
Rust Fungi on Medicinal Plants in Guizhou Province with Descriptions of Three New Species
by
Qianzhen Wu
1,2,
Minghui He
1,
Tiezhi Liu
3,
Hongmin Hu
1,2,
Lili Liu
1,4,5,
Peng Zhao
6,* and
Qirui Li
1,2,* 1
State Key Laboratory of Functions and Applications of Medicinal Plants, Guizhou Medical University, Guiyang 550004, China
2
The High Efficacy Application of Natural Medicinal Resources Engineering Center of Guizhou Province (The Key Laboratory of Optimal Utilization of Natural Medicine Resources), School of Pharmaceutical Sciences, Guizhou Medical University, University Town, Guian New District, Guizhou 550004, China
3
College of Chemistry and Life Sciences, Chifeng University, Chifeng 024000, China
4
Immune Cells and Antibody Engineering Research Center of Guizhou Province, Guizhou Medical University, Guiyang 550004, China
5
Key Laboratory of Biology and Medical Engineering, Guizhou Medical University, Guiyang 550004, China
6
State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences (CAS), Beijing 100101, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2023, 9(9), 953; https://doi.org/10.3390/jof9090953
Submission received: 12 July 2023
/
Revised: 7 September 2023
/
Accepted: 7 September 2023
/
Published: 21 September 2023
/
Corrected: 1 November 2023
(This article belongs to the Special Issue Rust Fungi)
Abstract
:During the research on rust fungi in medicinal plants of Guizhou Province, China, a total of 9 rust fungal species were introduced, including 3 new species (Hamaspora rubi-alceifolii, Nyssopsora altissima, and Phragmidium cymosum), as well as 6 known species (Melampsora laricis-populina, Melampsoridium carpini, Neophysopella ampelopsidis, Nyssopsora koelrezidis, P. rosae-roxburghii, P. tormentillae). Notably, N. ampelopsidis and P. tormentillae were discovered for the first time in China, while M. laricis-populina, Me. carpini, and Ny. koelreuteriae were first documented in Guizhou Province. Morphological observation and molecular phylogenetic analyses of these species with similar taxa were compared to confirm their taxonomic identities, and taxonomic descriptions, illustrations and host species of those rust fungi on medicinal plant are provided.
1. Introduction
Rust fungi comprise the largest and most ubiquitous group of obligately biotrophic fungi on vascular plants [1]. The economic impact of rust fungi cannot be ignored. Because of rust fungi, many economic plants suffer diseases and reduce yields [2]. For example, when wheat is harmed by Puccinia striiformis Westend., its yield can be reduced up to 50% [3]. The impact of rust fungi on tropical crops is also immeasurable [4]. The coffee rust fungus has a global distribution and is often found in the coffee growing areas of China, with a greater impact on the main coffee producing countries [2,5,6].
More than 8000 species of rust fungi have been identified, mainly on the basis of their morphological characteristics of teliospores and spermonogia, however, there are still a larger number of genera incertae sedis [7,8,9,10,11]. Rust occurs on ferns to advanced monocots and dicots, and they are obligate biotrophic phytopathogens that produce not only basidiospore but also four other different types of spores: aeciospore, urediniospore, teliospore, and spermatia [8,12,13]. Many species in the order Pucciniales were not described with all types of spores, and have various lifestyles (micro-, hemi-, demi-, or macrocyclic), with alternation on single (autoecious) or two unrelated host plants (heteroecious) [7,14]. To date, approximately 8400 rust species are currently recognized worldwide, and 71 genera and 1175 species have been discovered in China so far [11]. However, species diversity, host alternation and geographic distribution of rust fungi in China remain poorly understood.
Medicinal plants are also infected by rust fungi on a large scale [15]. By the end of 2020, a total of 79 rust species have been reported on 76 medicinal plant species from 33 families, and these rust fungi restricts the development and utilization of medicinal plants and affects the quality of botanicals [16,17]. In recent years, as many as 3924 species of medicinal plants are cultivated in Guizhou province, and the number of medicinal plants is increasing number year by year [18]. As important pathogenic fungi, rust can infect the leaf and stem of a variety of medicinal plants and affect their quality and yield which eventually hampered the development and utilization of medicinal plant resources [19]. For example, rust diseases are frequently found on pepper leaves in Guizhou province, and those diseases seriously affect the normal development of pepper, with their incidence reaching 90% in serious infections [20]. However, there are few researches on rust species infecting medicinal plant diseases in Guizhou province [21]. Therefore, it is of great significance to investigate the species diversity of rust fungi on important medicinal plants for local medicinal production.
In 2021, an investigation of rust fungi on the medicinal plants was carried out in Guizhou province, China. Nine species including three new species were found on medicinal plants. Detailed descriptions and illustrations of all those novel species and other species on the medicinal plants are provided.
2. Materials and Methods
2.1. Sample Collections
Rust infected specimens were collected from Guiyang, Qingzhen and Anshun cities in Guizhou province, China. All hosts and habitats information of specimens was recorded. For each specimen, part of specimens was kept in a refrigerator at 4 °C, and the other part was made as a dry specimen [22]. Specimens were deposited in both Mycological Herbarium of the Chifeng University, Inner Mongolia, China (CFSZ) and Herbarium of Guizhou Medical University (GMB).
2.2. Morphology
The specimens were observed under a stereomicroscope (Nikon SMZ745T, Nikon Corporation, Tokyo, Japan) and shot with a Canon digital camera (Canon EOS 1500D, Canon Inc., Tokyo, Japan) fitted on. Microscope images of the samples were taken by a Canon EOS 700D digital camera fitted on the Nikon ECLIPSE Ni compound microscope (Nikon, Japan). Measurements were taken with the Tarosoft (R) Image Frame Work (v.0.9.7). More than 30 morphological characteristics such as teliospores, urediniospores, and paraphyses were measured for each specimen. Photo plates were arranged by using Adobe Photoshop CS6 v. 13 (Adobe Systems Software Ireland Ltd, San Jose, USA). The different spore stages of rust fungi are designated by the following Roman numerals: spermogonia/spermatia (0), aecia/aeciospores (I), uredinia/urediniospores (II), telia/teliospores (III), and basidia/basidiospore (IV). We applied the definitions of spore stage based on Cummins and Hiratsuka [7], and followed morphological types of spermogonia designated by Hiratsuka and Hiratsuka [23].
2.3. DNA Extraction, Polymerase Chain Reaction (PCR), and Sequencing
The rust sori were picked out into a sterilized centrifuge tube with a sterilized fine needle for DNA extraction. Genomic DNA was extracted following the manufacturer’s protocol of the OMEGA E.Z.N.A.® Fungal Genomic DNA Extraction Kit (D3390, Guangzhou Feiyang Bioengineering Co., Ltd., Guangzhou, China). DNA extracts were stored at –20 °C. PCR was carried out in a volume of 25 μL containing 9.5 μL of ddH2O, 12.5 μL of 2 × Taq PCR Master Mix (2 × Taq Master Mix with dye, TIANGEN, China), 1 μL of DNA extraction and 1 μL of forward and reverse primers (10 µm each) in each reaction. Primers pairs, ITS4/ITS5 and LR0R/LR5 (Sangon Biotech, Shanghai, China) were used to amplify the regions of internal transcribed spacer (ITS) and large subunit ribosomal (LSU), respectively [8,24,25,26]. PCR profiles for the ITS and LSU were: initially at 95 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing at 52 °C for 1 min, polymerization at 72 °C for 1.5 min and a final extension at 72 °C for 10 min. PCR products were sequenced by Sangon Biotech (Shanghai) Co., Ltd., China.
2.4. Phylogenetic Studies
All sequences used for phylogenetic tree construction were listed in Table 1. Sequences were aligned by MAFFT v. 7.394 (https://www.ebi.ac.uk/Tools/msa/mafft/) (accessed on 12 September 2023) [27] and adjusted to ensure maximum similarity using TrimAl v1.4.1 [28]. Alignments were converted from FASTA to PHYLIP format by using Alignment Transformation Environment online (https://sing.ei.uvigo.es/ALTER/) (accessed on 12 September 2023) [29]. Maximum Likelihood (ML) analyses and Bayesian posterior probabilities (BYPP) based on a combination of ITS and LSU sequence data were performed using RAxML-HPC 7.4.2 BlackBox [30] and MrBayes v. 3.2.7 tools in the CIPRES Science Gateway platform [31,32]. GTR+I+G was estimated as the best-fit substitution model by jModelTest2 on XSEDE v.2.1.6 [33,34]. The Bootstrap values of ML analyses were obtained by running 1000 replicates by using a Markov chain Monte Carlo (MCMC) method to approximate the posterior probabilities of trees. Six simultaneous Markov Chains were run for 3,000,000 generations and trees were sampled every 1000th generation. Finally, the trees were visualized in FigTree v.1.4.4 [35] and edited by using Adobe Photoshop CS6 v. 13 software. The final alignment and phylogenetic trees were deposited in TreeBASE v. 2 under the submission ID30041 (http://www.treebase.org/) (accessed on 12 September 2023).
