
Unprecedented Antimicrobial and Cytotoxic Polyketides from Cultures of Diaporthe africana sp. nov.
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
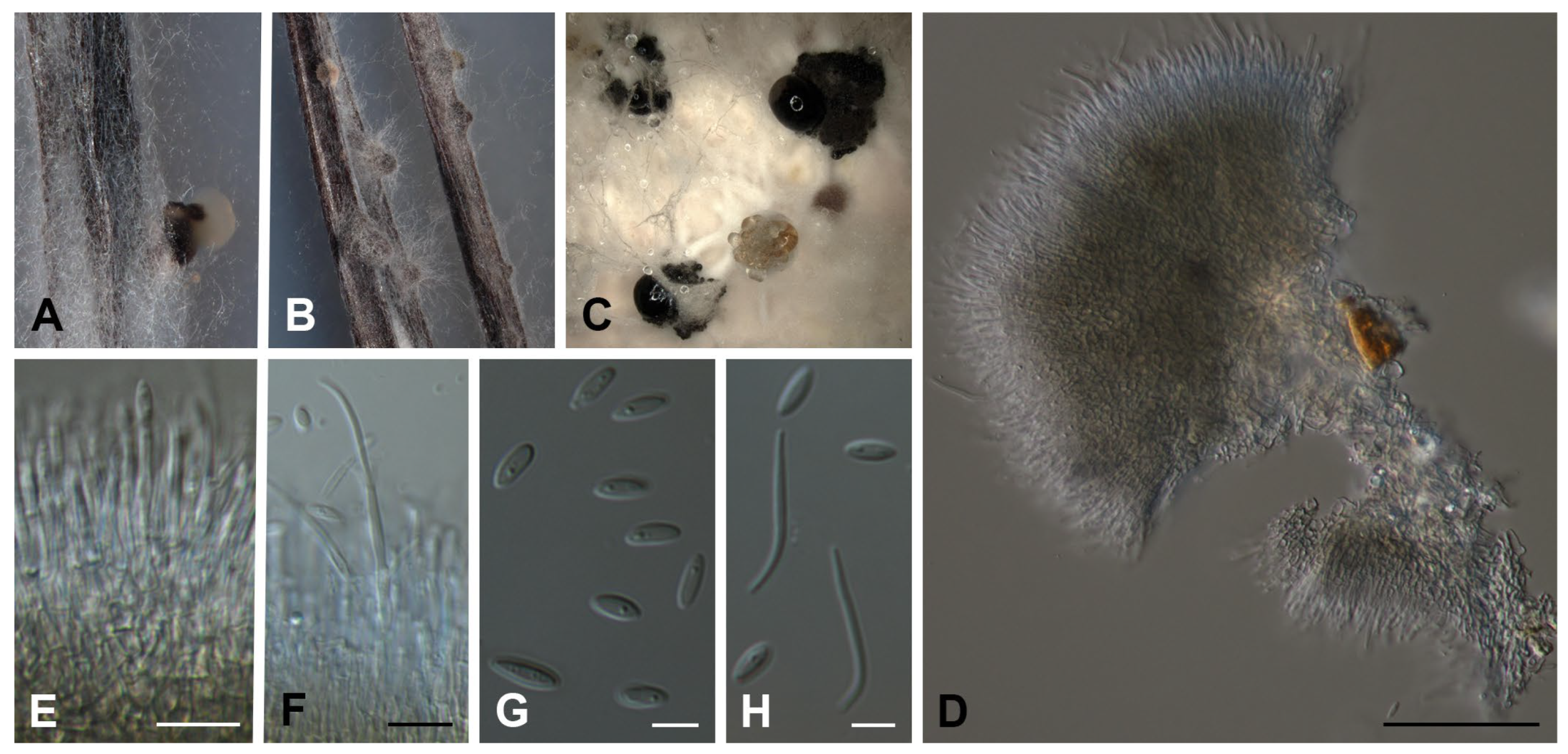
2.2. Fungal Isolation and Identification
2.3. Fermentation and Extraction
2.4. Isolation of Compounds 1–8
2.5. Determination of Amino Acid Stereochemistry
2.6. Antimicrobial and Cytotoxic Activities
3. Results and Discussion
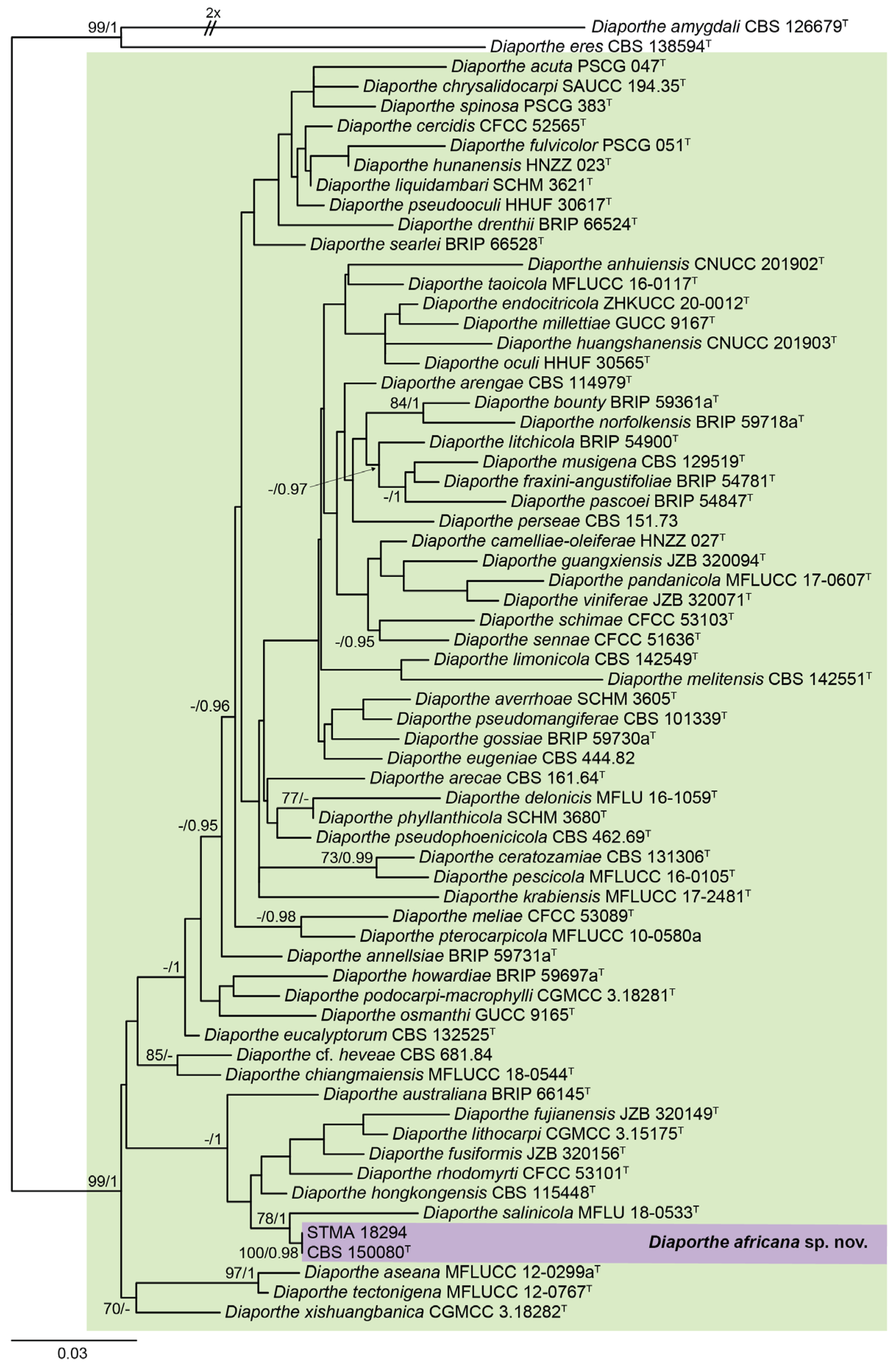
3.1. Molecular Phylogeny and Taxonomy
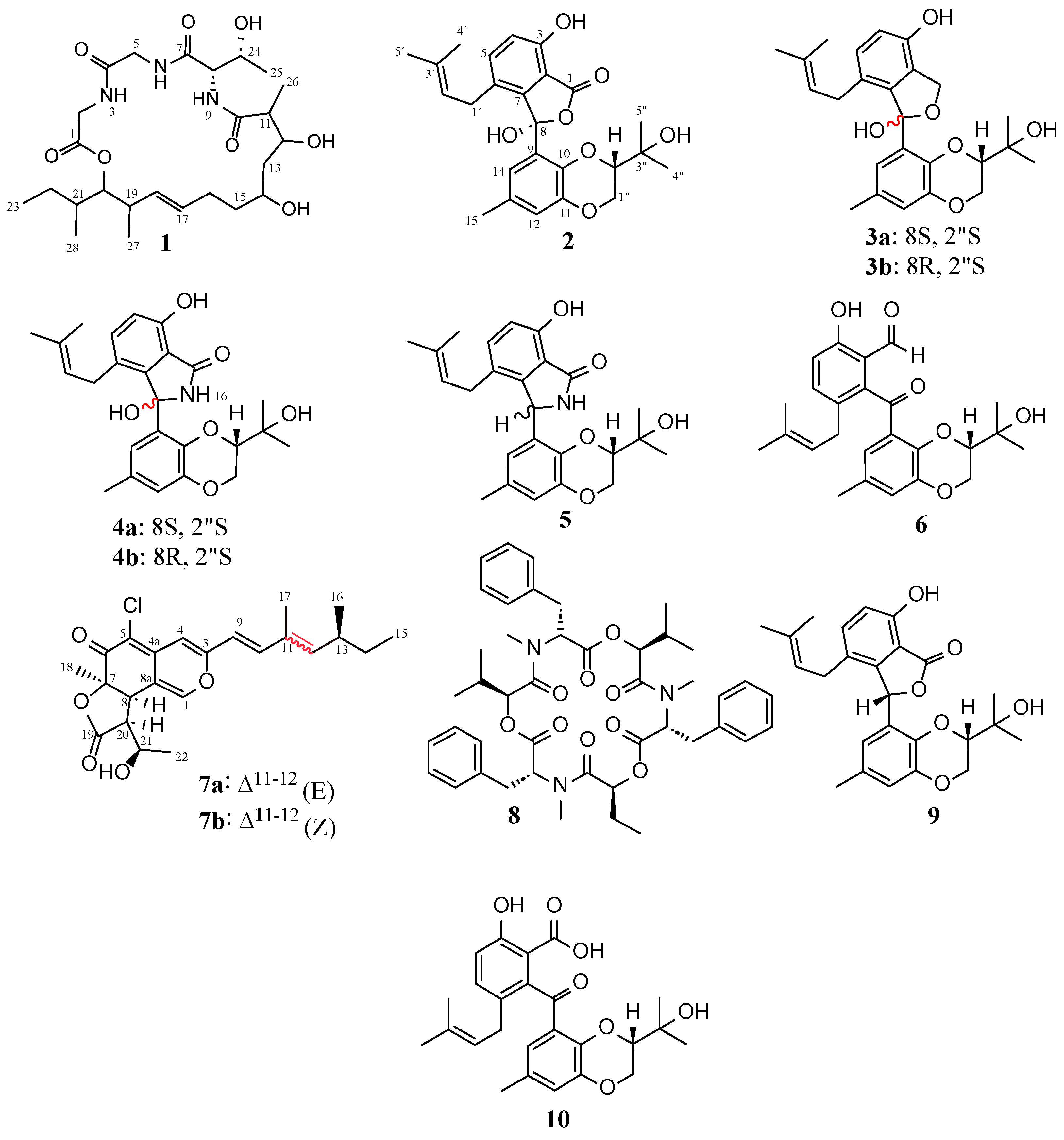
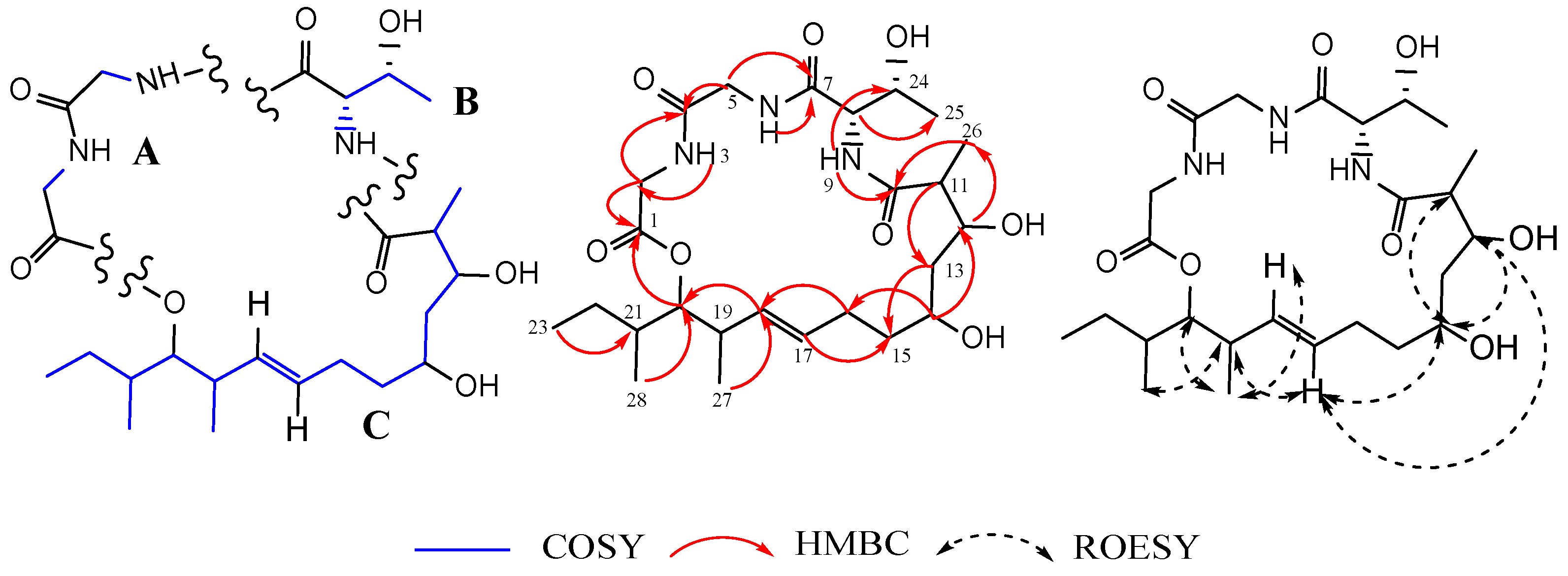
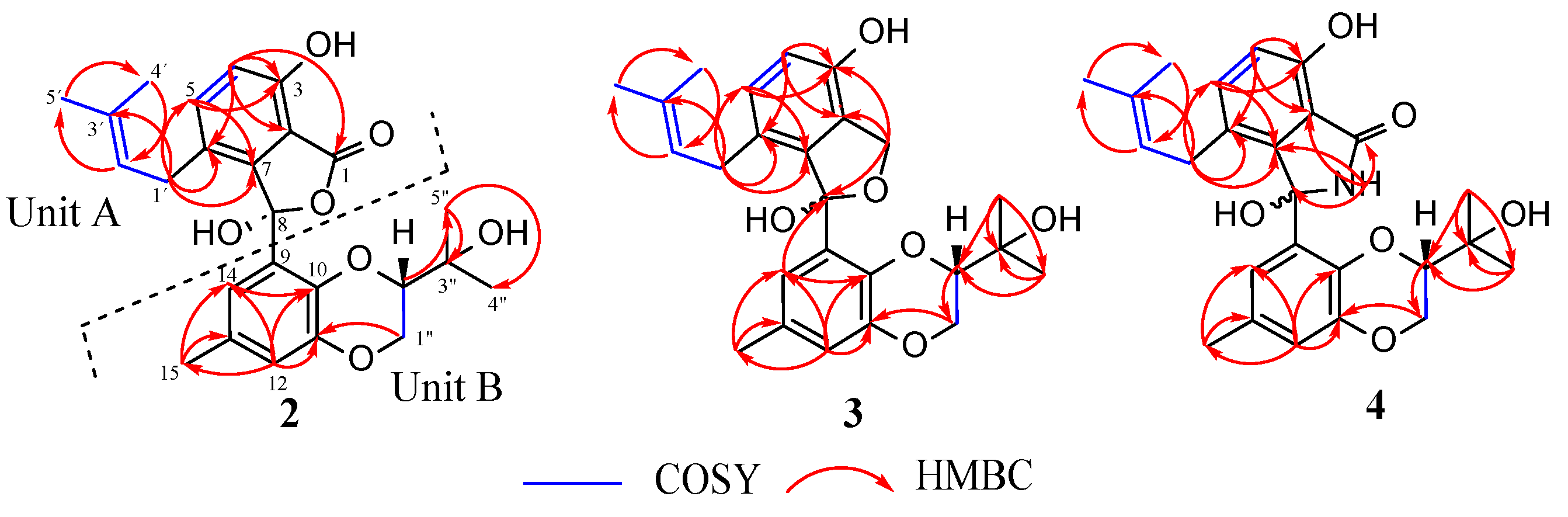
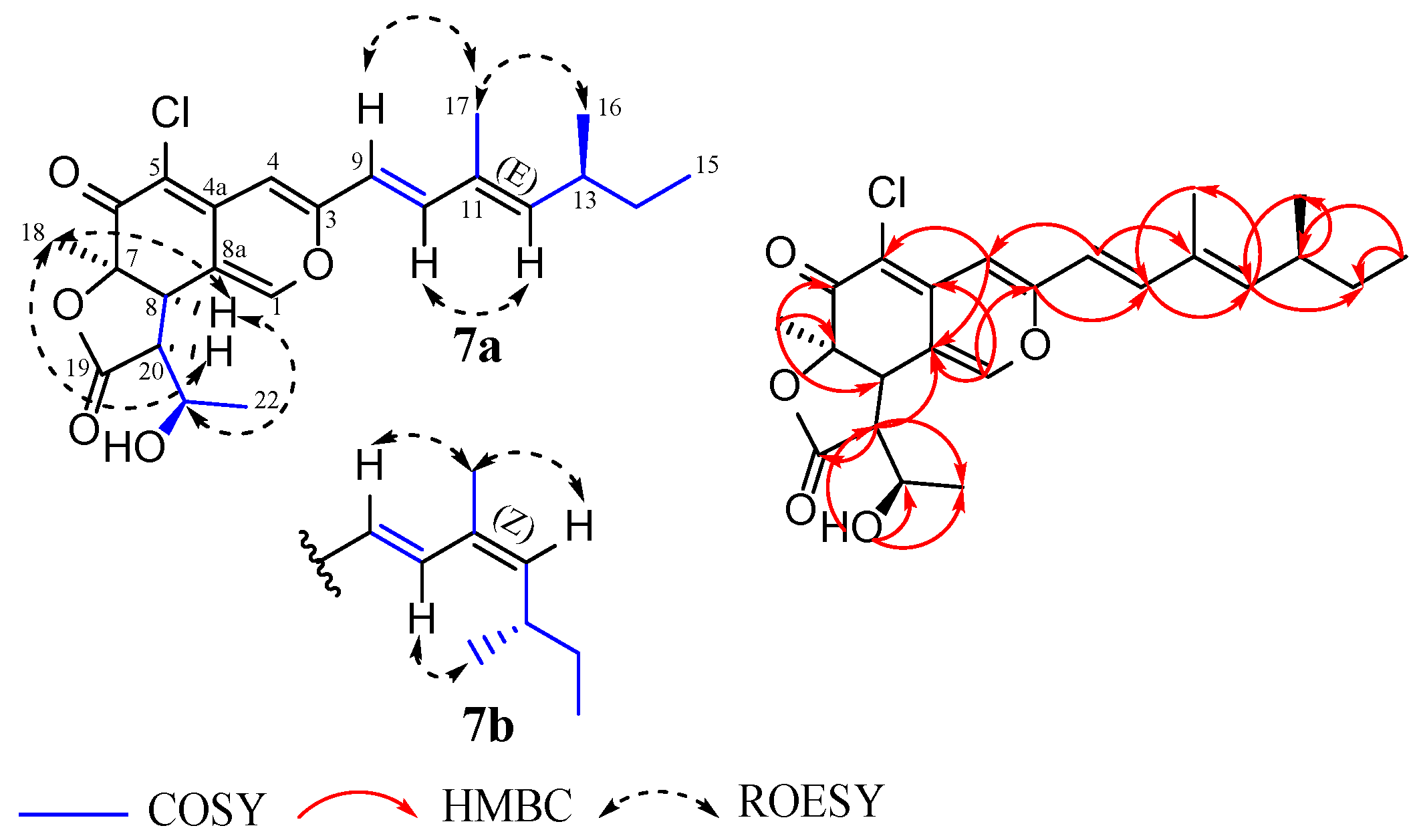
3.2. Structural Elucidation of Compounds 1–8
3.3. Physicochemical Data for Compounds 1–4, 7
3.4. Biological Activities of Compounds 1–8
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, T.C.; Lu, Y.H.; Wang, J.F.; Song, Z.Q.; Hou, Y.G.; Liu, S.S.; Liu, C.S.; Wu, S.H. Bioactive secondary metabolites of the genus Diaporthe and anamorph Phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 2021, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Chepkirui, C.; Stadler, M. The genus Diaporthe: A rich source of diverse and bioactive metabolites. Mycol. Prog. 2017, 16, 477–494. [Google Scholar] [CrossRef]
- Catalano, A.; Iacopetta, D.; Ceramella, J.; Scumaci, D.; Giuzio, F.; Saturnino, C.; Aquaro, S.; Rosano, C.; Sinicropi, M.S. Multidrug resistance (MDR): A widespread phenomenon in pharmacological therapies. Molecules 2022, 27, 616. [Google Scholar] [CrossRef] [PubMed]
- Hilário, S.; Gonçalves, M.F.M. Endophytic Diaporthe as promising leads for the development of biopesticides and biofertilizers for a sustainable agriculture. Microorganisms 2022, 10, 2453. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Khan, B.; Dai, Q.; Lin, J.; Kang, L.; Rajput, N.A.; Yan, W.; Liu, G. Potential of secondary metabolites of Diaporthe species associated with terrestrial and marine origins. J. Fungi 2023, 9, 453. [Google Scholar] [CrossRef] [PubMed]
- Riga, R.; Happyana, N.; Quentmeier, A.; Zammarelli, C.; Kayser, O.; Hakim, E.H. Secondary metabolites from Diaporthe lithocarpus isolated from Artocarpus heterophyllus. Nat. Prod. Res. 2021, 35, 2324–2328. [Google Scholar] [CrossRef]
- Niaz, S.; Khan, D.; Naz, R.; Safdar, K.; Abidin, S.Z.U.; Khan, I.U.; Gul, R.; Khan, W.U.; Khan, M.A.U.; Lan, L. Antimicrobial and antioxidant chlorinated azaphilones from mangrove Diaporthe perseae sp. isolated from the stem of Chinese mangrove Pongamia pinnata. J. Asian Nat. Prod. Res. 2021, 23, 1077–1084. [Google Scholar] [CrossRef]
- Yang, X.-W.; Salome, C.; Rongbiao Pi, S.; Zhou, X.; Liu, S.; Luo, X.; Yang, J.; Chen, F.; Lin, X.; Chen, C.; et al. Structurally diverse polyketides from the mangrove-derived fungus Diaporthe sp. SCSIO 41011 with their anti-influenza A virus activities. Front. Chem. 2018, 1, 282. [Google Scholar] [CrossRef] [Green Version]
- Matio Kemkuignou, B.; Schweizer, L.; Lambert, C.; Gisèle, E.; Anoumedem, M.; Kouam, S.F.; Stadler, M.; Marin-Felix, Y.; Felix, Y.M. New polyketides from the liquid culture of Diaporthe breyniae sp. nov. (Diaporthales, Diaporthaceae). MycoKeys 2022, 90, 85–118. [Google Scholar] [CrossRef]
- Smith, H.; Wingfield, M.J.; Crous, P.W.; Coutinho, I.A. Sphaeropsis sapinea and Botryosphaeria dothidea endophytic in Pinus spp. and Eucalyptus spp. in South Africa. S. Afr. J. Bot. 1996, 62, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Verkley, G.J.M.; Groenewald, J.Z.; Samson, R.A. Fungal Biodiversity. CBS Laboratory Manual Series 1; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2009. [Google Scholar]
