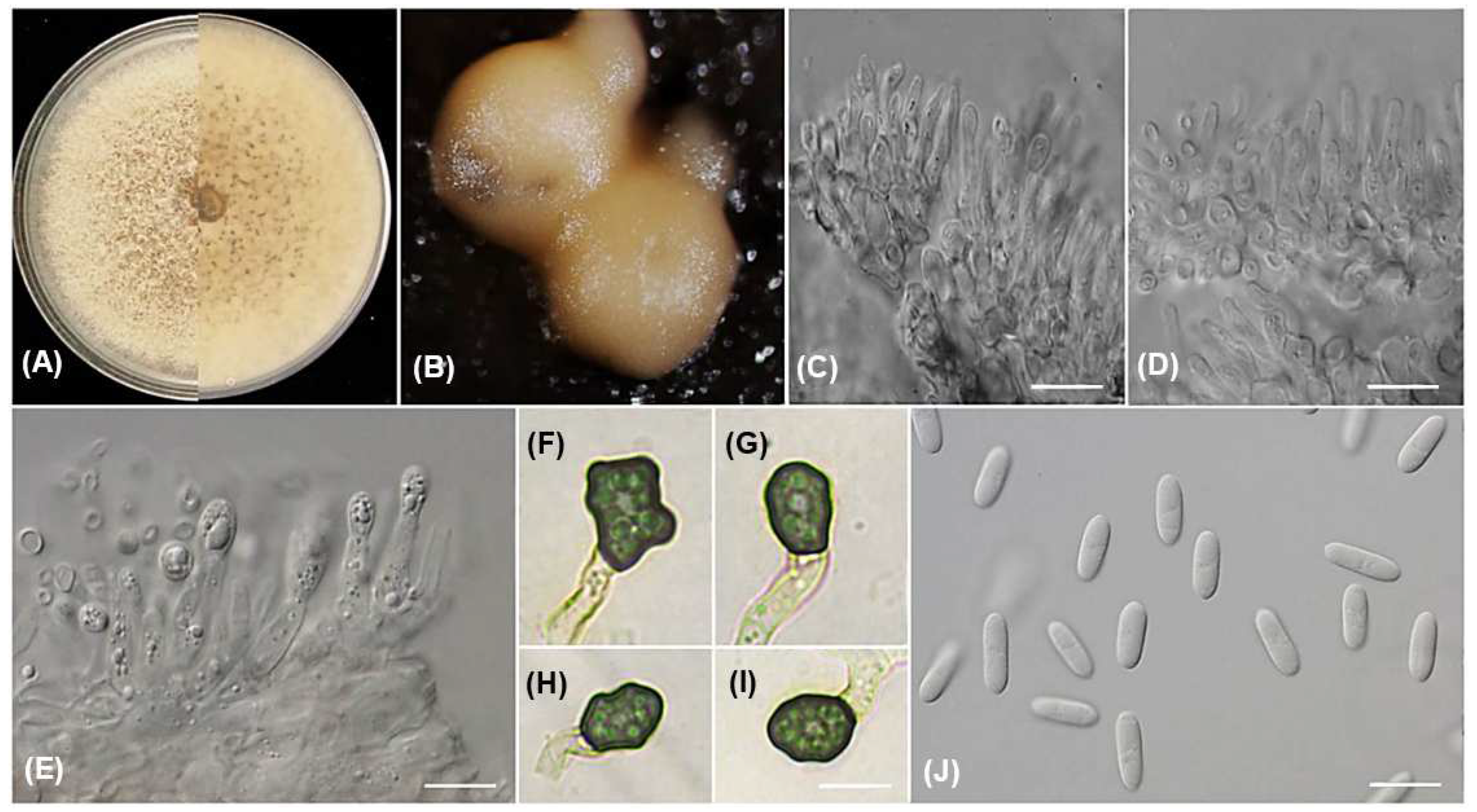
2.3. Morphological Characterization
2.4. Pathogenicity Tests
2.5. Data Analysis
3. Results
3.1. Symptoms Observation and Isolation
3.2. Molecular Characterization
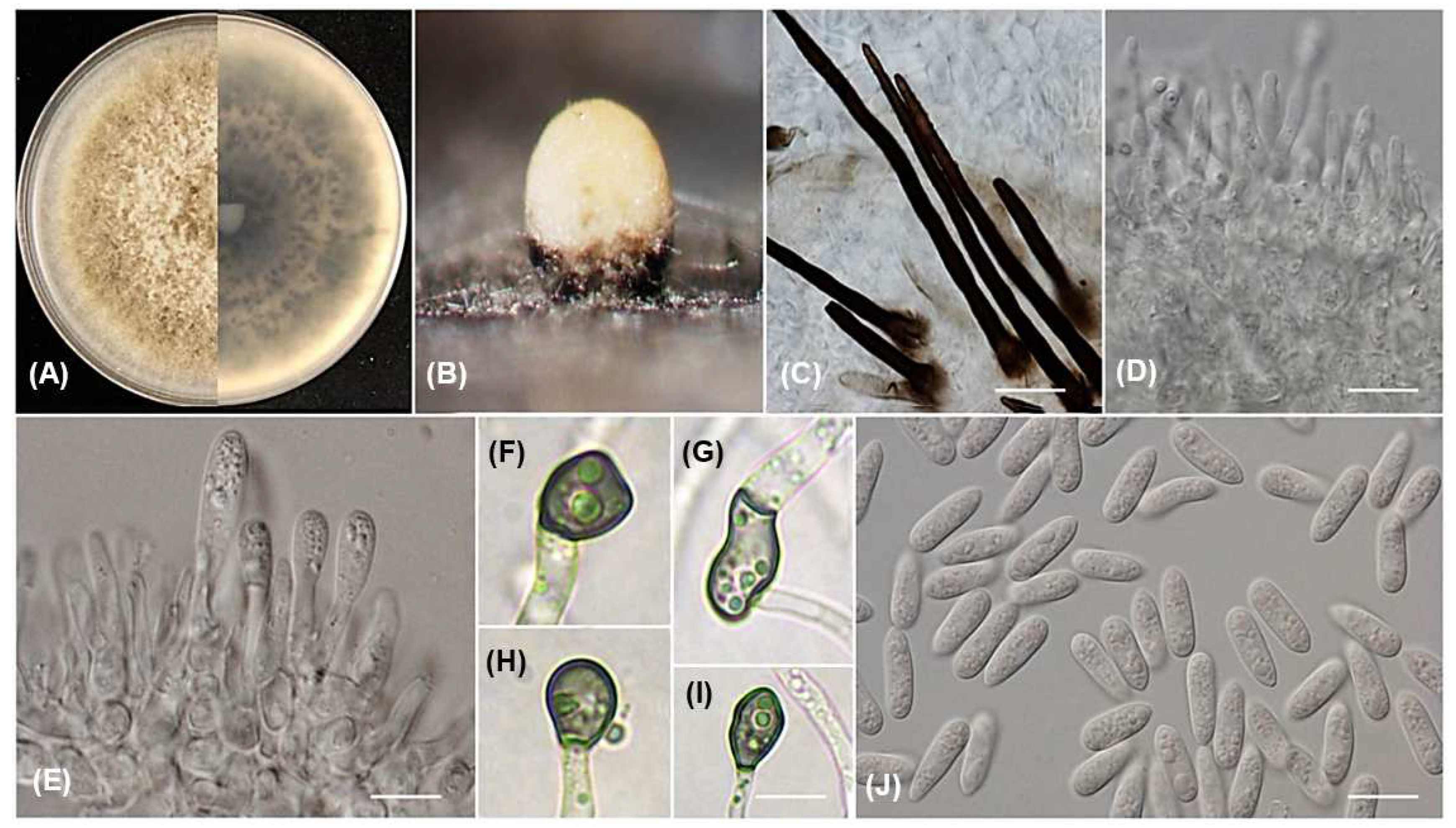
3.3. Taxonomy
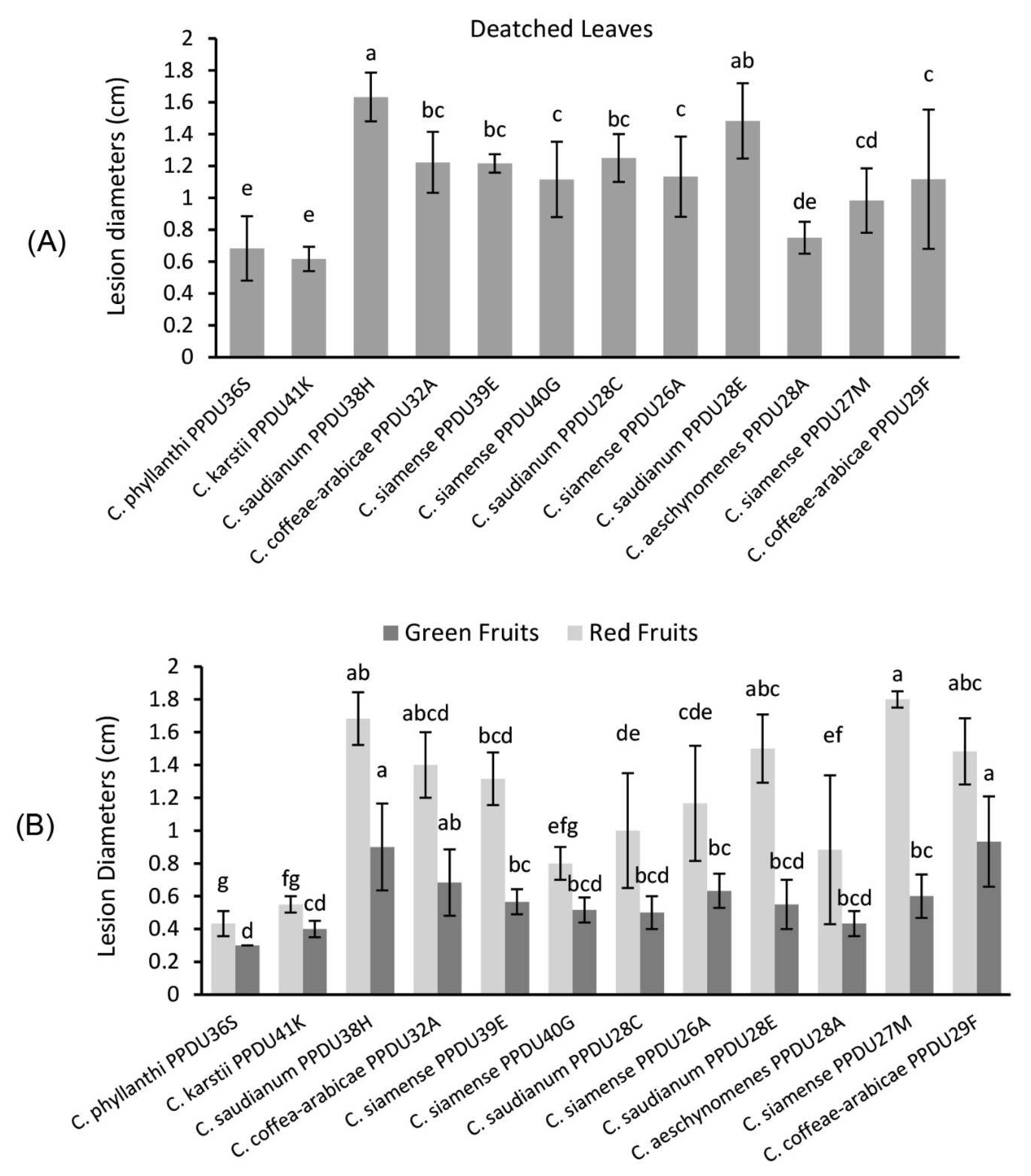
3.4. Pathogenicity Tests
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hindorf, H.; Omondi, C.O. A Review of Three Major Fungal Diseases of Coffea arabica L. in the Rainforests of Ethiopia and Progress in Breeding for Resistance in Kenya. J. Adv. Res. 2011, 2, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmari, K.; Abu Zeid, I.; Al-Attar, A. Coffee Arabica in Saudi Arabia: An Overview. Int. J. Pharm. Phytopharm. Res. 2020, 10, 71–78. [Google Scholar]
- Eskes, A. Identification, Description, and Collection of Coffee Types in P. D. R. Yemen; IPGRI: Rome, Italy, 1989. [Google Scholar]
- Tounekti, T.; Mosbah, M.; Al-Turki, T.; Khemira, H. Genetic Diversity Analysis of Coffee (Coffea arabica L.) Germplasm Accessions Growing in the Southwestern Saudi Arabia Using Quantitative Traits. Nat. Resour. 2017, 8, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Avelino, J.; Allinne, C.; Cerda, R.; Willocquet, L.; Savary, S. Multipledisease System in Coffee: From Crop Loss Assessment to Sustainable Management. Annu. Rev. Phytopathol. 2018, 56, 611–635. [Google Scholar] [CrossRef]
- McDonald, J. A Preliminary Account of a Disease of Green Coffee Berries in Kenya Colony. Trans. Br. Mycol. Soc. 1926, 11, 145–154. [Google Scholar] [CrossRef]
- Waller, J.M.; Bigger, M.; Hillocks, R. Coffee Pests, Diseases and Their Management; CABI: Wallingford, UK, 2007; 434p, ISBN 978-1-84593-209-1. [Google Scholar]
- Weir, B.; Johnston, P.; Damm, U. The Colletotrichum gloeosporioides Species Complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Alemu, F. Assessment of the Current Status of Coffee Diseases at Gedeo and Sidama Zone, Ethiopia. Int. J. Adv. Res. 2013, 1, 192–202. [Google Scholar]
- Rayner, R. Coffee Berry Disease—A Survey of Investigations Carried Out Up to 1950. East Afr. Agric. J. 1952, 17, 130–158. [Google Scholar] [CrossRef]
- Waller, J.M.; Bridge, P.D.; Black, R.; Hakiza, G. Characterization of the Coffee Berry Disease Pathogen, Colletotrichum kahawae sp. Nov. Mycol. Res. 1993, 97, 989–994. [Google Scholar] [CrossRef]
- Hindorf, H. Colletotrichum–Population Auf Coffea arabica L. in Kenya: II. Qualitative and Quantitative Unterschiede in Der Colletotrichum–Population. J. Phytopathol. 1973, 77, 216–234. [Google Scholar] [CrossRef]
- Hindorf, H. Colletotrichum–Populationen Auf Coffea arabica L. in Kenia: III. Verbreitung von Colletotrichum-Arten Auf Den Einzelnen Organen Des Kaffeestrauches. J. Phytopathol. 1973, 77, 324–338. [Google Scholar] [CrossRef]
- Hindorf, H. Colletotrichum-Population Auf Coffea arabica L. in Kenia: I. Eine Methode Zur Systematischen Trennung von Pilzpopulationen. J. Phytopathol. 1973, 77, 97–116. [Google Scholar] [CrossRef]
- Lu, L.; Tibpromma, S.; Karunarathna, S.C.; Jayawardena, R.S.; Lumyong, S.; Xu, J.; Hyde, K.D. Comprehensive Review of Fungi on Coffee. Pathogens 2022, 11, 411. [Google Scholar] [CrossRef] [PubMed]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.; Hyde, K.D. Characterization of Colletotrichum Species Associated with Coffee Berries in Northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Nguyen, P.T.H.; Pettersson, O.V.; Olsson, P.; Liljeroth, E. Identification of Colletotrichum Species Associated with Anthracnose Disease of Coffee in Vietnam. Eur. J. Plant Pathol. 2010, 127, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Cai, L.; Crous, P.W.; Damm, U. The Colletotrichum gigasporum Species Complex. Pers. Mol. Phylogeny Evol. Fungi 2014, 33, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Cristóbal-Martínez, A.L.; de Jesús Yáñez-Morales, M.; Solano-Vidal, R.; Segura-León, O.; Hernández-Anguiano, A.M. Diversity of Colletotrichum Species in Coffee (Coffea arabica) Plantations in Mexico. Eur. J. Plant Pathol. 2017, 147, 605–614. [Google Scholar] [CrossRef]