3. Results
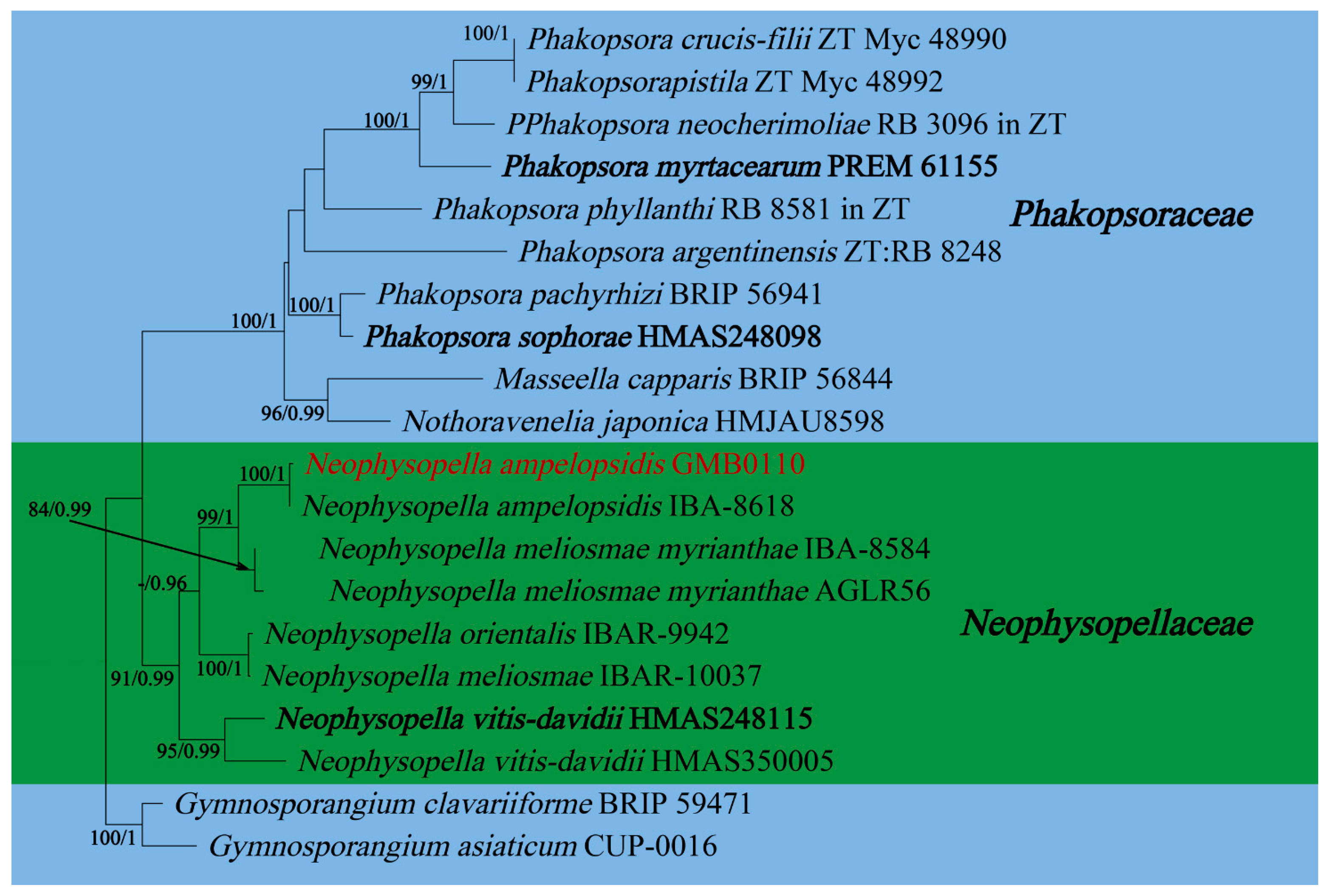
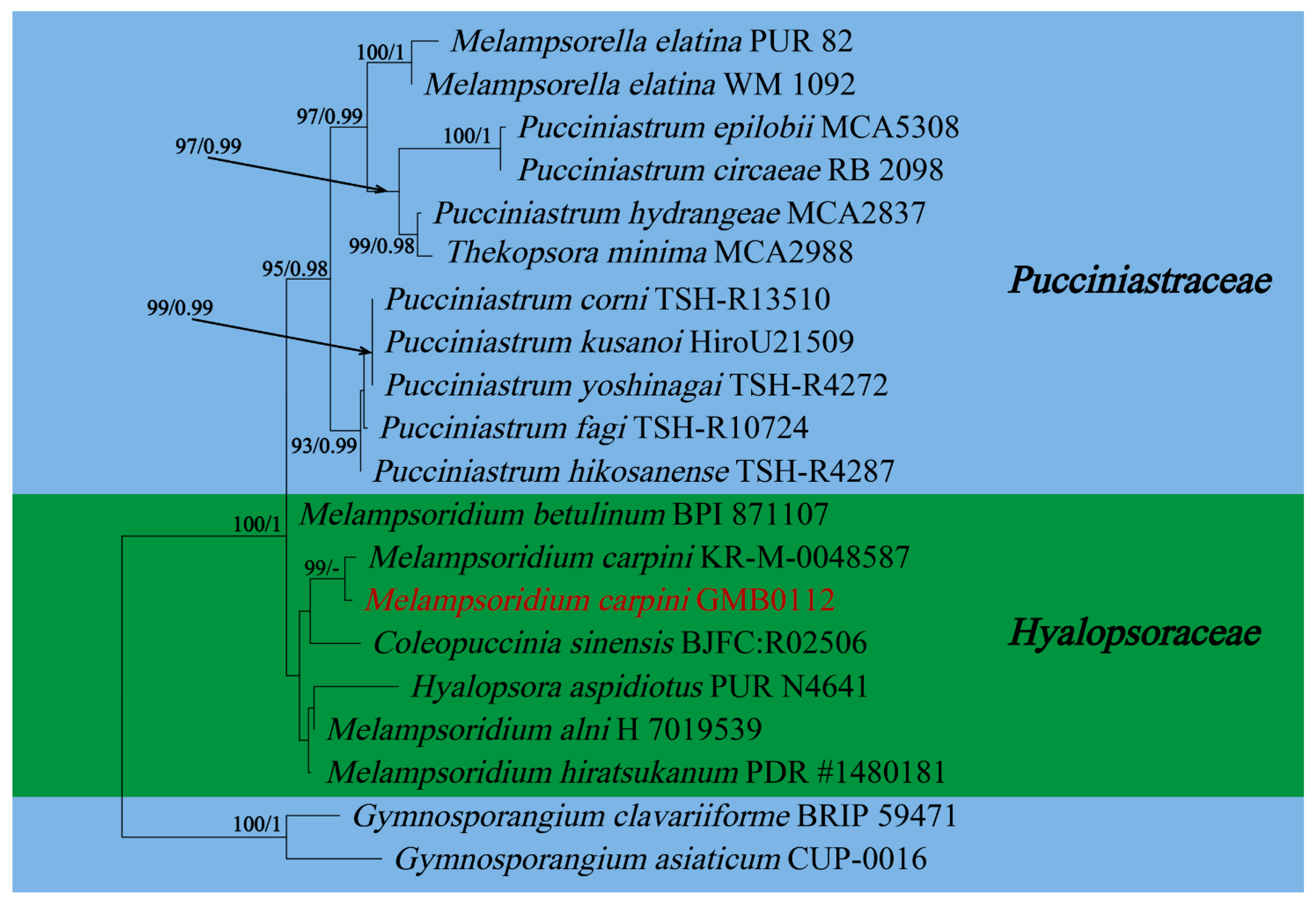
In this study, 11 samples were collected from 9 species of medicinal plants in nine genera in seven families in Guizhu Province. Ten species including three new species were identified based on morphological and molecular phylogenetic studies. Morphological and phylogenetically allied taxa were selected for the final phylogenetic analyses, mainly following Zhao et al. [11]. The alignment for Melampsoraceae includes 1200 character (ITS: 532 bp, LSU: 1200 bp) (Figure 1). The dataset of phylogenetic tree from Neophysopellaceae and Phakopsoraceae has 1162 characters (ITS: 474 bp, LSU: 1162 bp) including gaps (Figure 2). The alignment for Neophysopellaceae and Phakopsoraceae phylogenetic tree contained 20 taxa. The taxa from genera Gerwasia, Hamaspora, Kuehneola, Phragmidium, Trachyspora, and Xenodochus were included in the phylogenetic tree of the family Phragmidiaceae (Figure 3). The alignment is made up of 71 species and has 1244 characters including gaps (ITS: 392 bp, LSU: 1244 bp). The phylogenetic tree of the families Pucciniastraceae and Hyalopsoraceae includes the taxa from genera Coleopuccinia, Hyalopsora, Melampsorella, Melampsoridium, and Pucciniastrum (Figure 4). In the phylogenetic tree of Pucciniastraceae, there are 20 taxa with 1244 characters, including gaps (ITS: 351 bp, LSU: 1244 bp). Twenty-seven representative species from the Gymnosporangiaceae, Sphaerophragmiaceae, and Uredinineae incertae sedis were chosen for Figure 5, which included 1272 characters including gaps (ITS: 398, LSU: 1272). Based on morphological and multi-locus phylogenetic characterisation, 13 specimens were identified to 9 species in 6 genera 5 families (Hamaspora rubi-alceifolii sp. nov., Nyssopsora altissima sp. nov., Phragmidium cymosum sp. nov., Melampsora laricis-populina, Melampsoridium carpini, Neophysopella ampelopsidis, Ny. koelreuteriae, P. rosae-roxburghii and P. tormentillae). Among them, three novel species were recognized, moreover, 2 species were reported first in China, and 4 species were first reported on medicinal plant.
Taxonomy
Based on morphology and molecular phylogeny, all collected rust specimens on medicinal plants were identified as three new species (Hamaspora rubi-alceifolii sp. nov., Nyssopsora altissima sp. nov., Phragmidium cymosum sp. nov.) and six known species (Melampsora laricis-populina, Melampsoridium carpini, Neophysopella ampelopsidis, Ny. koelreuteriae, P. rosae-roxburghii and P. tormentillae) All species parasitic on medicinal plants are described and illustrated below.
Hamaspora rubi-alceifolii Q. Z. Wu, T. Z. Liu, P. Zhao & Q. R. Li, sp. nov. Figure 6
MycoBank number: MB847104
Etymology: Epithet follows the epithet of host species, Rubus alceifolius Poir.
Holotype: GMB0109
Parasitic on the leave and stem of R. alceifolius. Telia up to 5 mm long, mostly hypophyllous, occasionally amphigenous and stem, caespitose, filiform, fluffy, golden yellow when fresh, white when dry; Teliospores 116–230 × 20–10 µm (av. = 184 × 17 µm, n = 30), fusiform, hyaline 3–6 septate, mostly 5–6 septate, smooth, the contents yellow when fresh, solid apex 9–30 µm (av. = 23 µm, n = 30). Spermogonia, aecia and uredinia were not observed.
Materials examined: CHINA, Guizhou Province, Guiyang City, Campus of Guizhou Medical University (26°22′48.37″ N, 106°37′30.33″ E), III on leaf of R. alceifolius Poir., 6 October 2021, Q. Z. Wu and L. L. Liu, GMB0109, holotype, CFSZ 50531, isotype; CHINA, Guizhou Province, Guiyang City, Huaxi District, III on leaf of R. alceifolius Poir., 20 October 2021, Q. Z. Wu and L. L. Liu, GMB0116.
Notes: Hamaspora rubi-alceifolii is characterized by 5–6 septate teliospore with long solid apex up to 30 µm, and hypophyllous telia (Figure 6). Phylogenetically, it formed a distinct clade sister to H. acutissima with high support values (100% ML, 1 BYPP; Figure 3). Morphologically, the differences between H. rubi-alceifolii and H. acutissima are in the number of septa in the teliospores (5–6 vs. 2–3), smaller teliospores (116–230 × 10–20 µm vs. 158–205 × 18–25 µm), and smaller solid apex (9–30 µm vs. 20–40 µm). Both H. rubi-sieboldii and H. rubi-alceifolii exhibit similar teliospore morphology with consistent solid apex size. However, the difference between H. rubi-alceifolii and H. rubi-sieboldii is that the former has more cells (5–6 vs. 4) and smaller solid apex (116–230 × 10–20 µm vs. 118–240 × 15–23 µm) [73,74,75,76].
Roots and leaves of Rubus alceifolius is a traditional Chinese medicine, which can be used for treatment of acute and chronic hepatitis, hepatosplenomegaly and other liver damage diseases [77,78,79,80,81]. Previously, thirteen Hamaspora species have already been reported on Rubus species [76]. For the convenience of recognition, a worldwide identification key for the Hamaspora has been provided.
![Jof 09 00953 i001]()

Melampsora laricis-populina Kleb., Z. PflKrankh. 12: 43 (1902) Figure 7
Mycobank number: MB18930
Uredinia, mostly hypophyllous, seldom epiphyllous, in little groups, 0.25 mm, Bright yellow. Urediniospores oblong to broadly ellipsoid or obovoid, hyaline to light yellow with yellowish granules, 33–45 × 21–26 µm (av. = 37 × 24 µm, n = 30), wall 1.8–3 µm thick, echinulate except at the smooth apex; Paraphyses clavate to capitate, hyaline to pale yellow, 50–71 × 12–20 µm (av. = 62 × 17 µm, n = 30), wall thick, up to 9 µm at the apex.
Materials examined: CHINA, Guizhou Province, Kaili City, Xiasi Town (26°26′29.48″ N, 107°47′30.33″ E) II on Populus lasiocarpa Oliv., 1 October 2021, Q. Z. Wu, GMB0097.
Notes: Melampsora laricis-populina is characterized by the echinulate urediniospores that are obovate or oval, with golden yellow cytoplasm; roughly spherical paraphyses with swollen tips. The uredinial morphologies of our specimen (GMB0097) are consistent with those of M. laricis-populina [82,83]. Phylogenetic tree (Figure 1) showed that our specimen (GMB0097) was clustered with M. laricis-populina with the high bootstrap supports (83/-). Thus, here we confirmed the rust fungus on Populus lasiocarpa as M. laricis-populina. Populus lasiocarpa has hemostatic function and mainly used for treatment for bleeding from trauma [84,85]. Previously, M. laricis-populina has already been reported on Populus lasiocarpa in Japan [86] and Norway [87], and it has been reported in northwest region of China [88].
Melampsoridium carpini (Nees) Dietel, in Engler & Prantl, Nat. Pflanzenfam., Teil. I (Leipzig) 1(1**): 551 (1900) Figure 8
Basionym: Caeoma carpini Nees, Syst. Pilze (Würzburg): 16 (1816) [1816-17]
Mycobank number: 205589
Uredinia, hypophyllous, scattered or grouped on yellow spots, 0.2 mm diam., yellow. Urediniospores long obovoid, clavate or pear shaped, 19–31 × 13–18 µm (av. = 26 × 15 µm, n = 30), yellow; spore wall hyaline, echinulate, on apex smooth, 0.8–1.5 µm thick.
Materials examined: CHINA, Guizhou Province, Qingzhen City, Xiasi Town (26°27′18.48″ N, 107°20′7.33″ E) II on Carpinus turczaninowii Hance, 8 October 2021, Q. Z. Wu and L. L. Liu, GMB0112, CFSZ 50543.