- The Royal Horticultural Society London. R.H.S. Colour Chart; The Royal Horticultural Society London: London, UK, 1966. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Ayres, D.L.; Darling, A.; Zwickl, D.J.; Beerli, P.; Holder, M.T.; Lewis, P.O.; Huelsenbeck, J.P.; Ronquist, F.; Swofford, D.L.; Cummings, M.P.; et al. BEAGLE: An application programming interface and high-performance computing library for statistical phylogenetics. Syst. Biol. 2012, 61, 170–173. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Hou, C. Three new species of Diaporthe from China based on morphological characters and DNA sequence data analyses. Phytotaxa 2019, 422, 157–174. [Google Scholar] [CrossRef]
- Tan, Y.P.; Shivas, R.G. Nomenclatural novelties. Index Aust. Fungi 2022, 2, 1–12. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Wrona, C.J.; Mohankumar, V.; Schoeman, M.H.; Tan, Y.P.; Shivas, R.G.; Jeff-Ego, O.S.; Akinsanmi, O.A. Phomopsis husk rot of macadamia in Australia and South Africa caused by novel Diaporthe species. Plant Pathol. 2020, 69, 911–921. [Google Scholar] [CrossRef]
- Chang, C.Q.; Cheng, Y.H.; Xiang, M.M.; Jiang, Z.D. New species of Phomopsis on woody plants in Fujian Province. Mycosystema 2005, 24, 6–11. [Google Scholar]
- Yang, Q.; Tang, J.; Zhou, G.Y. Characterization of Diaporthe species on Camellia oleifera in Hunan Province, with descriptions of two new species. MycoKeys 2021, 84, 15–33. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Romberg, M.; Mel’nik, V.A.; Verkley, G.J.M.; Groenewald, J.Z. Fungal planet description sheets: 92–106. Persoonia 2011, 27, 130–162. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China, with twelve new species described. MycoKeys 2018, 39, 97–149. [Google Scholar] [CrossRef] [Green Version]
- de Silva, N.; Hyde, K.D.; Lumyong, S.; Phillips, A.J.L.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Tennakoon, D.S.; Suwannarach, N.; Karunarathna, S.C. Morphology, phylogeny, host association and geography of fungi associated with plants of Annonaceae, Apocynaceae and Magnoliaceae. Mycosphere 2022, 13, 955–1076. [Google Scholar] [CrossRef]
- Huang, S.; Xia, J.; Zhang, X.; Sun, W. Morphological and phylogenetic analyses reveal three new species of Diaporthe from Yunnan, China. MycoKeys 2021, 78, 49–77. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 3621. [Google Scholar] [CrossRef] [PubMed]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Burgess, T.I.; Decock, C.A.; Dreyer, L.L.; Granke, L.L.; Guest, D.I.; Hardy, G.E.S.J.; Hausbeck, M.K.; et al. Fungal Planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef]
- Tan, Y.P.; Edwards, J.; Grice, K.R.E.; Shivas, R.G. Molecular phylogenetic analysis reveals six new species of Diaporthe from Australia. Fungal Divers. 2013, 61, 251–260. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Jayawardena, R.S.; Li, H.L.; Zhou, Y.Y.; Zhang, W.; Phillips, A.J.L.; Wanasinghe, D.N.; Dissanayake, A.J.; Li, X.H.; Li, Y.H.; et al. Microfungi associated with Camellia sinensis: A case study of leaf and shoot necrosis on Tea in Fujian, China. Mycosphere 2021, 12, 430–518. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Dissanayake, A.J.; Li, X.; Liu, M.; Wanasinghe, D.N.; Xu, J.; Zhao, W.; Zhang, W.; Zhou, Y.; Hyde, K.D.; et al. High genetic diversity and species complexity of Diaporthe Associated with grapevine dieback in China. Front. Microbiol. 2019, 10, 1936. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.H.; Sun, W.; Su, Y.Y.; Cai, L. Three new species of Phomopsis in Gutianshan Nature Reserve in China. Mycol. Prog. 2014, 13, 111–121. [Google Scholar] [CrossRef]
- Cao, L.; Luo, D.; Lin, W.; Yang, Q.; Deng, X. Four new species of Diaporthe (Diaporthaceae, Diaporthales) from forest plants in China. MycoKeys 2022, 91, 25–47. [Google Scholar] [CrossRef]