- Cao, X.R.; Xu, X.M.; Che, H.Y.; West, J.S.; Luo, D.Q. Characteristics and Distribution of Colletotrichum Species in Coffee Plantations in Hainan, China. Plant Pathol. 2019, 68, 1146–1156. [Google Scholar] [CrossRef]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum Species Complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum-Current Status and Future Directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [Green Version]
- de Silva, D.D.; Groenewald, J.Z.; Crous, P.W.; Ades, P.K.; Nasruddin, A.; Mongkolporn, O.; Taylor, P.W.J. Identification, Prevalence and Pathogenicity of Colletotrichum Species Causing Anthracnose of Capsicum annuum in Asia. IMA Fungus 2019, 10, 8. [Google Scholar] [CrossRef]
- Khodadadi, F.; González, J.B.; Martin, P.L.; Giroux, E.; Bilodeau, G.J.; Peter, K.A.; Doyle, V.P.; Aćimović, S.G. Identification and Characterization of Colletotrichum Species Causing Apple Bitter Rot in New York and Description of C. noveboracense sp. nov. Sci. Rep. 2020, 10, 11043. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-W.; Manawasinghe, I.; Liao, X.-N.; Mao, J.; Dong, Z.; Jayawardena, R.; Wanasinghe, D.; Shu, Y.-X.; Luo, M. Endophytic Colletotrichum (Sordariomycetes, Glomerellaceae) Species Associated with Citrus grandis cv. “Tomentosa” in China. MycoKeys 2023, 95, 163–188. [Google Scholar] [CrossRef]
- Guevara-Suarez, M.; Cárdenas, M.; Jiménez, P.; Afanador-Kafuri, L.; Restrepo, S. Colletotrichum Species Complexes Associated with Crops in Northern South America: A Review. Agronomy 2022, 12, 548. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense Species Complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef] [Green Version]
- dos Santos Vieira, W.A.; Bezerra, P.A.; da Silva, A.C.; Veloso, J.S.; Câmara, M.P.S.; Doyle, V.P. Optimal Markers for the Identification of Colletotrichum Species. Mol. Phylogenet. Evol. 2020, 143, 106694. [Google Scholar] [CrossRef] [PubMed]
- de Silva, D.; Talhinhas, P.; Várzea, V.; Cai, L.; Paulo, O.; Batista, D. Application of the Apn2/MAT Locus to Improve the Systematics of the Colletotrichum gloeosporioides Complex: An Example from Coffee (Coffea spp.) Hosts. Mycologia 2012, 104, 396–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manova, V.; Stoyanova, Z.; Rodeva, R.; Boycheva, I.; Korpelainen, H.; Vesterinen, E.; Wirta, H.; Bonchev, G. Morphological, Pathological and Genetic Diversity of the Colletotrichum Species, Pathogenic on Solanaceous Vegetable Crops in Bulgaria. J. Fungi 2022, 8, 1123. [Google Scholar] [CrossRef]
- Al-Faifi, Z.; Alsolami, W.; Abada, E.; Khemira, H.; Almalki, G.; Modafer, Y. Fusarium oxysporum and Colletotrichum musae Associated with Wilt Disease of Coffea arabica in Coffee Gardens in Saudi Arabia. Can. J. Infect. Dis. Med. Microbiol. 2022, 2022, 3050495. [Google Scholar] [CrossRef]
- Agostini, J.P.; Timmer, L.W. Selective Isolation Procedures for Differentiation of Two Strains of Colletotrichum gloeosporioides from Citrus. Plant Dis. 1992, 76, 1176–1178. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N. Morphological Approaches in Studying Fungi: Collection, Examination, Isolation, Sporulation and Preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A Plant DNA Minipreparation: Version II. Plant Mol. Biol. Report. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. A Guid. Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic RDNA ITS2 Types within a Monophyletic Lineage of the Fungus Fusarium are Non-orthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and Molecular Characterization of the Glyceraldehyde-3-Phosphate Dehydrogenase-Encoding Gene and CDNA from the Plant Pathogenic Fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Qiao, Y.-H.; Zhang, C.-N.; Li, M.; Li, H.; Mao, Y.-F.; Chen, F.-M. Species of the Colletotrichum spp., the Causal Agents of Leaf Spot on European Hornbeam (Carpinus betulus). J. Fungi 2023, 9, 489. [Google Scholar] [CrossRef]