Notes: Carpinus turczaninowii is commonly used to treat bruises, canker sores and swellings, as recorded in the Pharmacopoeia of the People’s Republic of China. Carpinus turczaninowii can alleviate arterial damage and inflammation caused by hyperglycemia [89]. It contains Pheophorbide A, which has anti-cancer and anti-inflammatory activity [90,91]. We collected rust infected Carpinus turczaninowii in Guizhou province in China, and the urediospores of our specimen (GMB0112) are consistent with those of Me. carpini, which is characterized by sparse thorns on the surface and a smooth top of urediospores [92,93,94]. According to the phylogenetic tree (Figure 4.), the new collection (GMB0112) was clustered with Me. carpini with the high bootstrap values (99/-). This species has been found in Anhui, Chongqing, Sichuan, Taiwan and other provinces in China [94], which is the first record of Me. carpini from Guizhou province, China.
Neophysopella ampelopsidis (Dietel & P. Syd.) Jing X. Ji & Kakish., in Ji, Li, Li & Kakishima, Mycol. Progr. 18(6): 863 (2019) Figure 9
Basionym: Phakopsora ampelopsidis Dietel & P. Syd. [as ’ampelosidis’], in Dietel, Hedwigia 37: 217 (1898)
Mycobank number: MB830298
Uredinia hypophyllous, grouped on brown or yellow spots. the paraphyses, incurved, 30–43 × 6–14 µm (av. = 37 × 11 µm, n = 30), wall 2–3 µm thick (av. = 2.5 µm, n = 30). Urediniospores, obovoid or obovoid-ellipsoid, dark yellow or brown, 20–32 × 15–21 µm (av. = 25 × 18 µm, n = 30), walls hyaline, echinulate. The wall was colorless or pale yellowish, equally ca 0.8–1.5 µm thick, and evenly echinulate.
Materials examined: CHINA, Guizhou, Qingzhen City, Xiasi Town (26°27′18.18″ N, 107°20′7.13″ E) II on Ampelopsis sinica (Mig.) W.T. Wang., 22 July 2021. Q. Z. Wu and L. L. Liu, GMB0110, CFSZ 50532.
Notes: Ampelopsis sinica root (ASR) is a traditional Chinese medicine known to have a hepatoprotective function. Moreover, it has been proven having anti-hepatocellular carcinogenic activity and to inhibit Hepatitis B virus activity [95,96,97]. Our rust collection (GMB00110) on Ampelopsis sinica is compatible with Neophysopella ampelopsidis [98]. Phylogenetically, our collection clustered with N. ampelopsidis (IBA-8618) with the high bootstrap supports (100/1, in Figure 2). N. ampelopsidis has been previously introduced from Japan, Philippines and Taiwan provinces of China [98]. This is the first record of N. ampelopsidis from the Chinese mainland.
Nyssopsora altissima Q. Z. Wu, T. Z. Liu, P, Zhao & Q. R. Li, sp. nov. Figure 10
MycoBank number: MB847103
Etymology: Epithet follows the epithet of host species, Ailanthus altissima (Mill.) Swingle.
Holotype: GMB0103
Parasitic on the leave of A. altissima. Uredinia, usually amphigenous, sparse or aggregated, pulverulent, golden; Urediniospores subglobose or ellipsoid, 18–23 × 16–24 µm (av. = 21 × 20 µm, n =30), cell wall 1.2–2.4 µm (av. = 1.5 µm, n = 30) thick, echinulate. Telia soft hypophyllous, aggregated, rounded, pulverulent, dark; Teliospores subglobose or globose-trigonal, septa constricted slightly, 2–3 cells are mostly 3, reddish-brown to opaque, 28–38 × 20–39.5 µm (av. = 33.5 × 33 µm, n = 30), projections up to 14 (av. = 12, n = 30), 1–6 (av. = 2, n = 30) apical furcations, 2.8–7.8 µm (av. = 4.5 µm n = 30) long and 1.3–3.3 µm (av. = 2 µm, n = 30) thick, each cell has 1–2 germination pores; pedicel hyaline, persistent, about 33–57 µm (av. = 44 µm, n = 30) long, 5.5–9 µm (av. = 7.8 µm, n = 30) thick.
Materials examined: CHINA, Guizhou Province, Guiyang City, Campus of Guizhou Medical University (26°22′48.37″ N, 106°37′30.33″ E), II, III on leave of A. altissima, 8 July 2021, Q.Z. Wu and L.L. Liu, GMB0103, holotype, CFSZ 50535, isotype.
Notes: Phylogenetically, Nyssopsora altissima was phylogenetically allied to Ny. echinata with a high bootstrap support (86/0.99, Figure 5). Morphologically, Ny. altissima differs from Ny. echinata by the number of germination pores in teliospores (3 vs. 2), the bigger teliospores (28–38 × 20–39.5 µm vs. 25–30 × 23–27 µm), and the shorter processes (2.8–7.8 µm vs. 6.6–12 µm). Morphologically, Ny. altissima can be distinguished from Ny. cedrelae by several morphological differences that bigger urediniospores (18–23 × 16–24 µm compared to 15–19 × 13–17 µm), smaller teliospores (28–38 × 20–39.5 µm vs. 28–45 × 18–43 µm), and fewer projections (1–14 vs. 15–22). Additionally, Ny. altissima has shorter hyaline (33–57 µm vs. 40–65 µm) and finer hyaline (5.5–9 µm vs. 10–12 µm) than those of Ny. cedrelae [76,99].
Ailanthus altissima has been used as a medicinal herb to hemostasis and Anti-diarrhea documented in Chinese pharmacopoeia [100]. Ailanthus altissima has the anti-malarial, anti-viral and anti-tumor active ingredient and shows potential as a novel drug for the treatment of prostate cancer [101,102,103]. To date, another rust species, Ny. cedrelae, has already been reported on Ailanthus altissima [76], here we reported another Nyssopsora species on Ailanthus altissima. Compared to Ny. cedrelae, Ny. altissima exhibits larger-sized urediniospores (18–23 × 16–24 µm vs. 15–19 × 13–17 µm), shorter apical furcations (2.8–7.8 µm vs. 2–10 µm), and a shorter pedicel (33–57 µm vs. 120 µm).
Nyssopsora koelreuteriae (Syd. & P. Syd.) Tranzschel, J. Soc. bot. Russie 8: 132 (1925) [1923] Figure 11
Basionym: Triphragmium koelreuteriae Syd. & P. Syd. 1913
Mycobank number: MB335240
Parasitic on leaves of Koelreuteria bipinnata, surrounded by yellowish margins spermagonia, aecia and uredinia unknown. Telia amphigenous, scattered or slightly clustered, densely aggregated in groups, confluent, naked, erumpent, blackish, chestnut-brown. Teliospores 27–31 × 24–32 µm (av. = 28 × 27 µm, n = 30), 3-celled with a single proximal cell and two collateral distal cells, triquetrous pyriform, strongly constricted at septa, blackish–brown; walls uniformly 1.5–3 µm thick, pale yellow when young and becoming blackish–brown when older. Projections up to 18 and branched at the tips, 2–3 branched at apex. Pedicel 20–26 × 4.5–6.5 µm (av. = 23 × 6 µm, n = 30), persistent, hyaline.
Materials examined: CHINA, Guizhou Province, Anshun City, Longdong Scenic Area (26°6′32.28″ N, 105°52′30.23″ E) III on Koelreuteria bipinnata Franch., July 10, 2021 Q.Z. Wu, GMB0105.
Notes: The leaves of Koelreuteria bipinnata have strong antimicrobial activity, and their extracts contain a variety of components that can inhibit bacteria and fungi [104,105]. We collected rust samples on the leaves of Koelreuteria bipinnata, and it can be identified as Ny. koelreuferiae based on both morphological and molecular evidences (Figure 5 and Figure 11). This rust fungus has already been reported in Zhejiang province, China in 1928 [106], and here we reported it on the same host species in Guizhou province.
Phragmidium cymosum Q. Z. Wu, T. Z. Liu, P, Zhao & Q. R. Li, sp. nov. Figure 12
MycoBank number: MB847102
Etymology: Epithet follows to the epithet of host species, Rosa cymosa Tratt.
Holotype: GMB0108
Parasitic on the leaves of Rosa cymosa. Uredinia Uredo-type (Uraecium type II), hypophyllous, yellowish, aggregated, 0.1–0.4 mm (av. = 0.2 mm, n = 30), with thick-walled, gradual thickening by the roots of the paraphyses, incurved, intermixed paraphyses, 25–70 × 9–15 µm (av. = 44 × 11 µm, n = 30), wall 3–11 µm thick (av. = 7 µm, n = 30), top width 5–13 µm thick (av. = 8.5 µm, n = 30), Urediniospores borne singly, mostly echinulate, globose, broadly or obovoid, echinulate, 19–27 × 21–29 µm (av. = 24 × 26 µm, n = 30), cytoplasm orange, wall 1–2.5 µm thick (av. = 1.5 µm, n = 30).
Materials examined: CHINA, Guizhou Province, Guiyang City, Campus of Guizhou Medical University (26°22′46.37″ N, 106°37′29.33″ E), II on R. cymosa, 6 October 2021, Q. Z. Wu, GY-XGQW GMB0108, holotype, CFSZ 50542, isotype; CHINA, Guizhou Province, Guiyang City, Campus of Guizhou Medical University (26°22′48.22″ N, 106°37′30.12″ E), II on R. cymosa, 6 October 2021, Q. Z. Wu, GMB0115.
Notes: Phragmidium cymosum formed a distinct lineage in the phylogenetic tree (Figure 3) with a high support rate (98/1). Morphologically, it differs from those species by its paraphyses with relatively thick wall, by the width of the urediniospores (21–29 µm in P. cymosum; 18–21 µm in P. japonicum; 11–18 µm in P. jiangxiense; 15–20 in P. rosae-multiflorae) [63,76]. In addition, the paraphyses of P. cymosum (25–70 × 9–15 µm) are thicker compared to those of P. jiangxiense (22–41 × 6–10 µm), and the cell walls of the paraphyses in P. cymosum (3–11 µm) are thicker than those of P. rosae-multiflorae (1 µm). Phragmidium rosae-multiflorae was once reported on the same host species in China [107].
The leaves, flowers and roots of Rosa cymosa can be used as the Chinese herbal medicine. Moreover, it has anti-inflammatory components, can be used to treat burns, analgesic [108,109,110].
Phragmidium rosae-roxburghii J.E. Sun & Yong Wang bis Figure 13
MycoBank number: MB845041
Parasitic on the leave of Rosa roxburghii Tratt. Uredinia Uredo-type (Uraecium type II), hypophyllous, yellowish, aggregated, with thick-walled, gradual thickening by the roots of the paraphyses, incurved, intermixed paraphyses, 23–50 × 8–13.5 µm (av. = 36.5 × 10 µm, n = 30), wall 2–8.5 µm thick (av. = 5.5 µm, n = 30), Urediniospores borne singly, mostly echinulate, globose, broadly or obovoid, echinulate, 18–29 × 17–24 µm (av. = 24 × 20 µm, n = 30), cytoplasm orange, wall 0.5–1.5 µm thick (av. = 1 µm, n = 30).
Materials examined: CHINA, Guizhou province, Anshun City, Longdong Scenic Area (26°6′46.32″ N, 105°52′32.23″ E) II on Rosa roxburghii Tratt, 10 July 2021 Q.Z. Wu, GMB0104, CFSZ 50540.
Notes: Rosa roxburghii as a Chinese herbal medicine is used as a remedy for respiratory diseases. It has been recently reported to be an antioxidant and anticoagulant, and also be used to treat dyspepsia, dysentery, hypo immunity, and neurasthenia [65,111]. Our collection (GMB0104) and P. rosae-roxburghii form a clade in phylogenetic tree with a high support value (100/1). The morphological characteristics of GMB0104 are consistent with those of P. rosae-roxburghii. Phragmidium rosae-roxburghii was introduced based on the specimen collected from Guizhou province [65]. Here we reported it on medicinal plant in the same province.
Phragmidium tormentillae Fuckel, Jb. nassau. Ver. Naturk. 23-24: 46 (1870) [1869-70] Figure 14
Mycobank number: MB177066
Spermogonia were not observed. Uredinia hypophyllous, scattered, subepidermal and erumpent becoming pulverulent, 0.5 mm diam. Urediniospores circular or obovoid, pale yellow, 18–24 × 14–20 µm (av. = 211 × 17 µm, n = 30), walls hyaline, echinulate, 1–1.5 mm thick.