- Long, H.; Zhang, Q.; Hao, Y.-Y.; Shao, X.-Q.; Wei, X.-X.; Hyde, K.D.; Wang, Y.; Zhao, D.-G. Diaporthe species in south-western China. MycoKeys 2019, 57, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, K.; Mochizuki, K.; Takagi, D.; Ishida, K.; Sunada, A.; Ohkusu, K.; Kamei, K.; Hashimoto, A.; Tanaka, K. Identification and antifungal sensitivity of two new species of Diaporthe isolated. J. Infect. Chemother. 2019, 25, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Tibpromma, S.; Hyde, K.D.; Bhat, J.D.; Mortimer, P.E.; Xu, J.; Promputtha, I.; Doilom, M.; Yang, J.B.; Tang, A.M.C.; Karunarathna, S.C. Identification of endophytic fungi from leaves of Pandanaceae based on their morphotypes and DNA sequence data from southern Thailand. MycoKeys 2018, 33, 25–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dissanayake, A.J.; Zhang, W.; Liu, M.; Hyde, K.D.; Zhao, W.S.; Li, X.H.; Yan, J.Y. Diaporthe species associated with peach tree dieback in Hubei, China. Mycosphere 2017, 8, 533–549. [Google Scholar] [CrossRef]
- Gao, Y.; Duan, W.; Crous, P.W.; Cai, L. Diaporthe is paraphyletic. IMA Fungus 2017, 8, 153–187. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. Multi-locus phylogeny reveals three new species of Diaporthe from Thailand. Cryptogam. Mycol. 2012, 33, 295–309. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, N.; Tian, C.M. New species and records of Diaporthe from Jiangxi Province, China. MycoKeys 2021, 77, 41–64. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Du, Z.; Tian, C.M. Diaporthe species occurring on Senna bicapsularis in southern China, with descriptions of two new species. Phytotaxa 2017, 302, 145–155. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2017, 82, 107–182. [Google Scholar] [CrossRef]
- Becker, K.; Wongkanoun, S.; Wessel, A.C.; Bills, G.F.; Stadler, M.; Luangsa-Ard, J.J. Phylogenetic and chemotaxonomic studies confirm the affinities of Stromatoneurospora phoenix to the Coprophilous Xylariaceae. J. Fungi 2020, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Harms, K.; Surup, F.; Stadler, M.; Stchigel, A.M.; Marin-Felix, Y. Morinagadepsin, a depsipeptide from the fungus Morinagamyces vermicularis gen. et comb. nov. Microorganisms 2021, 9, 1191. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Wessel, A.C.; Luangsa-Ard, J.J.; Stadler, M. Viridistratins A–C, Antimicrobial and Cytotoxic Benzo[j]fluoranthenes from Stromata of Annulohypoxylon viridistratum (Hypoxylaceae, Ascomycota). Biomolecules 2020, 10, 805. [Google Scholar] [CrossRef] [PubMed]
- Mulwa, L.S.; Jansen, R.; Praditya, D.F.; Mohr, K.I.; Okanya, P.W.; Wink, J.; Steinmann, E.; Stadler, M. Lanyamycin, a macrolide antibiotic from Sorangium cellulosum, strain Soce 481 (Myxobacteria). Beilstein J. Org. Chem. 2018, 14, 1554. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Udayanga, D.; Wang, X.; Hou, X.; Mei, X.; Fu, Y.; Hyde, K.D.; Li, H. Endophytic Diaporthe associated with Citrus: A phylogenetic reassessment with seven new species from China. Fungal Biol. 2015, 119, 331–347. [Google Scholar] [CrossRef]
- Erper, I.; Turkkan, M.; Ozcan, M.; Luongo, L.; Belisario, A. Characterization of Diaporthe hongkongensis species causing stem-end rot on kiwifruit in Turkey. J. Plant Pathol. 2017, 99, 779–782. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Tsai, I.; Wang, J.Y.; Withee, P.; Tanjira, M.; Lin, S.R.; Suwannarach, N.; Kumla, J.; Elgorban, A.M.; Cheewangkoon, R. Molecular phylogenetic diversity and biological characterization of Diaporthe species associated with leaf spots of Camellia sinensis in Taiwan. Plants 2021, 10, 1434. [Google Scholar] [CrossRef]
- Du, Y.; Wang, X.; Guo, Y.; Xiao, F.; Peng, Y.; Hong, N.; Wang, G.; Du, Y.; Wang, X.; Guo, Y.; et al. Biological and molecular characterization of seven Diaporthe species associated with kiwifruit shoot blight and leaf spot in China. Phytopathol. Mediterr. 2021, 60, 177–198. [Google Scholar] [CrossRef]
- Cui, H.; Lin, Y.; Luo, M.; Lu, Y.; Huang, X.; She, Z. Diaporisoindoles A–C: Three isoprenylisoindole alkaloid derivatives from the mangrove endophytic fungus Diaporthe sp. SYSU-HQ3. Org. Lett. 2017, 19, 5621–5624. [Google Scholar] [CrossRef]
- Luo, X.; Lin, X.; Tao, H.; Wang, J.; Li, J.; Yang, B.; Zhou, X.; Liu, Y. Isochromophilones A–F, cytotoxic chloroazaphilones from the marine mangrove endophytic fungus Diaporthe sp. SCSIO 41011. J. Nat. Prod. 2018, 81, 934–941. [Google Scholar] [CrossRef]