- Liu, F.; Weir, B.S.; Damm, U.; Crous, P.W.; Wang, Y.; Liu, B.; Wang, M.; Zhang, M.; Cai, L. Unravelling Colletotrichum Species Associated with Camellia: Employing ApMat and GS Loci to Resolve Species in the C. gloeosporioides Complex. Pers. Mol. Phylogeny Evol. Fungi 2015, 35, 63–86. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Ma, Z.Y.; Hou, L.W.; Diao, Y.Z.; Wu, W.P.; Damm, U.; Song, S.; Cai, L. Updating Species Diversity of Colletotrichum, with a Phylogenomic Overview. Stud. Mycol. 2022, 101, 1–56. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An Online Initiative to Launch Mycology into the 21st Century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Batista, D.; Silva, D.N.; Vieira, A.; Cabral, A.; Pires, A.S.; Loureiro, A.; Guerra-Guimaraes, L.; Pereira, A.P.; Azinheira, H.; Talhinhas, P. Legitimacy and Implications of Reducing Colletotrichum kahawae to Subspecies in Plant Pathology. Front. Plant Sci. 2017, 7, 2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.-J.; Grabke, A.; Schnabel, G. Investigation of the Colletotrichum gloeosporioides Species Complex Causing Peach Anthracnose in South Carolina. Plant Dis. 2015, 99, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Crouch, J.A.; Clarke, B.; Hillman, B. What Is the Value of ITS Sequence Data in Colletotrichum Systematics and Species Diagnosis? A Case Study Using the Falcate-Spored Graminicolous Colletotrichum Group. Mycologia 2009, 101, 648–656. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar, N.; Weir, B.S.; Hyde, K.D.; Shenoy, B.D. The ApMat Marker Can Resolve Colletotrichum Species: A Case Study with Mangifera indica. Fungal Divers. 2013, 61, 117–138. [Google Scholar] [CrossRef]
- Sharma, G.; Pinnaka, A.K.; Shenoy, B.D. Resolving the Colletotrichum siamense Species Complex Using ApMat Marker. Fungal Divers. 2014, 71, 247–264. [Google Scholar] [CrossRef]
- Vieira, W.A.S.; Michereff, S.J.; de Morais, M.A.; Hyde, K.D.; Câmara, M.P.S. Endophytic Species of Colletotrichum Associated with Mango in Northeastern Brazil. Fungal Divers. 2014, 67, 181–202. [Google Scholar] [CrossRef]
- Wu, J.; Hu, S.; Ye, B.; Hu, X.; Xiao, W.; Yu, H.; Chuanqing, Z. Diversity and Resistance to Thiophanate-Methyl of Colletotrichum spp. in Strawberry Nursery and the Development of Rapid Detection Using LAMP Method. Agronomy 2022, 12, 2815. [Google Scholar] [CrossRef]
- Ismail, A.M.; Cirvilleri, G.; Yaseen, T.; Epifani, F.; Perrone, G.; Polizzi, G. Characterization of Colletotrichum Species Causing Anthracnose Disease of Mango in Italy. J. Plant Pathol. 2015, 97, 167–171. [Google Scholar] [CrossRef]
- Nguyen, T.H.P.; Säll, T.; Bryngelsson, T.; Liljeroth, E. Variation among Colletotrichum gloeosporioides Isolates from Infected Coffee Berries at Different Locations in Vietnam. Plant Pathol. 2009, 58, 898–909. [Google Scholar] [CrossRef]
- Kenny, M.; Galea, V.; Price, T. Germination and Growth of Colletotrichum acutatum and Colletotrichum gloeosporioides Isolates from Coffee in Papua New Guinea and Their Pathogenicity to Coffee Berries. Australas. Plant Pathol. 2012, 41, 519–528. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| District | No. of Farms | Longitude (E) | Latitude (N) | Altitude (m) |
|---|---|---|---|---|
| Jazan | 1 | 43°8′19.9″ | 17°22′14.3″ | 785 |
| 2 | 43°8′20.4″ | 17°22′22.8″ | 803 | |
| 3 | 43°8′20.4″ | 17°22′27.9″ | 812 | |
| 4 | 43°8′19.9″ | 17°22′14.3″ | 1043 | |
| 5 | 43°8′34.9″ | 17°17′13.5″ | 861 | |
| Asir | 6 | 42°24′39″ | 18°9′41″ | 1880 |
| 7 | 42°22′10″ | 18°11′43″ | 1360 | |
| 8 | 42°23′3″ | 18°11′32″ | 1500 | |
| 9 | 42°38′3″ | 18°13′17″ | 2120 | |
| 10 | 40°18′45″ | 18°11′21″ | 1396 | |
| 11 | 42°19′8″ | 18°12′45″ | 1510 | |
| 12 | 42°6′4″ | 18°49′38″ | 1660 | |
| 13 | 42°5′57″ | 18°49′32″ | 1580 | |
| 14 | 42°4′17″ | 19°9′30″ | 1320 | |
| 15 | 43°10′47″ | 17°40′46″ | 1200 | |
| 16 | 43°10′50″ | 17°40′50″ | 1210 | |
| Najran | 17 | 44°10′20″ | 17°29′5″ | 1290 |
| 18 | 44°3′36″ | 17°26′30″ | 1340 | |
| Al Baha | 19 | 41°25′55.1″ | 19°47′27.5″ | 1100 |
| 20 | 41°21′35″ | 19°45′1.3″ | 1084 | |
| 21 | 41°22′36.1″ | 19°43′35.3″ | 1258 | |
| 22 | 41°21′16.5″ | 19°45′36″ | 1204 | |
| 23 | 41°26′25.7″ | 20°2′9.8″ | 2187 |
| Locus | Product Name | Primer | Sequence (5′–3′) | Reference |