Materials examined: CHINA, Guizhou Province, Qingzhen City, Xiasi Town (26°27′16.48″ N, 107°20′8.23″ E) II on Potentilla simulatrix Th. Wolf, 22 July 2021 Q.Z. Wu, GMB0114.
Notes: Potentilla is documented in most areas of China, which was used as a traditional Chinese medicine for hemostasis and treatment of malaria [112]. The pharmacological activities of Potentilla are mainly related to antioxidant, hypoglycemic, anti-inflammatory, antibacterial, antitumor, and cardiovascular system protective effects [113]. It also has positive effects on hemorrhagic cystitis [114]. Our collection on Potentilla simulatrix (GMB0110) is located in the same clade with P. tormentillae (Figure 3). Based on the size, shape and wall thickness of the urediniospores, as well as the presence of sparse spines on the surface of the urediniospores, new collection is identified as P. tormentillae [60,115,116]. However, no spermatozoa and paraphyses were observed in new collections. Phragmidium tormentillae was firstly collected in Norway in 1895 [116]. This is the first record for Chinese mainland.
4. Discussion
The research on rust diseases in medicinal plants has been relatively less compared to those in economic crops, mainly because in the past, medicinal plants were mostly sourced from the wild, with fewer incidents of rust diseases and thus had not received much attention. However, as the cultivation area of medicinal plants continues to increase, rust diseases have gradually become one of the important diseases affecting the quality and yield of medicinal materials. Attention on rust diseases in medicinal plants have been steadily increasing over time [17].
A total of 79 rust species were found to cause diseases on 76 species of medicinal plants from 33 families in China [16,17]. Rust diseases have become the primary diseases on some important medicinal plants in their primary growth regions, with strong prevalence and large damage areas, such as safflower rust disease (Puccinia carthami), Japanese yam rust (Puccinia dioscoreae), Radix glehniae rust (Puccinia phellopteri), and Rust on bulbus fritillariae ussuriensis (Uromyces aecidiiformis) [117,118,119,120]. Most researches on rusts on medicinal plants have been focused on the descriptions of symptoms, the incidence scopes and geographic distribution and the rough morphological descriptions of some spores [17]. However, high morphological variations, wide host range and complicated life cycles, identification of rust fungi is very difficult solely based on morphologies or host specificity. Herein, with the aid of morphological and molecular data, ten rust species have been found on medicinal plants collected from Guizhou province, including three new species and six known species. Among them, Hamaspora rubi-alceifolii, Nyssopsora altissima and Phragmidium cymosum were introduced as new to science. Neophysopella ampelopsidis, Phragmidium tormentillae was firstly introduced in Chinese mainland. Melampsora laricis-populina, Melampsoridium carpini, and Nyssopsora koelreuteriae were documented for the first time in Guizhou province. The accurate identification of rust fungi on medicinal plants will lay the foundation for disease control of medicinal plants.
The use of DNA sequences is becoming more and more important in the identification of rust fungi. Despite early research on rust fungi, the taxonomic system remains perplexing [121]. Distinguishing between individual rust fungi based solely on morphology is challenging [13,122,123]. Because the vast majority of rust fungi cannot be cultured on the artificial medium, pure culture strain cannot be obtained. Therefore, there is no enough DNA sequence available for a large number of rust species for a long time. As DNA extraction techniques continue to improve, valid DNA sequences will become increasingly available [61], and phylogenetic and morphological-based approaches will resolve the taxonomic confusion in rust fungi [11,124]. There were 337 species of rust fungi in 76 genera of 14 families using both morphological and molecular data from 86 natural reserves and national parks in the past five years [11]. Because molecular phylogenetic approaches can be used to connect the telial and aecial stages of rust fungi, they used more additional characters for species recognition [125]. Thus, their studies using DNA-based phylogenetic approach have facilitated precise identification of rust fungi at familial, generic, and species level. These studies can present a significant contribution to the knowledge of rust flora in China, especially those on medicinal plants.
Author Contributions
Conceptualization, Q.L. and L.L.; investigation, Q.W., L.L. and H.H.; morphological examinations, molecular sequencing, and phylogenetic analyses, Q.W. and M.H.; specimen identification, T.L., P.Z. and Q.W.; writing—original draft preparation, Q.W.; writing—review and editing, P.Z. and T.L.; supervision, Q.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Guizhou Province Ordinary Colleges and Universities Youth Science and Technology Talent Growth Project [2021]154; National Natural Science Foundation of China (31960005, 32000009, and 32170019); Guizhou Provincial Education Department Scientific Research Project for Higher Education Institutions ([2022]064).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All newly generated sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank/, accessed on 16 December 2022; Table 1). All new taxa were deposited in MycoBank (https://www.mycobank.org/, accessed on 3 January 2023; MycoBank identifiers follow new taxa).
Acknowledgments
The authors would like to extend their heartfelt gratitude to the State Key Laboratory of Functions and Applications of Medicinal Plants, Guizhou Medical University, Science and Technology Department of Guizhou Province, Mycological Herbarium of Chifeng University, and Nalin N. Wijayawardene for their invaluable guidance and support in enhancing the English writing of this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kolmer, J.A.; Ordonez, M.E.; Groth, J.V. The rust fungi. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2009. [Google Scholar]
- Rhiney, K.; Guido, Z.; Knudson, C.; Avelino, J.; Bacon, C.M.; Leclerc, G.; Aime, M.C.; Bebber, D.P. Epidemics and the future of coffee production. Proc. Natl. Acad. Sci. USA 2021, 118, e2023212118. [Google Scholar] [PubMed]
- Zatybekov, A.; Genievskaya, Y.; Rsaliyev, A.; Maulenbay, A.; Yskakova, G.; Savin, T.; Turuspekov, Y.; Abugalieva, S. Identification of quantitative trait loci for leaf rust and stem rust seedling resistance in bread wheat using a genome–wide association Study. Plants 2021, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.; Tisserant, E.; Talhinhas, P.; Azinheira, H.; Vieira, A.; Petitot, A.S.; Loureiro, A.; Poulain, J.; Da Silva, C.; Silva Mdo, C.; et al. 454–pyrosequencing of Coffea arabica leaves infected by the rust fungus Hemileia vastatrix reveals in planta–expressed pathogen–secreted proteins and plant functions in a late compatible plant–rust interaction. Mol. Plant Pathol. 2012, 13, 17–37. [Google Scholar]
- Chen, Z. Investigation of coffee leaf rust (Hemileia vastatrix) physiological races in China. Mycosystema 1998, 17, 21–28. [Google Scholar]
- Li, L.; Várzea, V.M.P.; Xia, Q.; Xiang, W.; Tang, T.; Zhu, M.; He, C.; Pereira, A.P.; Silva, M.C.; Wu, W.; et al. First report of Hemileia vastatrix (Coffee leaf rust) physiological races emergent in coffee germplasm collections in the coffee–cropping regions of China. Plant Dis. 2021, 105, 4162. [Google Scholar] [CrossRef] [PubMed]
- Cummins, G.B.; Hiratsuka, Y. Illustrated Genera of Rust Fungi, 2nd ed.; American Phytopathological Society (APS Press): St Paul, MN, USA, 1983. [Google Scholar]
- Aime, M.C. Toward resolving family–level relationships in rust fungi (Uredinales). Mycoscience 2006, 47, 112122. [Google Scholar]
- Spatafora, J.W.; Aime, M.C.; Grigoriev, I.V.; Martin, F.; Stajich, J.E.; Blackwell, M. The fungal tree of life: From molecular systematics to genome-scale phylogenies. In The Fungal Kingdom; Wiley: Hoboken, NJ, USA, 2017; pp. 1–34. [Google Scholar]
- Aime, M.C.; Bell, C.D.; Wilson, A.W. Deconstructing the evolutionary complexity between rust fungi (Pucciniales) and their plant hosts. Stud. Mycol. 2018, 89, 143–152. [Google Scholar] [CrossRef]
- Zhao, P.; Zhang, Z.F.; Hu, D.M.; Tsui, K.M.; Qi, X.H.; Phurbu, D.; Gafforov, Y.; Cai, L. Contribution to rust flora in China I, tremendous diversity from natural reserves and parks. Fungal Divers. 2021, 110, 1–58. [Google Scholar]
- Zhuang, J.Y.; Wei, S.X.; Wang, Y.C. Flora Fungorum Sinicorum; Science Press: Beijing, China, 1998; Volume 10. [Google Scholar]
- McTaggart, A.R.; Shivas, R.G.; van der Nest, M.A.; Roux, J.; Wingfield, B.D.; Wingfield, M.J. Host jumps shaped the diversity of extant rust fungi (Pucciniales). New Phytol. 2016, 209, 1149–1158. [Google Scholar] [CrossRef]
- Laundon, G.F.A. Coleosporium rust new to New Zealand. Plant Dis. Rep. 1978, 62, 796–797. [Google Scholar]
- Demers, J.E.; Liu, M.; Hambleton, S.; Castlebury, L.A. Rust fungi on Panicum. Mycologia 2017, 109, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Delate, K.; McKern, A.; Burcham, R. Evaluation of organic soybean rust treatments for organic production neely–kinyon trial, 2005. Iowa State Univ. Res. Demonstr. Farms Prog. Rep. 2006, 2005, 1. [Google Scholar]
- Yu, Z.L.; Yang, J.; Lei, M.Y.; Quan, J.; Yang, T.J.; Yang, C.Q. Status quo of and challenges for research on rust disease in medicinal plants. Zhongguo Zhong Yao Za Zhi-China J. Chin. Mater. Medica 2021, 46, 3566–3576. (In Chinese) [Google Scholar]
- Zou, T.C. Research on sustainable utilisation of medicinal plant germplasm resources in Guizhou. J. Pharm. Pract. 2000, 5, 316–323. (In Chinese) [Google Scholar]
- Xia, Z.M.; Zhang, Z.M. Preliminary report on the survey of pest and weed species and damage of medicinal plants in Guizhou Province. Plant Dr. 2003, 16, 26–28. (In Chinese) [Google Scholar]
- Xia, Z.P.; Hu, X.P.; Wei, C.S. The occurrence and control of pepper rust in the top altar of Guizhou Province. Mod. Agric. Technol. 2018, 16, 117. (In Chinese) [Google Scholar]
- Huang, J.; Zang, Z.G.; Huang, R.M.; Song, B.A. Survey and research on diseases of key medicinal plants in Guizhou Province. Agrochemicals 2002, 10, 6–9. [Google Scholar]
- Wei, G.R.; Wang, X.J. Purification and storage of Puccinia striiformis f. sp. Tritici. J. Henan Agric. Sci. 2011, 40, 90–92. [Google Scholar]
- Hiratsuka, Y.; Hiratsuka, N. Morphology of spermogonia and taxonomy of rust fungi. Rep. Tottori Mycol. Inst. 1980, 18, 257–268. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Beenken, L.; Lutz, M.; Scholler, M. DNA barcoding and phylogenetic analyses of the genus Coleosporium (Pucciniales) reveal that the North American goldenrod rust C. solidaginis is a neomycete on introduced and native Solidago species in Europe. Mycol. Prog. 2017, 16, 1073–1085. [Google Scholar] [CrossRef]