- Zhang, C.; Ondeyka, J.G.; Herath, K.B.; Guan, Z.; Collado, J.; Platas, G.; Pelaez, F.; Leavitt, P.S.; Gurnett, A.; Nare, B.; et al. Tenellones A and B from a Diaporthe sp.: Two highly substituted benzophenone inhibitors of parasite cGMP-dependent protein kinase activity. J. Nat. Prod. 2005, 68, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. The structure of beauvericin, a new depsipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 1969, 10, 4255–4258. [Google Scholar] [CrossRef]
- Ola, A.R.B.; Aly, A.H.; Lin, W.; Wray, V.; Debbab, A. Structural revision and absolute configuration of lateritin. Tetrahedron Lett. 2014, 55, 6184–6187. [Google Scholar] [CrossRef]
- Caloni, F.; Fossati, P.; Anadón, A.; Bertero, A. Beauvericin: The beauty and the beast. Environ. Toxicol. Pharmacol. 2020, 75, 103349. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.Q.; Du, S.T.; Xiao, J.; Wang, D.C.; Han, W.B.; Zhang, Q.; Gao, J.M. Isolation and characterization of antifungal metabolites from the Melia azedarach-Associated Fungus Diaporthe eucalyptorum. J. Agric. Food Chem. 2020, 68, 2418–2425. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Liu, Y.; Li, J.; Huang, X.; Yan, T.; Cao, W.; Liu, H.; Long, Y.; She, Z. Diaporindenes A–D: Four unusual 2,3-dihydro-1H-indene analogues with anti-inflammatory activities from the mangrove endophytic fungus Diaporthe sp. SYSU-HQ3. J. Org. Chem. 2018, 83, 11804–11813. [Google Scholar] [CrossRef]
- Xu, J.; Liu, Z.; Chen, Y.; Tan, H.; Li, H.; Li, S.; Guo, H.; Huang, Z.; Gao, X.; Liu, H.; et al. Lithocarols A–F, six tenellone derivatives from the deep-sea derived fungus Phomopsis lithocarpus FS508. Bioorg. Chem. 2019, 87, 728–735. [Google Scholar] [CrossRef]
- Xu, J.; Tan, H.; Chen, Y.; Li, S.; Guo, H.; Huang, Z.; Li, H.; Gao, X.; Liu, H.; Zhang, W. Lithocarpinols A and B, a pair of diastereomeric antineoplastic tenellone derivatives from the deep-sea derived fungus Phomopsis lithocarpus FS508. Chin. Chem. Lett. 2019, 30, 439–442. [Google Scholar] [CrossRef]
- Xu, J.; Chen, Y.; Liu, Z.; Li, S.; Wang, Y.; Ren, Y.; Liu, H.; Zhang, W. Lithocarpins E–G, potent anti-tumor tenellone-macrolides from the deep-sea fungus Phomopsis lithocarpus FS508. Chin. J. Chem. 2021, 39, 1104–1112. [Google Scholar] [CrossRef]
- Xu, J.; Tan, H.; Chen, Y.; Li, S.; Huang, Z.; Guo, H.; Li, H.; Gao, X.; Liu, H.; Zhang, W. Lithocarpins A–D: Four tenellone-macrolide conjugated [4 + 2] hetero-adducts from the deep-sea derived fungus Phomopsis lithocarpus FS508. Org. Chem. Front. 2018, 5, 1792–1797. [Google Scholar] [CrossRef]
- Liu, H.B.; Liu, Z.M.; Chen, Y.C.; Tan, H.B.; Li, S.N.; Li, D.L.; Liu, H.X.; Zhang, W.M. Cytotoxic diaporindene and tenellone derivatives from the fungus Phomopsis lithocarpus. Chin. J. Nat. Med. 2021, 19, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Quang, D.N.; Stadler, M.; Fournier, J.; Tomita, A.; Hashimoto, T. Cohaerins C–F, four azaphilones from the xylariaceous fungus Annulohypoxylon cohaerens. Tetrahedron 2006, 62, 6349–6354. [Google Scholar] [CrossRef]
- Panthama, N.; Kanokmedhakul, S.; Kanokmedhakul, K.; Soytong, K. Chemical constituents from the fungus Chaetomium cupreum RY202. Arch. Pharm. Res. 2015, 38, 585–590. [Google Scholar] [CrossRef]
- Omura, S.; Tanaka, H.; Matsuzaki, K.; Ikeda, H.; Masuma, R. Isochromophilones I and II, novel inhibitors against gp120-CD4 binding from Penicillium sp. J. Antibiot. 1993, 46, 1908–1911. [Google Scholar] [CrossRef] [Green Version]
- Whalley, W.B.; Ferguson, G.; Marsh, W.C.; Restivo, R.J. The Chemistry of Fungi. Part LXVIII. The Absolute Configuration of (+)-Sclerotiorin and of the Azaphilones. J. Chem. Soc. Perkin Trans. 1 1976, 13, 1366–1369. [Google Scholar] [CrossRef]
- Chen, C.; Manchand, S.; Gray, R.W.; Whalley, W.B. The chemistry of fungi. Part LXIV. The structure of monascin: The relative stereochemistry of the azaphilones. J. Chem. Soc. C Org. 1969, 3577–3579. [Google Scholar] [CrossRef]