|---|---|---|---|---|
| ITS | Internal transcribed spacer | ITS-1F | CTT GGT CAT TTA GAG GAA GTA A | [35] |
| ITS-4R | TCC TCC GCT TAT TGA TAT GC | |||
| ACT | Actin | ACT-512F | ATG TGC AAG GCC GGT TTC GC | [36] |
| ACT-783R | TAC GAG TCC TTC TGG CCC AT | |||
| CHS-1 | Chitin synthase | CHS-79F | TGG GGC AAG GAT GCT TGG AAG AAG | [36] |
| CHS-345R | TGG AAG AAC CAT CTG TGA GAG TTG | |||
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | GDF | GCC GTC AAC GAC CCC TTC ATT GA | [39] |
| GDR | GGG TGG AGT CGT ACT TGA GCA TGT | |||
| TUB2 | β-Tubulin 2 | T1F | AAC ATG CGT GAG ATT GTA AGT | [37] |
| Bt2bR | ACC CTC AGT GTA GTG ACC CTT GGC | [38] | ||
| ApMat | Mat1–2 | AMF1 | TCATTCTACGTATGTGCCCG | [29] |
| AMR1 | CCAGAAATACACCGAACTTGC |
| Species Identity | Culture No. | Host | Country | GenBank Accession Numbers | |||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | ACT | TUB2 | CHS-1 | GAPDH | ApMat | ||||
| C. aenigma | ICMP 18608 * | Persea americana | Israel | JX010244 | JX009443 | JX010389 | JX009774 | JX010044 | KM360143 |
| C. aeschynomenes | ICMP 17673; ATCC 201874 * | Aeschynomene virginica | USA | JX010176 | JX009483 | JX010392 | JX009799 | JX009930 | KM360145 |
| C. aeschynomenes | PPDU28A | Coffea arabica | Saudi Arabia | OR048775 | OR050686 | OR050783 | OR050738 | OR050756 | OR050711 |
| C. alatae | ICMP 17919 * | Dioscorea alata | India | JX010190 | JX009471 | JX010383 | JX009837 | JX009990 | KC888932 |
| C. alienum | ICMP 12071 * | Malus domestica | New Zealand | JX010251 | JX009572 | JX010411 | JX009882 | JX010028 | KM360144 |
| C. analogum | YMF 1.06943 | Unknown | China | OK030860 | OK513599 | OK513629 | OK513559 | OK513663 | - |
| C. annellatum | CBS 129826 * | Hevea brasiliensis | Colombia | JQ005222 | JQ005570 | JQ005656 | JQ005396 | - | - |
| C. aotearoa | ICMP 18537 * | Coprosma sp. | New Zealand | JX010205 | JX009564 | JX010420 | JX009853 | JX010005 | KC888930 |
| C. arecicola | CGMCC 3.19667 | Areca catechu | China | MK914635 | MK935374 | MK935498 | MK935541 | MK935455 | MK935413 |
| C. artocarpicola | MFLUCC 18–1167 * | Artocarpus heterophyllus | Thailand | MN415991 | MN435570 | MN435567 | MN435569 | MN435568 | - |
| C. asianum | ICMP 18580; CBS 130418 * | Coffea arabica | Thailand | FJ972612 | JX009584 | JX010406 | JX009867 | JX010053 | FR718814 |
| C. australianum | VPRI 43074; UMC001 | Citrus reticulata | Australia | MG572137 | MK473452 | MG572148 | MW091986 | MG572126 | MG572170 |
| C. australianum | VPRI 43075; UMC002 * | Citrus sinensis | Australia | MG572138 | MN442109 | MG572149 | MW091987 | MG572127 | MG572171 |
| C. beeveri | CBS 128527 * | Brachyglottis repanda | New Zealand | JQ005171 | JQ005519 | JQ005605 | JQ005345 | - | - |
| C. boninense | ICMP 17904; CBS 123755 * | Crinum asiaticum var. sinicum | Japan | JQ005153 | JQ005501 | JQ005588 | JQ005327 | - | - |
| C. brasiliense | CBS 128501 * | Passiflora edulis | Brazil | JQ005235 | JQ005583 | JQ005669 | JQ005409 | - | - |
| C. brassicicola | CBS 101059 | Brassica oleracea var. gemmifera | New Zealand | JQ005172 | JQ005520 | JQ005606 | JQ005346 | - | - |
| C. bromeliacearum | LC0951 | Bromeliad | China | MZ595832 | MZ664130 | MZ673956 | MZ799267 | - | - |
| C. camelliae | ICMP 10643 * | Camellia williamsii | United Kingdom | JX010224 | JX009540 | JX010436 | JX009891 | JX009908 | KJ954625 |
| C. camelliae-japonicae | CGMCC 3.18118 *, LC6416 | Camellia japonica | China | KX853165 | KX893576 | KX893580 | MZ799271 | - | - |
| C. cangyuanense | YMF1.05001 | Unknown | China | OK030864 | OK513603 | OK513633 | OK513563 | OK513667 | |
| C. catinaense | CBS 142417; CPC 27978 * | Citrus reticulata | Italy | KY856400 | KY855971 | KY856482 | KY856136 | - | - |
| C. chamaedoreae | LC13868, NN052885 | Chamaedorea erumpens | China | MZ595890 | MZ664188 | MZ674008 | MZ799274 | - | - |
| C. changpingense | MFLUCC 15-0022 | Fragaria ananassa | China | KP683152 | KP683093 | KP852490 | KP852449 | KP852469 | - |
| C. chongqingense | CS0612 | Camellia sinensis | China | MG602060 | MT976107 | MG602044 | MT976117 | - | - |