- Colmán, A.; Evans, H.; Salcedo, S.; Braun, U.; Belachew–Bekele, K.; Barreto, R. A fungus–eat–fungus world: Digitopodium, with particular reference to mycoparasites of the coffee leaf rust, Hemileia vastatrix. IMA Fungus 2021, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Capella–Gutiérrez, S.; Silla–Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large–scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Glez–Peña, D.; Gómez–Blanco, D.; Reboiro–Jato, M.; Fdez–Riverola, F.; Posada, D. ALTER: Program–oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, W14–W18. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML–VI–HPC: Maximum likelihood–based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; Volume 2010, pp. 1–8. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Nylander, J.J.E.B.C. MrModeltest v2. Program Distributed by the Author, Version 2; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A.; Drummond, A. FigTree: Tree Figure Drawing Tool, v1. 4.2; University of Edinburgh: Edinburgh, UK, 2012. [Google Scholar]
- Feau, N.; Vialle, A.; Allaire, M.; Maier, W.; Hamelin, R.C. DNA barcoding in the rust genus Chrysomyxa and its implications for the phylogeny of the genus. Mycologia 2011, 103, 1250–1266. [Google Scholar] [CrossRef]
- Zhao, P.; Qi, X.H.; Crous, P.W.; Duan, W.J.; Cai, L. Gymnosporangium species on Malus: Species delineation, diversity and host alternation. Persooniai 2020, 45, 68–100. [Google Scholar] [CrossRef]
- Shen, Y.M.; Chung, W.H.; Huang, T.C.; Rodeva, R.; Hung, T.H. Unveiling Gymnosporangium corniforme, G. unicorne, and G. niitakayamense sp. nov. in Taiwan. Mycoscience 2017, 59, 218–228. [Google Scholar] [CrossRef]
- Cao, B.; Tao, S.; Tian, C.; Liang, Y. Coleopuccinia in China and its relationship to Gymnosporangium. Phytotaxa 2018, 347, 235–242. [Google Scholar] [CrossRef]
- Aime, M.C.; McTaggart, A.R. A higher–rank classification for rust fungi, with notes on genera. Fungal Syst. Evol. 2021, 7, 21–47. [Google Scholar] [CrossRef]
- McKenzie, E.H.C.; Padamsee, M.; Dick, M. First report of rust on Alnus in New Zealand is Melampsoridium betulinum, not M. hiratsukanum. Plant Pathol. Quar. 2013, 3, 59–65. [Google Scholar] [CrossRef]
- Blomquist, C.L.; Scheck, H.J.; Haynes, J.; Woods, P.W.; Bischoff, J. First published report of rust on white alder caused by Melampsoridium hiratsukanum in the United States. Plant Dis. 2014, 98, 155. [Google Scholar] [CrossRef]
- Bubner, B.; Buchheit, R.; Friedrich, F.; Kummer, V.; Scholler, M. Species identification of European forest pathogens of the genus Milesina (Pucciniales) using urediniospore morphology and molecular barcoding including M. woodwardiana sp. nov. MycoKeys 2019, 48, 1–40. [Google Scholar] [CrossRef]
- Vialle, A.; Feau, N.; Frey, P.; Bernier, L.; Hamelin, R.C. Phylogenetic species recognition reveals host–specific lineages among poplar rust fungi. Mol. Phylogenetics Evol. 2013, 66, 628–644. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, Q.H.; Tian, C.M.; Wang, Q.; Kakishima, M. Melampsora salicis–bakko, a new species on willows in Japan evidenced by morphological and molecular phylogenetic analyses. Mycol. Prog. 2016, 15, 13. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, Q.H.; Tian, C.M.; Kakishima, M. Integrating a numerical taxonomic method and molecular phylogeny for species delimitation of Melampsora species (Melampsoraceae, Pucciniales) on willows in China. PLoS ONE 2015, 10, e0144883. [Google Scholar] [CrossRef]
- Deadman, M.L.; Al–Sa’di, A.M.; Al–Maqbali, Y.M.; Al–Jahdhami, S.; Patzelt, A.; Aime, M.C. First report of the rust Melampsora euphorbiae on Euphorbia heterophylla in Oman. J. Plant Pathol. 2006, 88, 229. [Google Scholar]
- Wang, L.L.; Li, K.M.; Liu, Y.; Tian, C.M. Melampsora salicis–michelsonii sp. nov. on Salix michelsonii and Melampsora salicis–cavaleriei on Salix serrulatifolia from China. Phytotaxa 2020, 435, 280–292. [Google Scholar] [CrossRef]
- Zheng, W.; Newcombe, G.; Hu, D.; Cao, Z.; Yu, Z.; Peng, Z. The first record of a north American poplar leaf rust fungus, Melampsora medusae, in China. Forests 2019, 10, 182. [Google Scholar] [CrossRef]
- Ali, B.; Sohail, Y.; Toome–Heller, M.; Mumtaz, A.S. Melampsora pakistanica sp. nov.; a new rust fungus on Euphorbia helioscopia (Sun spurge) from Pakistan. Mycol. Prog. 2016, 15, 1285–1292. [Google Scholar] [CrossRef]
- Vialle, A.; Feau, N.; Allaire, M.; Didukh, M.; Martin, F.; Moncalvo, J.M.; Hamelin, R.C. Evaluation of mitochondrial genes as DNA barcode for Basidiomycota. Mol. Ecol. Resour. 2009, 9, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Pei, M.H.; Bayon, C.; Ruiz, C. Phylogenetic relationships in some Melampsora rusts on Salicaceae assessed using rDNA sequence information. Mycol. Res. 2005, 109, 401–409. [Google Scholar] [CrossRef]
- Chatasiri, S.; Ono, Y. Phylogeny and taxonomy of the Asian grapevine leaf rust fungus, Phakopsora euvitis, and its allies (Uredinales). Mycoscience 2008, 49, 66–74. [Google Scholar] [CrossRef]
- Pota, S.; Chatasiri, S.; Unartngam, J.; Yamaoka, Y.; Hosaka, K.; Ono, Y. Taxonomic identity of a Phakopsora fungus causing the grapevine leaf rust disease in Southeast Asia and Australasia. Mycoscience 2014, 56, 198–204. [Google Scholar] [CrossRef]
- Beenken, L. Pucciniales on Annona (Annonaceae) with special focus on the genus Phakopsora. Mycol. Prog. 2014, 13, 791–809. [Google Scholar] [CrossRef]
- Maier, W.; McTaggart, A.R.; Roux, J.; Wingfield, M.J. Phakopsora myrtacearum sp. nov.; a newly described rust (Pucciniales) on eucalypts in eastern and southern Africa. Plant Pathol. 2016, 65, 189–195. [Google Scholar] [CrossRef]
- Dixon, L.J.; Castlebury, L.A.; Aime, M.C.; Glynn, N.C.; Comstock, J.C. Phylogenetic relationships of sugarcane rust fungi. Mycol. Prog. 2010, 9, 459–468. [Google Scholar] [CrossRef]
- Alaei, H.; De Backer, M.; Nuytinck, J.; Maes, M.; Höfte, M.; Heungens, K. Phylogenetic relationships of Puccinia horiana and other rust pathogens of Chrysanthemum× morifolium based on rDNA ITS sequence analysis. Mycol. Res. 2009, 113, 668–683. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, B.; Tao, S.; Tian, C.; Liang, Y. Phragmidium species parasitizing species of Rosaceae in Tibet, China, with descriptions of three new species. Mycol. Prog. 2018, 17, 967–988. [Google Scholar] [CrossRef]
- Yun, H.Y.; Minnis, A.M.; Kim, Y.H.; Castlebury, L.A.; Aime, M.C. The rust genus Frommeëlla revisited: A later synonym of Phragmidium after all. Mycologia 2011, 103, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Maier, W.; Begerow, D.; Weiß, M.; Oberwinkler, F. Phylogeny of the rust fungi: An approach using nuclear large subunit ribosomal DNA sequences. Can. J. Bot. 2003, 81, 12–23. [Google Scholar] [CrossRef]
- Yang, T.; Chang, W.; Cao, B.; Tian, C.; Zhao, L.; Liang, Y. Two new Phragmidium species identified on Rosa plants native to China. Phytotaxa 2015, 217, 182–190. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, Y.M.; Ono, Y. Taxonomic revision of species of Kuehneola and Phragmidium on Rosa, including two new species from China. Mycologia 2020, 112, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Deadman, M.L.; Al Sadi, A.M.; Al Maqbali, Y.M.; Farr, D.F.; Aime, M.C. Additions to the rust fungi (Pucciniales) from northern Oman. Sydowia 2011, 63, 155–168. [Google Scholar]
- Sun, J.E.; Zhang, Q.; Luo, W.M.; Yang, Y.Q.; An, H.M.; Wang, Y. Four new Phragmidium (Phragmidiaceae, Pucciniomycetes) species from Rosaceae plants in Guizhou Province of China. MycoKeys 2022, 93, 193–213. [Google Scholar] [CrossRef]
- Wilson, A.W.; Aime, M.C. The rose rust fungus, Phragmidium tuberculatum, is widespread in the Americas: First reports from California, Oregon, Massachusetts, and Honduras. Plant Dis. 2014, 98, 1581. [Google Scholar] [CrossRef]
- Liang, Y.M.; Tian, C.M.; Kakishima, M. Phylogenetic relationships on 14 morphologically similar species of Pucciniastrum in Japan based on rDNA sequence data. Mycoscience 2006, 47, 137–144. [Google Scholar] [CrossRef]
- Beenken, L.; Zoller, S.; Berndt, R. Rust fungi on Annonaceae II: The genus Dasyspora Berk. MA Curtis. Mycologia 2012, 104, 659–681. [Google Scholar] [CrossRef] [PubMed]
- McTaggart, A.R.; Doungsa–Ard, C.; Geering, A.D.W.; Aime, M.C.; Shivas, R.G. A co–evolutionary relationship exists between Endoraecium (Pucciniales) and its Acacia hosts in Australia. Persoonia 2015, 35, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Beenken, L.; Wood, A.R. Puccorchidium and Sphenorchidium, two new genera of Pucciniales on Annonaceae related to Puccinia psidii and the genus Dasyspora. Mycol. Prog. 2015, 14, 49. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Y.; Yu, Y.; Bi, C. First report of rust disease on Koelreuteria bipinnata caused by Nyssopsora koelreuteriae in China. Plant Dis. 2016, 100, 1014. [Google Scholar] [CrossRef]
- Baiswar, P.; Ngachan, S.V.; Chandra, S. Identification of Nyssopsora thwaitesii on Schefflera in northeast India. Australas. Plant Dis. Notes 2014, 9, 124. [Google Scholar] [CrossRef]
- Cummins, G.B. Uredinales of continental China collected by SY Cheo. I. Mycologia 1950, 42, 779–797. [Google Scholar] [CrossRef]
- Monoson, H.L. The species of Hamaspora. Mycopathol. Mycol. Appl. 1969, 37, 263–272. [Google Scholar] [CrossRef]
- Chen, Z.C. Additional materials for a rust-flora of Taiwan. Taiwania 1980, 25, 185–186. [Google Scholar]
- Zhuang, J.Y.; Wei, S.X.; Wang, Y.C. Flora Fungorum Sinicorum; Science Press: Beijing, China, 2012; Volume 41. [Google Scholar]
- Yao, Z.S.; Yang, W.L. The medicinal plants of the genus Rubus in Jiangxi Province and suggestions for their utilization. J. Chin. Med. Mater. 1995, 11, 551–554. (In Chinese) [Google Scholar]
- Ye, H.Z.; Hong, Z.F.; Wang, Y.H.; Chen, Y.H. Study of the therapeutic effect of Rubus alceaefolius on experimental liver injury. Chin. Arch. Tradit. Chin. Med. 2005, 23, 829–831. (In Chinese) [Google Scholar]
- Niero, R.; Filho, V.C. Therapeutic potential and chemical composition of plants from the genus Rubus: A mini review of the last 10 years. Nat. Prod. Comm. 2008, 3, 437–444. [Google Scholar] [CrossRef]
- Zhao, H.Y.; Zhao, J.Y.; Liu, Y.; Zheng, Y.Q.; Wu, J.; Hong, Z.F. Effect of total alkaloids of Rubus alceaefolius on oxidative stress in rats with non–alcoholic fatty liver disease. China J. Chin. Mater. Medica 2011, 36, 2383–2387. (In Chinese) [Google Scholar]
- Li, J.; Du, L.F.; He, Y.; Yang, L.; Li, Y.Y.; Wang, Y.F.; Gao, X.M. Chemical constituents and biological activities of plants from the genus Rubus. Chem. Biodivers. 2015, 12, 1809–1847. [Google Scholar] [CrossRef]
- Kuprevich, V.J.; Tranzchel, V.G. Cryptogamic Plants of the USSR, IV, Fungi (1), Rust Fungi No. 1. Family Melampsolaceae; Translated from Russian (1970); Israel Program for Scientific Translations: Jerusalem, Israel, 1957; pp. 362–379. [Google Scholar]
- Spiers, A.G.; Hopcroft, D.H. Comparative studies of the poplar rusts Melampsora medusae, M. larici–populina and their interspecific hybrid M. medusae–populina. Mycol. Res. 1994, 98, 889–903. [Google Scholar] [CrossRef]
- Yue, Y.J. Populus lasiocarpa leaves for diabetic foot ulcers in 5 cases of observation and care. Diabetes New World 2015, 35, 142. (In Chinese) [Google Scholar]
- Yue, Y.J. Care of one case of severe pressure ulcer treated with leaf dressing of Populus lasiocarpa. Chin. J. Misdiagn. 2008, 8, 8003. (In Chinese) [Google Scholar]
- Hiratsuka, N.; Sato, S.; Katsuya, K.; Zhao, M.; Hiratsuka, Y.; Kaneko, S.; Ono, Y.; Sato, T.; Harada, Y.; Hiratsuka, T. The Rust Flora of Japan; Tsukuba Shuppankai: Tsukuba-shi, Japan, 1992. [Google Scholar]
- Spaulding, P. Foreign Diseases of Forest Trees of the World; US Department of Agriculture: Washington, DC, USA, 1961; Volume 197.