- Steyn, P.S.; Vleggaar, R. The structure of dihydrodeoxy-8-epi-austdiol and the absolute configuration of the azaphilones. J. Chem. Soc. Perkin Trans. 1 1974, 2, 204–206. [Google Scholar] [CrossRef]
- Liu, H.; Xie, H.; Li, C.; Wang, L.; Chen, Q.; Ouyang, X.; Yan, C. Diaporisoindole B reduces lipid accumulation in THP-1 macrophage cells via MAPKS and PPARΓ-LXRᾳ pathways and promotes the reverse cholesterol transport by upregulating SR-B1 and LDLR in HepG2 cells. J. Nat. Prod. 2022, 85, 2769–2778. [Google Scholar] [CrossRef]
- Sood, S.; Sandhu, S.S.; Mukherjee, T.K. Pharmacological and therapeutic potential of beauvericin: A Short Review. J. Proteom. Bioinform. 2017, 10, 18–23. [Google Scholar] [CrossRef]
- Wu, Q.; Patocka, J.; Nepovimova, E.; Kuca, K. A review on the synthesis and bioactivity aspects of beauvericin, a Fusarium mycotoxin. Front. Pharmacol. 2018, 9, 1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolates 1 | GenBank Accession Numbers 2 | References | ||||
|---|---|---|---|---|---|---|---|
| ITS | tub2 | his3 | tef1 | cal | |||
| Diaporthe acuta | PSCG 047T | MK626957 | MK691225 | MK726161 | MK654802 | MK691125 | Guo et al. [23] |
| D. africana | CBS 150080T | OR198681 | OR225229 | OR225231 | OR225227 | OR225233 | Present study |
| STMA 18294 | OR198680 | OR225230 | OR225232 | OR225228 | OR225234 | Present study | |
| D. amygdali | CBS 126679T | KC343022 | KC343990 | KC343506 | KC343748 | KC343264 | Gomes et al. [24] |
| D. anhuiensis | CNUCC 201902T | MN219727 | MN227009 | MN224550 | MN224669 | MN224556 | Zhou and Hou [25] |
| D. annellsiae | BRIP 59731aT | OM918687 | OM960614 | - | OM960596 | - | Tan and Shivas [26] |
| D. arecae | CBS 161.64T | KC343032 | KC344000 | KC343516 | KC343758 | KC343274 | Gomes et al. [24] |
| D. arengae | CBS 114979T | KC343034 | KC344002 | KC343518 | KC343760 | KC343276 | Gomes et al. [24] |
| D. aseana | MFLUCC 12-0299aT | KT459414 | KT459432 | - | KT459448 | KT459464 | Hyde et al. [27] |
| D. australiana | BRIP 66145T | MN708222 | MN696530 | - | MN696522 | - | Wrona et al. [28] |
| D. averrhoae | SCHM 3605T | AY618930 | - | - | - | - | Chang et al. [29] |
| D. bounty | BRIP 59361aT | OM918690 | OM960617 | - | OM960599 | - | Tan and Shivas [26] |
| D. camelliae-oleiferae | HNZZ 027T | MZ509555 | MZ504718 | MZ504696 | MZ504707 | MZ504685 | Yang et al. [30] |
| D. ceratozamiae | CBS 131306T | JQ044420 | - | - | - | - | Crous et al. [31] |
| D. cercidis | CFCC 52565T | MH121500 | MH121582 | MH121460 | MH121542 | MH121424 | Yang et al. [32] |
| D. cf. heveae | CBS 681.84 | KC343117 | KC344085 | KC343601 | KC343843 | KC343359 | Gomes et al. [24] |
| D. chiangmaiensis | MFLUCC 18-0544T | OK393703 | - | - | OL439483 | - | de Silva et al. [33] |
| D. chrysalidocarpi | SAUCC 194.35T | MT822563 | MT855760 | MT855532 | MT855876 | MT855646 | Huang et al. [34] |
| D. delonicis | MFLU 16-1059T | MT215490 | MT212209 | - | - | - | Perera et al. [35] |
| D. drenthii | BRIP 66524T | MN708229 | MN696537 | - | MN696526 | - | Wrona et al. [28] |
| D. endocitricola | ZHKUCC 20-0012T | MT355682 | MT409290 | - | MT409336 | MT409312 | Dong et al. [36] |
| D. eres | CBS 138594T | KJ210529 | KJ420799 | KJ420850 | KJ210550 | KJ434999 | Udayanga et al. [37] |
| D. eucalyptorum | CBS 132525T | JX069862 | - | - | - | - | Crous et al. [38] |
| D. eugeniae | CBS 444.82 | KC343098 | KC344066 | KC343582 | KC343824 | KC343340 | Gomes et al. [24] |
| D. fraxini-angustifoliae | BRIP 54781T | JX862528 | KF170920 | - | JX852534 | - | Tan et al. [39] |
| D. fujianensis | JZB 320149T | MW010212 | MW056008 | - | - | MW205212 | Manawasinghe et al. [40] |
| D. fulvicolor | PSCG 051T | MK626859 | MK691236 | MK726163 | MK654806 | MK691132 | Guo et al. [23] |
| D. fusiformis | JZB 320156T | MW010218 | MW056014 | - | MW205234 | MW205218 | Manawasinghe et al. [40] |
| D. gossiae | BRIP 59730aT | OM918693 | OM960620 | - | OM960602 | - | Tan and Shivas [26] |
| D. guangxiensis | JZB 320094T | MK335772 | MK500168 | - | MK523566 | MK736727 | Manawasinghe et al. [41] |