| C. chrysophilum | CMM4268 *, CMM 4352 | Musa sp. | Brazil | KX094252 | KX093982 | KX094285 | KX094083 | KX094183 | KX094326 |
| C. cigarro | ICMP 18534 | Kunzea ericoides | New Zealand | JX010227 | JX009473 | JX010427 | JX009765 | JX009904 | HE655657 |
| C. citricola | CBS 134228 * | Citrus unchiu | China | KC293576 | KC293616 | KC293656 | KY856140 | - | - |
| C. clidemiae | ICMP 18658 * | Clidemia hirta | USA | JX010265 | JX009537 | JX010438 | JX009877 | JX009989 | KC888929 |
| C. cobbittiense | BRIP 66219 | Cordyline fruticosa | Australia | MH087016 | MH094134 | MH094137 | MH094135 | MH094133 | - |
| C. coffeae-arabicae | PPDU26B | Coffea arabica | Saudi Arabia | OR048779 | OR050690 | OR050787 | OR050742 | OR050760 | OR050715 |
| C. coffeae-arabicae | PPDU27D | Coffea arabica | Saudi Arabia | OR048777 | OR050688 | OR050785 | OR050740 | OR050758 | OR050713 |
| C. coffeae-arabicae | PPDU29F | Coffea arabica | Saudi Arabia | OR048768 | OR050679 | OR050776 | OR050731 | OR050749 | OR050704 |
| C. coffeae-arabicae | PPDU32A | Coffea arabica | Saudi Arabia | OR048764 | OR050675 | OR050772 | OR050727 | OR050745 | OR050700 |
| C. colombiense | CBS 129818 * | unknown | Colombia | JQ005174 | JQ005522 | JQ005608 | JQ005348 | - | - |
| C. condaoense | CBS 134299 | Ipomoea pescaprae | Vietnam | MH229914 | - | MH229923 | MH229926 | - | - |
| C. conoides | CAUG17; MYL24 | Actinidia deliciosa | China | KY995389 | KY995510 | KY995473 | KY995436 | KY995340 | MG198007 |
| C. constrictum | CBS 128504 | Citrus limon | New Zealand | JQ005238 | JQ005586 | JQ005672 | JQ005412 | - | - |
| C. cordylinicola | MFLUCC 090551; ICMP 18579 * | Cordyline fruticosa | Thailand | JX010226 | HM470235 | JX010440 | JX009864 | JX009975 | JQ899274 |
| C. cymbidiicola | IMI 347923 * | Cymbidium sp. | Australia | JQ005166 | JQ005514 | JQ005600 | JQ005340 | - | - |
| C. dacrycarpi | CBS 130241 * | Unknown | New Zealand | JQ005236 | JQ005584 | JQ005670 | JQ005410 | - | - |
| C. dimorphum | YMF1.07309 | Unknown | China | OK030867 | OK513606 | OK513636 | OK513566 | OK513670 | - |
| C. diversum | LC11292, CQ775 | Philodendron selloum | China | MZ595844 | MZ664142 | MZ673965 | MZ799272 | - | - |
| C. doitungense | MFLUCC 14-0128 | Dendrobium sp. | Thailand | MF448524 | MH376385 | MH351277 | - | - | - |
| C. dracaenigenum | MFLUCC 19-0430 | Dracaena sp. | Thailand | MN921250 | MT313686 | - | MT215575 | MT215577 | - |
| C. endophyticum | CAUG28; YTJB1 | Capsicum sp. | China | KP145441 | KP145329 | KP145469 | KP145385 | KP145413 | MH305548 |
| C. feijoicola | CBS 144633, CPC 34245 | Acca sellowiana | Portugal | MK876413 | MK876466 | MK876507 | MK876471 | - | - |
| C. fructicola | ICMP 18581; CBS 130416 * | Coffea arabica | Thailand | JX010165 | FJ907426 | JX010405 | JX009866 | JX010033 | JQ807838 |
| C. fructicola | VPRI 43079; UMC006 | Citrus reticulata | Australia | MG572142 | MK473454 | MG572153 | MW091991 | MG572131 | MG572175 |
| C. fructivorum | CBS 133125 * | Vaccinium macrocarpon | USA | JX145145 | MZ664126 | JX145196 | MZ799259 | MZ664047 | JX145300 |
| C. gloeosporioides | IMI 356878; ICMP 17821; CBS 112999 * | Citrus sinensis | Italy | JX010152 | JX009531 | JX010445 | JX009818 | JX010056 | JQ807843 |
| C. gloeosporioides | VPRI 43076; UMC003 | Citrus sinensis | Australia | MG572139 | MN442110 | MG572150 | MW091988 | MG572128 | MG572172 |
| C. gloeosporioides | VPRI 10312; A01-10312 | Citrus sinensis | Australia | MK469996 | MK470086 | MK470050 | MW091972 | MK470014 | MK470068 |
| C. gracile | YMF1.06939 | Unknown | China | OK030868 | OK513607 | OK513637 | OK513567 | OK513671 | - |
| C. grevilleae | CBS 132879 * | Grevillea sp. | Italy | KC297078 | KC296941 | KC297102 | KC296987 | KC297010 | - |
| C. grossum | CGMCC3.17614T; CAUG7; INIFAT 4145 | Capsicum sp. | China | KP890165 | KP890141 | KP890171 | KP890153 | KP890159 | MG826119 |
| C. hebeiense | MFLUCC13-0726 * | Vitis vinifera | China | KF156863 | KF377532 | KF288975 | KF289008 | KF377495 | KF377562 |
| C. hederiicola | MFLU 15-0689 | Hedera helix | Italy | MN631384 | MN635795 | MN635794 | ON971378 | - | |