- Zhuang, J.Y.; Wei, S.X.; Wang, Y.C. Flora Fungorum Sinicorum; Science Press: Beijing, China, 2005; Volume 25. [Google Scholar]
- Son, Y.K.; Yoon, S.R.; Bang, W.Y.; Bae, C.H.; Yeo, J.H.; Yeo, R.; An, J.; Song, J.; Kim, O.Y. Carpinus turczaninowii extract modulates arterial inflammatory response: A potential therapeutic use for atherosclerosis. Nutr. Res. Pract. 2019, 13, 302. [Google Scholar] [CrossRef] [PubMed]
- Cieckiewicz, E.; Angenot, L.; Gras, T.; Kiss, R.; Frederich, M. Potential anticancer activity of young Carpinus betulus leaves. Phytomedicine 2012, 19, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Qianqian, S.; Fang, X.; Zhu, Z.; Xiao, W.; Wang, Z.; Ding, G.; Zhao, L.; Li, Y.; Yu, P.; Ding, Z.; et al. Seasonal variation of pheophorbide a and flavonoid in different organs of two Carpinus species and its correlation with immunosuppressive activity. Vitr. Cell. Dev. Biol.-Anim. 2016, 52, 654–661. [Google Scholar]
- Majewski, T. Flora polska. In Grzyby (Mycota) 9: Basidiomycetes, Uredinales I; PWN: Warszawa, Poland, 1977. [Google Scholar]
- Zhuang, W.Y.; Guo, Y.L.; Wen, H.A.; Zhang, X.Q.; Lin, G.; Ning, Z.; Zhuang, J.Y.; Wei, S.X. Fungal flora of the Daba Mountains: Uredinales. Mycotaxon 1997, 61, 49–79. [Google Scholar]
- Zhuang, J.Y.; Wei, S.X.; Wang, Y.C. Flora Fungorum Sinicorum; Science Press: Beijing, China, 2021; Volume 62. [Google Scholar]
- Xi, Z.H.; Liu, X.J.; Mu, M.J.; Liang, Q.Y. Studies on the chemical composition of Ampelopsis sinica root. Chin. Tradit. Herb. Drugs 1992, 23, 611. (In Chinese) [Google Scholar]
- Pang, R.; Tao, J.; Zhang, S.-L.; Chen, K.-L.; Zhao, L.; Yue, X.; Wang, Y.-F.; Ye, P.; Hua, D.; Zhu, Y.; et al. Corrigendum to “Ethanol Extract from Ampelopsis sinica Root Exerts Anti–Hepatitis B Virus Activity via Inhibition of p53 Pathway In Vitro”. Evid. –Based Complement. Altern. Med. 2015, 2015, 603232-1. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; Huang, B.S.; Cao, Y.; Chen, K.L.; Li, J. Anti–hepatoma activities of ethyl acetate extract from Ampelopsis sinica root. Oncol. Rep. 2017, 37, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y. Taxonomy of the Phakopsora ampelopsidis species complex on vitaceous hosts in Asia including a new species, P. euvitis. Mycologia 2000, 92, 154–173. [Google Scholar] [CrossRef]
- Monoson, H. The species of Triphragmium, Nyssopsora and Triphragmiopsis. Mycopathologia 1974, 52, 115–131. [Google Scholar] [CrossRef]
- Ye, S.Y.; Lu, J.; Luo, S.M. Biological characteristics of Ailanthus altissima and its exploitation. Beijing Agric. 2013, 33, 79–80. (In Chinese) [Google Scholar]
- Tusculum; Li, T.; Wang, Y.L.; Zhao, M. Study on the chemical composition of the roots of Ailanthus altissima. Chin. Tradit. Herb. Drugs 2015, 46, 1426–1430. (In Chinese) [Google Scholar]
- Peng, S.; Yi, Z.; Liu, M. Ailanthone: A new potential drug for castration–resistant prostate cancer. Chin. J. Cancer 2017, 36, 25. [Google Scholar] [CrossRef]
- Bao, X.Z.; Yv, X.C.; Niu, Z.S.; Fu, L.; Li, Q. Effect of Ailanthinone on endogenous AR of prostate cancer 22RV1 cells. Northwest Pharm. J. 2022, 37, 62–66. (In Chinese) [Google Scholar]
- Li, X.; Jiang, W.B.; Weng, M.L. Landscape characteristics of Koelreuteria bipinnata and its development and utilization. Chin. Agric. Sci. Bull. 2009, 25, 141–146. (In Chinese) [Google Scholar]
- Cao, L.M.; Tuo, J.Y.; He, J.; Ten, T.; Cao, M. nalysis of fatty acid and amino acid composition of seed kernels of Koelreuteria bipinnata. China Oils Fats 2020, 45, 134–137. (In Chinese) [Google Scholar]
- Tranzschel, W. Contribution à l’étude du genre Triphragmium auct. (Triphragmium Link, Triphragmiopsis Naumov, Nyssopsora Arthur). J. Soc. Bot. Russ. 1925, 8, 132. [Google Scholar]
- Zhuang, J.Y. A general survey of rust flora in the Mt. Namjagbarwa region. Mauntain Res. 1984, 2, 198–203. [Google Scholar]
- Zhang, L.W.; ZHhao, J.N.; Xu, J.W. Study on the anti–inflammatory effect of aqueous extracts of Rosa cymosa roots. J. Jilin Agric. Univ. 2010, 32, 37–39. (In Chinese) [Google Scholar]
- Ouyang, L.M.; Huang, S.C.; Huang, X.L.; Huang, Y. Pharmacodynamic study on the anti–inflammatory and analgesic, hemostatic and intestinal motility inhibiting effects of aqueous extracts of roots and stems of Rosa minor. Mod. Chin. Med. 2012, 14, 4–8. (In Chinese) [Google Scholar]
- Fan, X.; Bai, L.; Lin, L.; Liao, D.; Gong, Y.; Liu, X.; Wang, Z.; Li, C. Studies on the chemical constituents and quality evaluation of Rosa cymosa Tratt. root. J. Sep. Sci. 2020, 43, 4379–4389. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wang, L.; Jin, F.; Li, L.; Wang, T.; Gao, M.; Yang, X. Neuroprotection of Kaji–Ichigoside F1 via the BDNF/Akt/mTOR Signaling Pathways against NMDA–Induced Neurotoxicity. Int. J. Mol. Sci. 2022, 23, 16150. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.X. Flora of China; Science Press: Beijing, China, 1990; Volume 37. [Google Scholar]
- Wu, J.; Zhang, Z.Q.; Yu, H.H.; Huang, F.B.; Chen, Z.L.; Chu, L.L.; Li, B.; Wang, W. Research progress on chemical constituents and pharmacological activities of Potentilla. Zhongguo Zhongyao Zazhi-China J. Chin. Mater. Med. 2022, 47, 1509–1538. (In Chinese) [Google Scholar]
- Juszczak, K.; Adamowicz, J.; Zapała, Ł.; Kluz, T.; Adamczyk, P.; Wdowiak, A.; Bojar, I.; Misiek, M.; Grzybowska, M.E.; Stangel–Wójcikiewicz, K.; et al. Potentilla chinensis aqueous extract attenuates cyclophosphamide–induced hemorrhagic cystitis in rat model. Sci. Rep. 2022, 12, 13076. [Google Scholar] [CrossRef]
- McKenzie, E.H.C.; Dingley, J.M. New plant disease records in New Zealand: Miscellaneous fungal pathogens III. N. Z. J. Bot. 1996, 34, 263–272. [Google Scholar] [CrossRef]
- Lye, K.A. Norwegian rust fungi-The genus Frommeëlla. Agarica 2012, 35, 59–66. [Google Scholar]
- Zhao, L.S.; Tian, X.J.; Zang, S.X.; Li, Y.Q.; Zhang, Z.M. Discovery of pycnospore stage of Puccinia phellopteri and study on rust of glehnia littoralis. Acta Phytopathol. Sin. 1996, 26, 49. [Google Scholar]
- Fu, J.F.; Zhou, R.J.; Yan, X.R.J. Occurrence of the rust disease on Dioscorea nipponica Makino in Liaoning Province. Plant Prot. 2004, 30, 90–92. [Google Scholar]
- Zhu, R.J.; Zheng, C.J. Characteristics and control measures of rust disease of Fritillaria ussuriensis. China Plant Prot. 2008, 28, 32–33. (In Chinese) [Google Scholar]
- Liao, A.P. Study on safflower rust caused by Puccinia carthami (Hutz) Corda. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2008. [Google Scholar]
- Schultze–Kraft, R.; Cook, B.; Ciprián, A. Clearing confusion in Stylosanthes taxonomy. 2. S. macrocephala M.B. Ferreira & Sousa Costa vs. S. capitata Vogel and S. bracteata Vogel. Trop. Grassl. –Forrajes Trop. 2020, 8, 250–262. [Google Scholar]
- Uchida, J.; Zhong, S.; Killgore, E. First report of a rust disease on ohia caused by Puccinia psidii in Hawaii. Plant Dis. 2006, 90, 524. [Google Scholar] [CrossRef]
- Ono, Y. Life cycle and nuclear behavior in three rust fungi (Uredinales). Mycoscience 2002, 43, 0037–0045. [Google Scholar] [CrossRef]
- Piškur, B.; Jurc, D. New findings of rusts (Pucciniales) on trees and other plants in Slovenia. In Proceedings of the Zbornik Predavanj in Referatov, 13. Slovenskega Posvetovanja o Varstvu Rastlin z Mednarodno Udeležbo, Rimske Toplice, Slovenia, 7–8 March 2017; Volume 2017, pp. 222–229. [Google Scholar]
- Zhao, P.; Li, Y.; Li, Y.; Liu, F.; Liang, J.; Zhou, X.; Cai, L. Applying early divergent characters in higher rank taxonomy of Melampsorineae (Basidiomycota, Pucciniales). Mycology 2022, 14, 11–36. [Google Scholar] [CrossRef]
Figure 1.
RAxML tree of the family Melampsoraceae based on rDNA ITS and LSU sequence. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to Rossmanomyces monesis and Chrysomyxa empetri [11]. The type specimens are shown as boldface. New sequences are in red.
Figure 1.
RAxML tree of the family Melampsoraceae based on rDNA ITS and LSU sequence. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to Rossmanomyces monesis and Chrysomyxa empetri [11]. The type specimens are shown as boldface. New sequences are in red.

Figure 2.
The RAxML tree of the family Neophysopellaceae and Phakopsoraceae based on rDNA ITS and LSU sequences. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G. asiaticum and G. clavariiforme [11]. The type specimens are shown as boldface. New sequences are in red.
Figure 2.
The RAxML tree of the family Neophysopellaceae and Phakopsoraceae based on rDNA ITS and LSU sequences. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G. asiaticum and G. clavariiforme [11]. The type specimens are shown as boldface. New sequences are in red.

Figure 3.
RAxML tree of the family Phragmidiaceae based on rDNA ITS and LSU sequences. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G. asiaticum and G. clavariiforme [11]. The type specimens are shown as boldface. New sequences are in red.
Figure 3.
RAxML tree of the family Phragmidiaceae based on rDNA ITS and LSU sequences. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G. asiaticum and G. clavariiforme [11]. The type specimens are shown as boldface. New sequences are in red.

Figure 4.
RAxML tree of the family Pucciniastraceae and Hyalopsoraceae based on rDNA ITS and LSU sequence. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G. asiaticum and G. clavariiforme [11]. New sequences are in red.
Figure 4.
RAxML tree of the family Pucciniastraceae and Hyalopsoraceae based on rDNA ITS and LSU sequence. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to G. asiaticum and G. clavariiforme [11]. New sequences are in red.