| D. hongkongensis | CBS 115448T | KC343119 | KC344087 | KC343603 | KC343845 | KC343361 | Gomes et al. [24] |
| D. howardiae | BRIP 59697aT | OM918695 | OM960622 | - | OM960604 | - | Tan and Shivas [26] |
| D. huangshanensis | CNUCC 201903T | MN219730 | MN227011 | MN224558 | MN224678 | - | Zhou and Hou [25] |
| D. hunanensis | HNZZ 023T | MZ509550 | MZ504713 | MZ504691 | MZ504702 | MZ504680 | Yang et al. [30] |
| D. krabiensis | MFLUCC 17-2481T | MN047101 | MN431495 | - | MN433215 | Dayarathne et al. [42] | |
| D. limonicola | CBS 142549T | MF418422 | MF418582 | MF418342 | MF418501 | MF418256 | Guarnaccia and Crous [43] |
| D. liquidambari | SCHM 3621T | AY601919 | - | - | - | - | Chang et al. [29] |
| D. litchicola | BRIP 54900T | JX862533 | KF170925 | - | JX862539 | - | Tan et al. [39] |
| D. lithocarpi | CGMCC 3.15175T | KC153104 | KF576311 | - | KC153095 | - | Gao et al. [44] |
| D. meliae | CFCC 53089T | MK432657 | MK578057 | ON081662 | ON081654 | - | Cao et al. [45] |
| D. melitensis | CBS 142551T | MF418424 | MF418584 | MF418344 | MF418503 | MF418258 | Guarnaccia and Crous [43] |
| D. millettiae | GUCC 9167T | MK398674 | MK502089 | - | MK480609 | MK502086 | Long et al. [46] |
| D. musigena | CBS 129519T | KC343143 | KC344111 | KC343627 | KC343869 | KC343385 | Gomes et al. [24] |
| D. norfolkensis | BRIP 59718aT | OM918699 | OM960626 | - | OM960608 | - | Tan and Shivas [26] |
| D. oculi | HHUF 30565T | LC373515 | LC373519 | - | LC373517 | - | Ozawa et al. [47] |
| D. osmanthi | GUCC 9165T | MK398675 | MK502090 | - | MK480610 | MK502087 | Long et al. [46] |
| D. pandanicola | MFLUCC 17-0607T | MG646974 | MG646930 | - | - | - | Tibpromma et al. [48] |
| D. pascoei | BRIP 54847T | JX862532 | KF170924 | - | JX862538 | - | Tan et al. [39] |
| D. perseae | CBS 151.73 | KC343173 | KC344141 | KC343657 | KC343899 | KC343415 | Gomes et al. [24] |
| D. pescicola | MFLUCC 16-0105T | KU557555 | KU557579 | - | KU557623 | KU557603 | Dissanayake et al. [49] |
| D. phyllanthicola | SCHM 3680T | AY620819 | - | - | - | - | Chang et al. [29] |
| D. podocarpi-macrophylli | CGMCC 3.18281T | KX986774 | KX999207 | KX999246 | KX999167 | KX999278 | Gao et al. [50] |
| D. pseudomangiferae | CBS 101339T | KC343181 | KC344149 | KC343665 | KC343907 | KC343423 | Gomes et al. [24] |
| D. pseudooculi | HHUF 30617T | LC373515 | LC373519 | - | LC373517 | - | Ozawa et al. [47] |
| D. pseudophoenicicola | CBS 462.69T | KC343184 | KC344152 | KC343668 | KC343910 | KC343426 | Gomes et al. [24] |
| D. pterocarpicola | MFLUCC 10-0580a | JQ619887 | JX275441 | - | JX275403 | JX197433 | Udayanga et al. [51] |
| D. rhodomyrti | CFCC 53101T | MK432643 | MK578046 | MK442990 | MK578119 | MK442965 | Cao et al. [45] |
| D. salinicola | MFLU 18-0553T | MN047098 | - | - | MN077073 | - | Dayarathne et al. [42] |
| D. schimae | CFCC 53103T | MK432640 | MK578043 | MK442987 | MK578116 | MK442962 | Yang et al. [52] |
| D. searlei | BRIP 66528T | MN708231 | MN696540 | - | - | - | Wrona et al. [28] |
| D. sennae | CFCC 51636T | KY203724 | KY228891 | - | KY228885 | KY228875 | Yang et al. [53] |
| D. spinose | PSCG 383T | MK626849 | MK691234 | MK726156 | MK654811 | MK691129 | Guo et al. [23] |
| D. taoicola | MFLUCC 16-0117T | KU557567 | KU557591 | - | KU557635 | - | Dissanayake et al. [49] |
| D. tectonigena | MFLUCC 12-0767T | KU712429 | KU743976 | - | KU749371 | KU749358 | Doilom et al. [54] |
| D. viniferae | JZB 320071T | MK341551 | MK500112 | - | MK500107 | MK500119 | Manawasinghe et al. [41] |
| D. xishuangbanica | CGMCC 3.18282T | KX986783 | KX999216 | KX999255 | KX999175 | - | Gao et al. [50] |
| 1 | |||
|---|---|---|---|
| No. | δC, Type | δH (J in Hz) | |
| Fragment A | 1 | 169.3, C | - |
| 2 | 40.1, CH2 | 3.69, dd (17.7, 4.9) 3.94, dd (17.7, 7.3) | |
| 3-NH | - | 8.36, dd (6.9, 5.1) | |
| 4 | 168.6, C | - | |
| 5 | 41.6, CH2 | 3.77, d (4.9) | |
| 6-NH | - | 7.43, t (5.0) | |
| Fragment B | 7 | 170.4, C | - |