| C. helleniense | CPC 26844; CBS 142418; CBS 142419 | Poncirus trifoliata | Greece | KY856446 | KY856019 | KY856528 | KY856186 | KY856270 | MW368907 |
| C. henanense | LC3030; CGMCC 3.17354; LF238 * | Camellia sinensis | China | KJ955109 | KM023257 | KJ955257 | MZ799256 | KJ954810 | KJ954524 |
| C. hippeastri | CBS 125376 * | Hippeastrum vittatum | China | JQ005231 | JQ005579 | JQ005665 | JQ005405 | - | - |
| C. hippeastri | CBS 241.78 | Hippeastrum vittatum | China | JX010293 | JX009485 | JQ005666 | JX009838 | - | - |
| C. horii | ICMP 10492 * | Diospyros kaki | Japan | GQ329690 | JX009438 | JX010450 | JX009752 | GQ329681 | JQ807840 |
| C. hystricis | CPC 28153; CBS 142411 * | Citrus hystrix | Italy | KY856450 | KY856023 | KY856532 | KY856190 | KY856274 | - |
| C. jiangxiense | LF687 *, CGMCC 3.17361 | Camellia sinensis | China | KJ955201 | KJ954471 | KJ955348 | MZ799257 | KJ954902 | KJ954607 |
| C. kahawae | IMI 319418; ICMP 17816 * | Coffea arabica | Kenya | JX010231 | JX009452 | JX010444 | JX009813 | JX010012 | JQ894579 |
| C. karstii | CBS 126532 | Citrus sp. | South Africa | JQ005209 | JQ005557 | JQ005643 | JQ005383 | - | - |
| C. karstii | CBS 129833 | Musa sp. | Mexico | JQ005175 | JQ005523 | JQ005609 | JQ005349 | - | - |
| C. karstii | VPRI 43652; UMC016 | Citrus sinensis | Australia | MW081179 | MW081187 | MW081183 | MW081191 | - | - |
| C. karstii | PPDU41K | Coffea arabica | Saudi Arabia | OR048754 | OR050665 | OR050762 | OR050717 | - | - |
| C. limonicola | CBS 142410; CPC 31141 * | Citrus limon | Malta | KY856472 | KY856045 | KY856554 | KY856213 | - | - |
| C. makassarense | CBS 143664, CPC 28612, CPC 28556 | Capsicum annuum | Indonesia | MH728812 | MH781477 | MH846560 | MH805847 | MH728821 | MH728831 |
| C. musae | ICMP 19119; CBS 116870 * | Musa sp. | USA | JX010146 | JX009433 | HQ596280 | JX009896 | JX010050 | KC888926 |
| C. nanhuaense | YMF1.04993 | Unknown | China | OK030870 | OK513609 | OK513639 | OK513569 | OK513673 | - |
| C. novae-zelandiae | CBS 128505 * | Capsicum annuum | New Zealand | JQ005228 | JQ005576 | JQ005662 | JQ005402 | - | - |
| C. noveboracense | AFKH109 | Malus domestica | USA | MN646685 | MN640565 | MN640569 | MN640567 | MN640564 | |
| C. nullisetosum | YMF1.06946 | Unknown | China | OK030872 | OK513611 | OK513641 | OK513571 | OK513675 | |
| C. nupharicola | ICMP 18187 * | Nuphar polysepala | USA | JX010187 | JX009437 | JX010398 | JX009835 | JX009972 | JX145319 |
| C. oblongisporum | YMF1.06938 | Unknown | China | OK030874 | OK513643 | OK513573 | OK513677 | - | |
| C. oncidii | CBS 129828 * | Oncidium sp. | Germany | JQ005169 | JQ005517 | JQ005603 | JQ005343 | - | - |
| C. pandanicola | MFLUCC 17-0571 | Pandanaceae | Thailand | MG646967 | MG646938 | MG646926 | MG646931 | MG646934 | - |
| C. pandanicola | SAUCC200204 | Unknown | China | MW786641 | MW883694 | MW888969 | MW883685 | MW846239 | - |
| C. pandanicola | SAUCC201152 | Unknown | China | MW786746 | MW883702 | MW888977 | MW883693 | MW876478 | - |
| C. parsonsiae | CBS 128525 * | Parsonsia capsularis | New Zealand | JQ005233 | JQ005581 | JQ005667 | JQ005407 | - | - |
| C. parvisporum | YMF1.06942 | Unknown | China | OK030876 | OK513613 | OK513645 | OK513575 | OK513679 | - |
| C. perseae | CBS 141365 *, GA100, GA 170 | Persea americana | Israel | KX620308 | KX620145 | KX620341 | MZ799260 | KX620242 | KX620180 |
| C. petchii | CBS 378.94 * | Dracaena marginata | Italy | JQ005223 | JQ005571 | JQ005657 | JQ005397 | - | - |
| C. phyllanthi | CBS 175.67 * | Phyllanthus acidus | India | JQ005221 | JQ005569 | JQ005655 | JQ005395 | - | - |
| C. phyllanthi | PPDU36S | Coffea arabica | Saudi Arabia | OR048762 | OR050673 | OR050770 | OR050725 | - | - |
| C. proteae | CBS 132882 * | Protea sp. | South Africa | KC297079 | KC296940 | KC297101 | KC296986 | KC297009 | - |
| C. pseudotheobromicola | MFLUCC 18–1602 | Prunus avium | China | MH817395 | MH853681 | MH853684 | MH853678 | MH853675 | - |
| C. psidii | ICMP 19120 * | Psidium sp. | Italy | JX010219 | JX009515 | JX010443 | JX009901 | JX009967 | KC888931 |
| C. queenslandicum | ICMP 1778 * | Carica papaya | Australia | JX010276 | JX009447 | JX010414 | JX009899 | JX009934 | KC888928 |