Figure 5.
The RAxML tree of the families Gymnosporangiaceae, Sphaerophragmiaceae and Uredinineae incertae sedis based on rDNA ITS and LSU sequences. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to Melampsoridium botulinum [40]. The type specimens are shown as boldface. New sequences are in red.
Figure 5.
The RAxML tree of the families Gymnosporangiaceae, Sphaerophragmiaceae and Uredinineae incertae sedis based on rDNA ITS and LSU sequences. ML bootstrap supports (≥70%) and Bayesian posterior probability (≥0.90) are indicated as ML/BYPP. The tree is rooted to Melampsoridium botulinum [40]. The type specimens are shown as boldface. New sequences are in red.

Figure 6.
Hamaspora rubi-alceifolii (GMB0109). (A,B) host and its habitat; (C–F) telia on the hypophyllous leaf and stem; (G) teliospore cluster; (H–L) fusiform teliospores with 5–6 septa. Scale bars: (D) = 1 mm; (E–G) = 0.5 mm; (H–L) =10 µm.
Figure 6.
Hamaspora rubi-alceifolii (GMB0109). (A,B) host and its habitat; (C–F) telia on the hypophyllous leaf and stem; (G) teliospore cluster; (H–L) fusiform teliospores with 5–6 septa. Scale bars: (D) = 1 mm; (E–G) = 0.5 mm; (H–L) =10 µm.

Figure 7.
Melampsora laricis-populina (GMB0097) (A,B) host and its habitat; (C–E) uredinia; (F–K) urediniospores and paraphyses. Scale bars: (D) = 1 mm; (E) = 0.1 mm; (F–K) = 10 µm.
Figure 7.
Melampsora laricis-populina (GMB0097) (A,B) host and its habitat; (C–E) uredinia; (F–K) urediniospores and paraphyses. Scale bars: (D) = 1 mm; (E) = 0.1 mm; (F–K) = 10 µm.

Figure 8.
Melampsoridium carpini (GMB0112) (A,B) host; (C–F) uredinia; (G–L) urediniospores. Scale bars: (E,F) = 0.1 mm; (G–L) = 10 µm.
Figure 8.
Melampsoridium carpini (GMB0112) (A,B) host; (C–F) uredinia; (G–L) urediniospores. Scale bars: (E,F) = 0.1 mm; (G–L) = 10 µm.

Figure 9.
Neophysopella ampelopsidis (GMB0110). (A,B) host and its habitat; (C–F) uredinia.; (G–L) urediniospores and paraphyses. Scale bars: (D–F) = 0.1 mm, (G–L) = 10 µm.
Figure 9.
Neophysopella ampelopsidis (GMB0110). (A,B) host and its habitat; (C–F) uredinia.; (G–L) urediniospores and paraphyses. Scale bars: (D–F) = 0.1 mm, (G–L) = 10 µm.

Figure 10.
Nyssopsora altissima (GMB0103). (A,B) host and its habitat; (C,D) telia on the amphigenous leaf; (E,F) telia; (G–I) teliospores; (J,K) urediniospores with handle. Scale bars: (E,F) = 0.1 mm; (G–K) = 10 µm.
Figure 10.
Nyssopsora altissima (GMB0103). (A,B) host and its habitat; (C,D) telia on the amphigenous leaf; (E,F) telia; (G–I) teliospores; (J,K) urediniospores with handle. Scale bars: (E,F) = 0.1 mm; (G–K) = 10 µm.

Figure 11.
Nyssopsora koelreuteriae (GMB0105). (A,B) Host and its habitat. (C–F) Telia (G–K) Teliospores on the stem. Scale bars: (E,F) = 0.1 mm; (G–K) = 10 µm.
Figure 11.
Nyssopsora koelreuteriae (GMB0105). (A,B) Host and its habitat. (C–F) Telia (G–K) Teliospores on the stem. Scale bars: (E,F) = 0.1 mm; (G–K) = 10 µm.

Figure 12.
Phragmidium cymosum (GMB0108). (A,B) host and its habitat; (C,D) uredinia on the hypophyllous leaf surfaces; (E,F) uredinia; (G–L) globose or obovoid urediniospores with echinulate spines or with abnormal protrusions and paraphyses. Scale bars: (D) = 1 mm; (E) = 0.1 mm; (F) = 0.5 mm; (G–L) = 10 µm.
Figure 12.
Phragmidium cymosum (GMB0108). (A,B) host and its habitat; (C,D) uredinia on the hypophyllous leaf surfaces; (E,F) uredinia; (G–L) globose or obovoid urediniospores with echinulate spines or with abnormal protrusions and paraphyses. Scale bars: (D) = 1 mm; (E) = 0.1 mm; (F) = 0.5 mm; (G–L) = 10 µm.

Figure 13.
Phragmidium rosae-roxburghii (GMB0104). (A,B) host and its habitat; (C,D) uredinia on the hypophyllous leaf surfaces; (E,F) uredinia; (G–K) globose or oval urediniospores with echinulate spines and paraphyses. Scale bars: (E,F) = 0.5 mm, (G–K) = 10 µm.
Figure 13.
Phragmidium rosae-roxburghii (GMB0104). (A,B) host and its habitat; (C,D) uredinia on the hypophyllous leaf surfaces; (E,F) uredinia; (G–K) globose or oval urediniospores with echinulate spines and paraphyses. Scale bars: (E,F) = 0.5 mm, (G–K) = 10 µm.

Figure 14.
Phragmidium tormentillae (GMB0114). (A) host; (B–F) uredinia; (G–J) urediniospores. Scale bars: (C–F) = 0.1 mm; (G–J) = 10 µm.
Figure 14.
Phragmidium tormentillae (GMB0114). (A) host; (B–F) uredinia; (G–J) urediniospores. Scale bars: (C–F) = 0.1 mm; (G–J) = 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
GenBank accession number and information of taxa used for phylogenetic analyses.
| Family Name | Species | Specimen No. | Host | Province, Country | GenBank Accession No. | Reference | |
|---|---|---|---|---|---|---|---|
| ITS | LSU | ||||||
| Chrysomyxaceae | Chrysomyxa empetri | QFB 25015 | Empetrum nigrum | Ste-Anne-des-Monts, Quebec, Canada | GU049434 | GU049526 | [36] |
| Rossmanomyces monesis | DAOM 221982 | Moneses (=Pyrola) uniflora | Graham Island, British Columbia, Canada | GU049476 | GU049547 | [36] | |
| Gymnosporangiaceae | Gymnosporangium asiaticum | CUP-0016 | Juniperus chinensis | Gifu prefecture, Japan | MN642593 | MN642617 | [37] |
| Gymnosporangium clavariiforme | BRIP 59471 | Crataegus sp. | — | — | MW049261 | [10] | |
| Gymnosporangium confusum | LD 1021 | Crataegus monogyna | Turkey | HM114219 | HM114219 | — | |
| Gymnosporangium cupressi | RSP9998 | — | — | KJ720169 | KJ720169 | — | |
| Gymnosporangium juniperi-virginianae | MCA3585 | Cupressaceae juniperus | Virginia, America | — | MG907217 | [10] | |
| Gymnosporangium sabinae | TNM F0030477 | Pyrus communis | Bulgaria: Sofia | KY964764 | KY964764 | [38] | |
| Gymnosporangium trachysorum | RN91 | — | — | KJ720184 | KJ720184 | — | |
| Hyalopsoraceae | Coleopuccinia sinensis | BJFC:R02506 | Cotoneaster microphyllus | China | — | MF802285 | [39] |
| Hyalopsora aspidiotus | PUR N4641 | Gymnocarpium dryopteris | China | — | MW049264 | [40] | |
| Melampsoridium alni | H 7019539 | Alnus mandshurica | Finland | KF031557 | KF031534 | [41] | |
| Melampsoridium betulinum | BPI 871107 (MCA2884) | Alnus sp. | Costa Rica | — | DQ354561 | [8] | |
| Melampsoridium hiratsukanum | PDR #1480181 | Alnus rhombifolia | USA: Santa Cruz County, California | KC313888 | KC313888 | [42] | |
| Melampsoridium carpini | KR-M-0048587 | Carpinus betulus | — | MH908486 | MH908486 | [43] | |
| Melampsoridium carpini | GMB0112 | Carpinus turczaninowii | Guizhou, China | OQ067091 | — | This study | |
| Melampsoraceae | Melampsora abietis-canadensis | 1399MEA-POG-USA | Populus grandidentata | Wisconsin, America | JN881733 | JN934918 | [44] |
| Melampsora abietis-populi | TSH-R20042 * | Populus yunnanensis | Yunnan, China | JN881739 | JN934933 | [44] | |
| Melampsora apocyni | LYR3 | Apocynum venetum | Xinjiang, China | KR296802 | KR296803 | — | |
| Melampsora caprearum | NYS-F-003819 | Salix caprea | Germany | KU550034 | KU550033 | [45] | |
| Melampsora coleosporioides | HNMAP3114 | Salix sp. | Japan | KF780755 | KF780638 | [46] | |
| Melampsora epiphylla | TSH-R3884 | Salix bakko | Japan | KF780787 | KF780670 | [45] | |
| Melampsora epitea | TNS-F-121034 | Salix viminalis | Germany | KX386070 | KX386097 | [35] | |
| Melampsora euphorbiae | HMAS350000 | Euphorbia kansui | Guangxi, China | MK518875 | MK518545 | [11] | |
| Melampsora euphorbiae | BRIP 39560 | Euphorbia peplus | Queensland, Australia | — | EF192199 | [40] | |
| Melampsora euphorbiae | BPI 871135 (U681) | Euphorbia heterophylla | Oman | — | DQ351722 | [47] | |
| Melampsora humilis | TSH-R7550 | Salix koriyanagi | Miyagi, Japan | KF780812 | — | [46] | |
| Melampsora hyperici-sampsonii | HMAS350001 * | Hypericum sampsonii | Guangxi, China | MK518877 | MK518547 | [11] | |
| Melampsora iranica | HMAAC4055 | Salix sp. | China | MK372158 | MK372191 | [48] | |
| Melampsora kamikotica | HNMAP3186 | Chosenia arbutifolia | China | KF780760 | KF780643 | [46] | |
| Melampsora laricis-pentandrae | HNMAP3201 | Salix pentandra | Inner Mongolia, China | KF780801 | KF780684 | [46] | |
| Melampsora laricis-populina | HMAS247977 | Populus simonii | Haixi Qinghai, China | MK028583 | MK064524 | [49] | |
| Melampsora laricis-populina | GMB0097 | Populus lasiocarpa | Guizhou, China | OQ067085 | — | This study | |
| Melampsora larici-tremulae | PFH04-5 | Populus tremula | France | JN881744 | JN934956 | [44] | |
| Melampsora medusae | BPI 0021209 | — | America | JX416848 | JX416843 | [44] | |
| Melampsora medusae f.sp. deltoidis | 98D10 | Populus × euramericana | South Africa | GQ479307 | JN934962 | [44] | |
| Melampsora medusae f.sp. tremuloidis | 1028ME-LAL-LJ | Larix laricina | Quebec, Lac Saint Jean, Peribonka, Canada | GQ479883 | JN934965 | [44] | |
| Melampsora microsora | TSH-R7335 | Salix subfragilis | Hiroshima, Japan | KF780833 | KF780730 | [46] | |
| Melampsora microspora | PUR:F17540 | populus nigra | Iraq | JN881737 | JN934931 | [44] | |
| Melampsora occidentalis | MO96H | Populus trichocarpa | Idaho, America | JN881740 | JN934934 | [44] | |
| Melampsora pakistanica | BA13c | Euphorbia helioscopia | Pakistan | KX237555 | KX237556 | [50] | |
| Melampsora populnea | 664ME-POA-BC45.1 | Populus alba | British Columbia, Canada | EU808021 | FJ666510 | [51] | |
| Melampsora populnea | 97MP10 | — | France | EU808035 | FJ666523 | [51] | |
| Melampsora populnea | AAH00-1 | Populus alba | England | AY444772 | AY444786 | [52] | |
| Melampsora pruinosae | BPI 1109446 | Populus diversifolia | Xinjiang Uygur Zizhiqu, Shule, China | GQ479899 | JN934938 | [44] | |