| 8 | 58.6, CH | 3.98, dd (7.8, 2.9) | |
| 9-NH | - | 7.85, d (7.8) | |
| 24 | 65.3, CH | 4.18, qd (6.3, 2.8) | |
| 24-OH | - | 5.02, br s | |
| 25 | 20.6, CH3 | 1.02, d (6.4) | |
| Fragment C | 10 | 175.3, C | - |
| 11 | 45.7, CH | 2.37, qd (7.5, 3.2) | |
| 12 | 70.6, CH | 3.83, m | |
| 13 | 41.2, CH2 | 1.65, t (6.9) | |
| 14 | 68.9, CH | 3.57, quin (6.1) | |
| 14-OH | - | 4.72, br s | |
| 15 | 36.4, CH2 | 1.38, m 1.48, m | |
| 16 | 27.5, CH2 | 1.88, m 1.99, m | |
| 17 | 130.7, CH | 5.45, m | |
| 18 | 131.6, CH | 5.09, dd (15.4, 8.9) | |
| 19 | 39.0, CH | 2.32, m | |
| 20 | 78.5, CH | 4.67, dd (9.6, 2.4) | |
| 21 | 34.8, CH2 | 1.62, td (7.0, 2.4) | |
| 22 | 26.4, CH2 | 1.15, m | |
| 23 | 11.7, CH3 | 0.84, t (7.1) | |
| 26 | 16.2, CH3 | 1.20, d (7.3) | |
| 27 | 17.2, CH3 | 0.88, d (6.9) | |
| 28 | 12.6, CH3 | 0.83, d (7.0) | |
| 2 | 3a | 3b | ||||
|---|---|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 171.9, C | - | 69.1, C | 4.97, d (12.3) 4.86, d (12.3) | 69.2, C | 4.97, d (12.3) 4.86, d (12.3) |
| 2 | 109.5, C * | - | 125.3, C | - | 125.3, C | - |
| 3 | 157.5, C * | - | 149.5, C | - | 149.6, C | - |
| 4 | 119.0, CH | 7.06, d (8.6) | 114.4, CH | 6.63, d (8.1) | 114.4, CH | 6.59, d (8.1) |
| 5 | 138.5, CH | 7.46, d (8.6) | 128.7, CH | 6.75, d (8.1) | 128.8, CH | 6.74, d (8.1) |
| 6 | 129.8, C | - | 125.5, C | - | 125.6, C | - |
| 7 | 143.5, C * | - | 142.6, C | - | 142.6, C | - |
| 8 | - | - | 105.8, C | - | 106.0, C | - |
| 9 | 131.0, C | - | 130.6, C | - | 131.1, C | - |
| 10 | 140.9, C | - | 139.3, C | - | 139.6, C | - |
| 11 | 143.3, C | - | 142.6, C | - | 142.5, C | - |
| 12 | 122.1, CH | 6.9, br s | 117.1, CH | 6.64, d (2.1) | 117.0, CH | 6.64, d (2.1) |
| 13 | 131.0, C | - | 127.7, C | - | 128.1, C | - |
| 14 | 122.9, CH | 6.9, br s | 120.2, CH | 7.24, d (2.1) | 120.1, CH | 7.22, d (2.1) |
| 15 | 20.6, CH3 | 2.24, s | 20.6, CH3 | 2.2, s | 20.7, CH3 | 2.2, s |
| 1′ | 29.7, CH2 | 3.13, br d (6.7) | 28.0, CH2 | 2.8, m (overlapped) | 28.0, CH2 | 2.8, m (overlapped) |
| 2′ | 121.4, CH | 5.06, br t (6.7) | 123.6, CH | 4.8, t (7.1) | 123.7, CH | 4.8, t (7.1) |
| 3′ | 133.9, C | - | 130.2, C | - | 130.2, C | - |
| 4′ | 17.6, CH3 | 1.50, s | 17.4, CH3 | 1.33, s | 17.4, CH3 | 1.34, s |
| 5′ | 25.6, CH3 | 1.64, s | 25.5, CH3 | 1.51, s | 25.6, CH3 | 1.52, s |
| 1″ | 64.2, CH2 | 3.93, t (9.7) 4.37, br d (9.5) | 63.7, CH2 | 4.27, dd (11.1, 2.1) 3.48, br dd (11.1, 9.0) | 64.3, CH2 | 4.27, dd (11.1, 2.0) 3.73, dd (11.1, 9.2) |
| 2″ | 79.6, CH | 3.99, m | 79.1, CH | 3.60, dd (9.0, 2.3) | 79.0, CH | 3.23, dd (9.2, 1.9) |
| 3″ | 72.4, C | - | 68.8, C | - | 69.1, C | - |
| 4″ | 23.5, CH3 | 1.20, s | 22.2, CH3 | 0.35, s | 23.0, CH3 | 0.52, s |
| 5″ | 25.6, CH3 | 1.30, s | 27.4, CH3 | 0.85, s | 27.2, CH3 | 0.84, s |
| No. | 4a | 4b | ||
|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 169.9, C | - | 169.9, C | - |
| 2 | 115.8, C | - | 115.9, C | - |
| 3 | 153.2, C | - | 153.3, C | - |
| 4 | 115.8, CH | 6.73, d (8.2) | 115.6, CH | 6.70, d (8.2) |
| 5 | 134.1, CH | 7.02, d (8.2) | 134.2, CH | 7.02, d (8.2) |
| 6 | 127.0, C | - | 127.0, C | - |
| 7 | 148.0, C | - | 148.0, C | - |
| 8 | 84.9, C | - | 85.1, C | - |
| 9 | 127.9, C | - | 127.9, C | - |
| 10 | 138.6, C | - | 138.8, C | - |
| 11 | 142.6, C | - | 142.7, C | - |
| 12 | 117.1, CH | 6.67, d (2.1) | 117.1, CH | 6.67, d (2.1) |
| 13 | 128.3, C | - | 128.5, C | - |
| 14 | 120.7, CH | 7.33, br s | 120.7, CH | 7.33, br s |
| 15 | 20.6, CH3 | 2.23, s | 20.6, CH3 | 2.23, s |
| 16-NH | - | 8.79, s | - | 9.04, br s |
| 1′ | 27.8, CH2 | 2.92, m (overlapped) | 27.8, CH2 | 2.92, m (overlapped) |
| 2′ | 122.9, CH | 4.79, br t (6.6) | 123.0, CH | 4.79, br t (6.6) |
| 3′ | 130.7, C | - | 130.8, C | - |
| 4′ | 17.3, CH3 | 1.35, s | 17.3, CH3 | 1.37, s |
| 5′ | 25.4, CH3 | 1.50, s | 25.5, CH3 | 1.52, s |
| 1″ | 63.8, CH2 | 3.46, dd (11.2, 9.2) 4.31, m | 64.1, CH2 | 3.68, dd (11.2, 9.2)4.31, m |
| 2″ | 79.4, CH | 3.56, dd (9.0, 1.8) | 79.4, CH | 3.29, m |
| 3″ | 68.8, C | - | 68.5, C | - |