| C. rhexiae | Coll1026, CBS 133134 * | Rhexia virginica | USA | JX145128 | MZ664127 | JX145179 | MZ799258 | MZ664046 | JX145290 |
| C. salsolae | ICMP 19051 * | Salsola tragus | Hungary | JX010242 | JX009562 | JX010403 | JX009863 | JX009916 | KC888925 |
| C. saudianum | PPDU28C | Coffea arabica | Saudi Arabia | OR048774 | OR050685 | OR050782 | OR050737 | OR050755 | OR050710 |
| C. saudianum | PPDU28E | Coffea arabica | Saudi Arabia | OR048773 | OR050684 | OR050781 | OR050736 | OR050754 | OR050709 |
| C. saudianum | PPDU28J | Coffea arabica | Saudi Arabia | OR048772 | OR050683 | OR050780 | OR050735 | OR050753 | OR050708 |
| C. saudianum | PPDU28L | Coffea arabica | Saudi Arabia | OR048771 | OR050682 | OR050779 | OR050734 | OR050752 | OR050707 |
| C. saudianum | PPDU29A | Coffea arabica | Saudi Arabia | OR048770 | OR050681 | OR050778 | OR050733 | OR050751 | OR050706 |
| C. saudianum | PPDU29B | Coffea arabica | Saudi Arabia | OR048769 | OR050680 | OR050777 | OR050732 | OR050750 | OR050705 |
| C. saudianum | PPDU31I | Coffea arabica | Saudi Arabia | OR048766 | OR050677 | OR050774 | OR050729 | OR050747 | OR050702 |
| C. saudianum | PPDU31M | Coffea arabica | Saudi Arabia | OR048765 | OR050676 | OR050773 | OR050728 | OR050746 | OR050701 |
| C. saudianum | PPDU38B | Coffea arabica | Saudi Arabia | OR048761 | OR050672 | OR050769 | OR050724 | - | OR050698 |
| C. saudianum | PPDU38F | Coffea arabica | Saudi Arabia | OR048760 | OR050671 | OR050768 | OR050723 | - | OR050697 |
| C. saudianum | PPDU38H* | Coffea arabica | Saudi Arabia | OR048759 | OR050670 | OR050767 | OR050722 | - | OR050696 |
| C. saudianum | PPDU38I | Coffea arabica | Saudi Arabia | OR048758 | OR050669 | OR050766 | OR050721 | - | OR050695 |
| C. siamense | VPRI 43077; UMC004 | Citrus limon | Australia | MG572140 | MK473453 | MG572151 | MW091989 | MG572129 | MG572173 |
| C. siamense | CPC 30209, UOM 13 | Capsicum annuum | Indonesia | MH707471 | MH781464 | MH846547 | MH805834 | MH707452 | MH713897 |
| C. siamense | CPC 30210, UOM14 | Capsicum annuum | Indonesia | MH707472 | MH781465 | MH846548 | MH805835 | MH707453 | MH713896 |
| C. siamense | CPC 30212, UOM16 | Capsicum annuum | Indonesia | MH707474 | MH781467 | MH846550 | MH805837 | MH707455 | MH713894 |
| C. siamense | CPC 30221, UOM25 | Capsicum annuum | Thailand | MH707475 | MH781468 | MH846551 | MH805838 | MH707456 | MH713893 |
| C. siamense | CPC 30222, UOM26 | Capsicum annuum | Thailand | MH707476 | MH781469 | MH846552 | MH805839 | MH707457 | MH713892 |
| C. siamense | CPC 30223, UOM27 | Capsicum annuum | Indonesia | MH707477 | MH781470 | MH846553 | MH805840 | MH707458 | MH713891 |
| C. siamense | ICMP 18578 CBS 130417 * | Coffea arabica | Thailand | JX010171 | FJ907423 | JX010404 | JX009865 | JX009924 | JQ899289 |
| C. siamense | BRIP 54270b; VPRI 43029; A10-43029 | Citrus australasica | Australia | MK469995 | MK470085 | MK470049 | MW091971 | MK470013 | MK470067 |
| C. siamense syn. C. endomangiferae | CMM 3814a | Mangifera indica | Brazil | KC702994 | KC702922 | KM404170 | KC598113 | KC702955 | KJ155453 |
| C. siamense syn. C. hymenocallidis | CBS 125378, ICMP 18642, LC0043a | Hymenocallis americana | China | JX010278 | JX009441 | JX010410 | GQ856730 | JX010019 | JQ899283 |
| C. siamense syn. C. hymenocallidis | CBS 112983, CPC 2291 | Protea cynaroides | Zimbabwe | KC297065 | KC296929 | KC297100 | KC296984 | KC297007 | KP703761 |
| C. siamense syn. C. hymenocallidis | CBS 113199. CPC 2290 | Protea cynaroides | Zimbabwe | KC297066 | KC296930 | KC297090 | KC296985 | KC297008 | KP703763 |
| C. siamense syn. C. hymenocallidis | CBS 116868 | Protea cynaroides | Zimbabwe | KC566815 | KC566961 | KP703429 | KC566382 | KC566669 | KP703764 |
| C. siamense syn. C. jasmini-sambac | CBS 130420; ICMP 19118 | Jasminum sambac | Viet Nam | HM131511 | HM131507 | JX010415 | JX009895 | HM131497 | JQ807841 |
| C. siamense | PPDU26A | Coffea arabica | Saudi Arabia | OR048780 | OR050691 | OR050788 | OR050743 | OR050761 | OR050716 |
| C. siamense | PPDU27B | Coffea arabica | Saudi Arabia | OR048778 | OR050689 | OR050786 | OR050741 | OR050759 | OR050714 |
| C. siamense | PPDU27M | Coffea arabica | Saudi Arabia | OR048776 | OR050687 | OR050784 | OR050739 | OR050757 | OR050712 |
| C. siamense | PPDU29H | Coffea arabica | Saudi Arabia | OR048767 | OR050678 | OR050775 | OR050730 | OR050748 | OR050703 |
| C. siamense | PPDU32B | Coffea arabica | Saudi Arabia | OR048763 | OR050674 | OR050771 | OR050726 | OR050744 | OR050699 |
| C. siamense | PPDU39D | Coffea arabica | Saudi Arabia | OR048757 | OR050668 | OR050765 | OR050720 | - | OR050694 |
| C. siamense | PPDU39E | Coffea arabica | Saudi Arabia | OR048756 | OR050667 | OR050764 | OR050719 | - | OR050693 |
| C. siamense | PPDU40G | Coffea arabica | Saudi Arabia | OR048755 | OR050666 | OR050763 | OR050718 | - | OR050692 |
| C. subhenanense | YMF1.06865 | Unknown | China | OK030883 | OK513618 | OK513647 | OK513581 | OK513684 | - |
| C. syzygicola | DNCL021; MFLUCC 10-0624 *, DU-2013c | Syzygium samarangense | Thailand | KF242094 | KF157801 | KF254880 | KJ947226 | KF242156 | KP743473 |
| C. tainanense | UOM 1119, Coll 1290 | Capsicum annuum | Taiwan | MH728805 | MH781487 | MH846570 | MH805857 | MH728819 | MH728824 |
| C. tainanense | CBS 143666, CPC30245, | Capsicum annuum | Taiwan | MH728818 | MH781475 | MH846558 | MH805845 | MH728823 | MH728836 |
| C. temperatum | CBS 133122 * | Vaccinium macrocarpon | USA | JX145159 | MZ664125 | JX145211 | MZ799254 | MZ664045 | JX145298 |
| C. theobromicola | ICMP 18649; CBS 124945 * | Theobroma cacao | Panama | JX010294 | JX009444 | JX010447 | JX009869 | JX010006 | KC790726 |
| C. ti | ICMP 4832 * | Cordyline sp. | New Zealand | JX010269 | JX009520 | JX010442 | JX009898 | JX009952 | KM360146 |
| C. torulosum | CBS 128544 * | Solanum melongena | New Zealand | JQ005164 | JQ005512 | JQ005598 | JQ005338 | - | - |
| C. tropicale | ICMP 18653; CBS 124949 * | Theobroma cacao | Panama | JX010264 | JX009489 | JX010407 | JX009870 | JX010007 | KC790728 |
| C. truncatum | CBS 151.35 * | Phaseolus lunatus | USA | GU227862 | GU227960 | GU228156 | GU228352 | - | - |
| C. truncatum | CBS 151.35 * | Phaseolus lunatus | USA | GU227862 | GU227960 | GU228156 | GU228352 | - | - |
| C. viniferum | GZAAS 5.08601; GC9 | Vitis vinifera | China | JN412804 | JN412795 | JN412813 | MW684718 | JN412798 | MT648530 |
| C. watphraense | MFLUCC 14-0123 | Dendrobium sp. | Thailand | MF448523 | MH376384 | MH351276 | - | - | - |
| C. wuxiense | CGMCC 3.17894 * | Camellia sinensis | China | KU251591 | KU251672 | KU252200 | KU251939 | KU252045 | KU251722 |
| C. xanthorrhoeae | BRIP 45094; ICMP 17903; CBS 127831 * | Xanthorrhoea sp. | Australia | JX010261 | JX009478 | JX010448 | JX009823 | JX009927 | KC790689 |
| C. xishuangbannaense | MFLUCC 19-0107 | Magnolia liliifera | China | MW346469 | MW652294 | - | MW660832 | MW537586 | - |
| C. yuanjiangensis | YMF1.04996 | Unknown | China | OK030885 | OK513620 | OK513649 | OK513583 | OK513686 | - |
| C. yulongense | CFCC 50818 | Vaccinium dunalianum | China | MH751507 | MH777394 | MK108987 | MH793605 | MK108986 | - |
| Colletotrichum sp. | CBS 123921, MAFF 238642 | Dendrobium kingianum | Japan | JQ005163 | JQ005511 | JQ005597 | JQ005337 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhudaib, K.; Ismail, A.M.; Magistà, D. Multi-Locus Phylogenetic Analysis Revealed the Association of Six Colletotrichum Species with Anthracnose Disease of Coffee (Coffea arabica L.) in Saudi Arabia. J. Fungi 2023, 9, 705. https://doi.org/10.3390/jof9070705
Alhudaib K, Ismail AM, Magistà D. Multi-Locus Phylogenetic Analysis Revealed the Association of Six Colletotrichum Species with Anthracnose Disease of Coffee (Coffea arabica L.) in Saudi Arabia. Journal of Fungi. 2023; 9(7):705. https://doi.org/10.3390/jof9070705
Chicago/Turabian StyleAlhudaib, Khalid, Ahmed Mahmoud Ismail, and Donato Magistà. 2023. "Multi-Locus Phylogenetic Analysis Revealed the Association of Six Colletotrichum Species with Anthracnose Disease of Coffee (Coffea arabica L.) in Saudi Arabia" Journal of Fungi 9, no. 7: 705. https://doi.org/10.3390/jof9070705