| 4″ | 21.9, CH3 | 0.33, s | 22.7, CH3 | 0.60, s |
| 5″ | 27.7, CH3 | 0.92, s | 27.4, CH3 | 0.86, s |
| No. | 7a | 7b | ||
|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 144.8, CH | 7.78, d (1.7) | 144.8, CH | 7.83, d (1.7) |
| 2 | - | - | - | - |
| 3 | 157.6, C | - | 157.2, C | - |
| 4 | 105.0, CH | 6.82, s | 105.8, CH | 6.88, s |
| 4a | 142.6, C | - | 142.5, C | - |
| 5 | 107.4, C | - | 107.7, C | - |
| 6 | 185.5, C | - | 185.6, C | - |
| 7 | 81.9, C | - | 81.9, C | - |
| 8 | 43.3, CH | 3.54, dd (11.9, 1.7) | 43.1, CH | 3.54, dd (11.9, 1.7) |
| 8a | 116.2, C | - | 116.2, C | - |
| 9 | 117.6, CH | 6.51, d (15.9) | 120.2, CH | 6.61, d (15.7) |
| 10 | 140.8, CH | 7.07, d (15.9) | 132.2, CH | 7.41, d (15.7) |
| 11 | 132.2, C | - | 130.2, C | - |
| 12 | 146.5, CH | 5.73, d (9.8) | 144.0, CH | 5.52, d (9.9) |
| 13 | 34.3, CH | 2.46, m | 33.4, CH | 2.64, m |
| 14 | 29.6, CH2 | 1.39, m 1.26, m | 29.7, CH2 | 1.39, m 1.26, m |
| 15 | 11.9, CH3 | 0.82, t (7.4) | 11.9, CH3 | 0.82, t (7.4) |
| 16 | 20.2, CH3 | 0.96, d (6.6) | 21.0, CH3 | 0.96, d (6.6) |
| 17 | 12.3, CH3 | 1.81, d (0.9) | 19.9, CH3 | 1.88, d (1.1) |
| 18 | 18.7, CH3 | 1.20, s | 18.7, CH3 | 1.20, s |
| 19 | 174.6, C | - | 174.6, C | - |
| 20 | 47.0, CH | 3.41, br dd (11.9, 5.7) | 47.0, CH | 3.41, br dd (11.9, 5.7) |
| 21 | 65.7, CH | 4.04, m | 65.6, CH | 4.04, m |
| 21-OH | - | 5.22, br d (4.4) | - | 5.22, br d (4.4) |
| 22 | 22.7, CH3 | 1.35, d (6.3) | 22.8, CH3 | 1.35, d (6.3) |
| MIC (µg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Test Organisms | 1 | 2 | 3 (3a + 3b) | 4 (4a + 4b) | 5 | 6 | 7 (7a + 7b) | 8 | References |
| Acinetobacter baumanii | - | - | - | - | - | - | - | - | 0.26 c |
| Bacillus subtilis | - | - | 33.3 | 66.6 | 66.6 | 16.6 | 16.6 | 8.3 | 8.3 o |
| Candida albicans | - | - | - | - | - | - | - | - | 16.6 n |
| Chromobacterium violaceum | - | - | - | - | - | - | - | - | 0.83 o |
| Escherichia coli | - | - | - | - | - | - | - | - | 1.7 o |
| Mucor hiemalis | - | - | 66.6 | 33.3 | 66.6 | 16.6 | 8.3 | 66.6 | 8.3 n |
| Mycobacterium smegmatis | - | - | - | - | - | - | 16.6 | 8.3 | 1.7 k |
| Pseudomonas aeruginosa | - | - | - | - | - | - | - | - | 0.21 g |
| Rhodoturula glutinis | - | - | - | 33.3 | 66.6 | 16.6 | - | 66.6 | 4.2 n |
| Schizosaccharomyces pombe | - | - | - | 33.3 | 33.3 | 66.6 | - | 33.3 | 8.3 n |
| Staphylococcus aureus | - | - | 33.3 | 66.6 | 66.6 | 16.6 | 66.6 | 8.3 | 0.83 o |
| Wickerhamomyces anomalus | - | - | - | - | - | 66.6 | - | - | 8.3 n |
| IC50 (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Cell Lines | 1 | 2 | 3 (3a + 3b) | 4 (4a + 4b) | 5 | 6 | 7 (7a + 7b) | 8 | Epothilone B |
| KB3.1 | - | SI | 32.9 | 31.9 | 22.0 | 20.0 | 5.3 | 2.3 | 4.5 × 10−5 |
| L929 | - | - | 51.6 | 59.2 | 54.4 | 49.5 | 12.9 | 2.8 | 4.3 × 10−4 |
| A431 | n.t | n.t | n.t | n.t | n.t | 19.3 | 4.1 | 2.4 | 5.5 × 10−5 |
| MCF-7 | n.t | n.t | n.t | n.t | n.t | 17.7 | 0.9 | 2.0 | 6.1 × 10−5 |
| A549 | n.t | n.t | n.t | n.t | n.t | 20.8 | 2.3 | 2.6 | 6.9 × 10−5 |
| SKOV-3 | n.t | n.t | n.t | n.t | n.t | 42.5 | 3.3 | 3.3 | 9.9 × 10−5 |
| PC-3 | n.t | n.t | n.t | n.t | n.t | 17.9 | 1.1 | 1.9 | 7.3 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matio Kemkuignou, B.; Lambert, C.; Stadler, M.; Kouam Fogue, S.; Marin-Felix, Y. Unprecedented Antimicrobial and Cytotoxic Polyketides from Cultures of Diaporthe africana sp. nov. J. Fungi 2023, 9, 781. https://doi.org/10.3390/jof9070781
Matio Kemkuignou B, Lambert C, Stadler M, Kouam Fogue S, Marin-Felix Y. Unprecedented Antimicrobial and Cytotoxic Polyketides from Cultures of Diaporthe africana sp. nov. Journal of Fungi. 2023; 9(7):781. https://doi.org/10.3390/jof9070781
Chicago/Turabian StyleMatio Kemkuignou, Blondelle, Christopher Lambert, Marc Stadler, Simeon Kouam Fogue, and Yasmina Marin-Felix. 2023. "Unprecedented Antimicrobial and Cytotoxic Polyketides from Cultures of Diaporthe africana sp. nov." Journal of Fungi 9, no. 7: 781. https://doi.org/10.3390/jof9070781