| Melampsora pulcherrima | O8ZK4 | Mercurialis annua | Italy | GQ479320 | JN934941 | [44] | |
| Melampsora ribesii-purpureae | PURP897-1 | Salix purpurea | — | AY444770 | AY444791 | [52] | |
| Melampsora euphorbiae | PDD 98363 | Ricinus communis | — | — | KJ716352 | [36] | |
| Melampsora rostrupii | 01G1.1 | Mercurialis perennis | France | EU808038 | JN934942 | [44] | |
| Melampsora salicis-albae | NWC-06210 | Salix alba | Rothamsted, England | KF780757 | KF780640 | [46] | |
| Melampsora salicis-argyraceae | HMAS52894 * | Salix argyracea | Xinjiang, China | KF780733 | KF780616 | [46] | |
| Melampsora salicis-bakko | TSH-R3879 * | Salix bakko | — | KC631854 | KC685611 | [45] | |
| Melampsora salicis-cavaleriei | HMAAC4043 | Salix serrulatifolia | China | MK277296 | MK277301 | [48] | |
| Melampsora salicis-delavayanae | HMAS248120 * | Salix delavayana | Yunnan, China | MK518954 | MK518651 | [11] | |
| Melampsora salicis-purpureae | HMAS62584 * | Salix purpurea | Shandong, China | KF780766 | KF780649 | [46] | |
| Melampsora salicis-viminalis | HMAS38658 * | Salix viminalis | Tibet, China | KF780732 | KF780615 | [46] | |
| Melampsora sp. | HMAS350003 | Xinjiang, China | MK518844 | MK518499 | [11] | ||
| Neophysopellaceae | Neophysopella ampelopsidis | IBA-8618 | Ampelopsis brevipedunculata | Kagoshima, Japan | AB354774 | AB354741 | [11] |
| Neophysopella ampelopsidis | GMB0110 | Ampelopsis sinica | Guizhou, China | OQ067090 | — | This study | |
| Neophysopella meliosmae | IBAR-10037 | Meliosma myriantha | Ibaraki, Japan | KC815591 | KC815650 | [53] | |
| Neophysopella meliosmae-myrianthae | AGLR56 | — | — | MK290819 | MK290819 | — | |
| Neophysopella meliosmae-myrianthae | IBA-8584 | Vitis coignetiae | Tochigi, Japan | AB354785 | AB354748 | [53] | |
| Neophysopella orientalis | IBAR-9942 | Meliosma tenuis | Tochigi, Japan | KC815597 | KC815656 | [54] | |
| Neophysopella vitis-davidii | HMAS350005 | Viola faurieana | Chongqing, China | MK518870 | MK518536 | [11] | |
| Neophysopella vitis-davidii | HMAS248115 * | Vitis davidii | Yunnan, China | — | MK518593 | [11] | |
| Phakopsoraceae | Masseeëlla capparis | RIP 56844 | Flueggea virosa | Northern Territory, Australia | JX136798 | JX136798 | [13] |
| Nothoravenelia japonica | HMJAU8598 | — | China | — | MK296509 | — | |
| Phakopsora argentinensis | ZT:RB 8248 | Croton cf. anisodontus | — | KF528009 | KF528009 | [55] | |
| Phakopsora crcis-filii | ZT Myc 48990 | Annona paludosa | Sinnamary, French Guiana | KF528016 | KF528016 | [55] | |
| Phakopsora myrtacearum | PREM 61155 * | Eucalyptus grandis | Maragua, Kenya | NR_132913 | KP729473 | [56] | |
| Phakopsora neocherimoliae | RB 3096 in ZT | Annona cherimola | San José Costa Rica | KF528011 | KF528011 | [55] | |
| Phakopsora pachyrhizi | BRIP 56941 | Neonotonia wightii | Warrumbungle, New South Wales, Australia | — | KP729475 | [56] | |
| Phakopsora phyllanthi | RB 8581 | Phyllanthus acidus | Ceará, Brazil | KF528025 | KF528025 | [55] | |
| Phakopsora pistila | ZT Myc 48992 | Annona sericea | French GuianaIracubo, French Guiana | KF528026 | KF528026 | [55] | |
| Phakopsora sophorae | HMAS248098 * | Leptopus chinensis | Beijing, China | — | MK518628 | [11] | |
| Phragmidiaceae | Arthuriomyces peckianus | BPI 879271 | Rubus occidentalis | New York, America | GU058010 | GU058010 | [57] |
| Gerwasia pittieriana | BPI 843556 | Rubus sp. | — | KY764065 | KY764065 | — | |
| Gerwasia rubi | HMAS249978 | Rubus parkeri | Yunnan, China | MK519039 | MK518737 | [11] | |
| Gerwasia rubi | HMAS249983 | Rubus setchuenensis | Fujian, China | — | MK518734 | [11] | |
| Gerwasia rubi | BRIP:58369 | Rubus sp. | — | — | KT199397 | [13] | |
| Gerwasia rubi-playfairiani | HMAS249840 * | Rubus playfairianus | Guangxi, China | MK518976 | — | [11] | |
| Hamaspora acutissima | BRIP:55606 | Rubus rolfei | Philippines | — | KT199398 | [13] | |
| Hamaspora rubi-alceifolii | GMB0116 | Rubus alceaefolius | Guizhou, China | — | OQ067533 | This study | |
| Hamaspora rubi-alceifolii | GMB0109 * | Rubus alceaefolius | Guizhou, China | — | OQ067532 | This study | |
| Hamaspora longissima | BPI 871506 | Rubus rigidus | Eastern Cape, South Africa | — | MW049262 | [40] | |
| Hamaspora sinica | HMAS249989 | Rubus setchuenensis | Guangdong, China | MK519049 | MK518636 | [11] | |
| Kuehneola japonica | BPI 910185 | Rosa sp. | — | KY764067 | — | — | |
| Kuehneola uredinis | LD1029 | Rubus sp. | New York, America | GU058013 | GU058013 | [57] | |
| Kuehneola uredinis | R216 | Rubus fruticosus | Belgium | EU014068 | EU014068 | [58] | |
| Phragmidium andersonii | HMAS53231 | Potentilla fruticosa | China | — | MG669120 | [59] | |
| Phragmidium barclayi | HMAS67281 | Rubus austrotibetanus | China | — | MG669117 | [59] | |
| Phragmidium barnardii | BRIP:56945 | Rubus multibracteatus | Queensland, Australia: | — | KT199402 | [13] | |
| Phragmidium biloculare | U564 | Potentilla flabellifolia | Washington, America | — | JF907670 | [60] | |
| Phragmidium brevipedicellatum | HMUT100463 | Potentilla multifida | Xinjiang, China | — | KU059170 | — | |
| Phragmidium butleri | HMAS67841 | Rosa macrophylla | China | — | MG669118 | [59] | |
| Phragmidium chayuensis | BJFC:R02532 * | Rosa duplicata | China | MH128374 | NG_064492 | [59] | |
| Phragmidium cibanum | BJFC:R02528 * | Rubus niveus | China | MH128370 | NG_064491 | [59] | |
| Phragmidium cymosum | GMB0115 | Rosa cymosa | Guizhou, China | OQ067097 | OQ067531 | This study | |
| Phragmidium cymosum | GMB0108 * | Rosa cymosa | Guizhou, China | OQ067096 | OQ067530 | This study | |
| Phragmidium duchesnea-indica | HMAS249846 * | Duchesnea indica | Yunnan, China | — | MK518681 | [11] | |
| Phragmidium fragariae | WM 1317 | Potentilla sterilis | — | — | AF426217 | [61] | |
| Phragmidium fructigenum | HMUT100472 | Rosa glomerata | Chongqing, China | — | KU059168 | — | |
| Phragmidium fusiforme | BJFC:R00942 | Rosa hugonis | China | — | KP407632 | [62] | |
| Phragmidium griseum | HMAS56906 | Rubus crataegifolius | Beijing, China | MH128377 | MG669115 | [59] | |
| Phragmidium griseum | HMAS350016 | Rosa sp. | Beijing, China | — | MK518530 | [11] | |
| Phragmidium handelii | BJFC:R01030 | Rosa webbiana | China | — | KP407631 | [62] | |
| Phragmidium ivesiae | U35 | Potentilla gracilis | Utah, America | — | JF907672 | [60] | |
| Phragmidium japonicum | HMAS41585 | Rosa laevigata | Fujian, China | MN264716 | MN264734 | [63] | |
| Phragmidium jiangxiense | BJFCR 03453 * | Rosa laevigata | Jiangxi, China | MN264715 | MN264733 | [63] | |
| Phragmidium kanasense | HMAS248114 * | Rosa fedtschenkoana | Xinjiang, China | — | MK518464 | [11] | |
| Phragmidium leucoaecium | BJFCR 02118 * | Rosa sp. | Yunnan, China | MN264719 | MN264737 | [63] | |
| Phragmidium mexicanum | HMAS350019 | Phoenix acaulis | Yunnan, China | MK518980 | MK518678 | [11] | |
| Phragmidium mexicanum | HMAS350020 | Duchesnea indica | Yunnan, China | MK518982 | MK518680 | [11] | |
| Phragmidium mexicanum | MCA2496 | Potentilla indica | Maryland, America | — | JF907660 | [60] | |
| Phragmidium montivagum | FO 47828 | Rosa cf. woodsii | — | — | AF426213 | [61] | |
| Phragmidium mucronatum | U-671 | Rosa sp. | Oman | — | HQ412646 | [64] | |
| Phragmidium mucronatum | BRIP:60097 | Rosa rubiginosa | — | — | MW049275 | [40] | |
| Phragmidium nambuanum | HMUT100465 | Rubus innominatus | Chongqing, China | — | KU059165 | — | |
| Phragmidium octoloculare | HMAS140416 | Rubus biflorus | China | MH128376 | MG669119 | [59] | |
| Phragmidium pauciloculare | HMUT100466 | Rubus innominatus | Chongqing, China | — | KU059162 | — | |
| Phragmidium pauciloculare | HMAS350023 | Rubus corchorifolius | Guangxi, China | MK518874 | MK518542 | [11] | |
| Phragmidium potentillae | BRIP:60089 | Acaena novae-zelandiae | Tasmania, Australia | — | KT199403 | [13] | |
| Phragmidium potentillae-canadensis | MCA2858 | Potentilla sp. | New York, America | — | JF907666 | [60] | |
| Phragmidium punjabense | BA65A * | Rosa sp. | Murree, Ghora Gali, Pakistan | KX358856 | KX358854 | [50] | |
| Phragmidium rosae-californicae | MVAP0000154 | Rosa californica | Putah Creek Reserve, Davis, California, America | MK045315 | MK045315 | — | |
| Phragmidium rosae-moschatae | BPI 893257 | Rosa macrophylla | — | — | KY798368 | — | |
| Phragmidium rosae-multiflorae | HMAS71053 | Rosa multiflora | Shanxi, China | — | KU059174 | [65] | |
| Phragmidium rosae-multiflorae | HMAS94924 | Rosa multiflora | Zhejiang, China | — | KU059175 | [65] | |
| Phragmidium rosae-multiflorae | BJFCR 03454 * | Rosa multiflora | Jiangxi, China | MN264721 | MN264739 | [63] | |
| Phragmidium rosae-roxburghii | HGUP21025 * | Rosa roxburghii | Guizhou, China | OL684818 | OL684831 | [65] | |
| Phragmidium rosae-roxburghii | GMB0104 | Rosa xanthina | Guizhou, China | OQ067092 | — | This study | |
| Phragmidium rubi-idaei | HMUT100470 | Rubus saxatilis | Chongqing, China | — | KU059163 | — | |
| Phragmidium rubi-oldhami | HMAS64306 | — | China | — | MG669116 | [59] | |
| Phragmidium tormentillae | GMB00114 | Potentilla simulatrix | Guizhou, China | OQ067093 | — | This study | |
| Phragmidium tormentillae | BPI 843392 | Duchesnea sp. | Maryland, America | — | DQ354553 | [8] | |
| Phragmidium tuberculatum | BPI 877977 | Rosa floribunda | Massachusetts, America | — | KJ841923 | [66] | |
| Phragmidium tuberculatum | BPI 877980 | Rosa floribunda | Oregon, America | — | KJ841922 | [66] | |
| Phragmidium violaceum | PDD 99246 | Rubus sp. | New Zealand | — | KJ716351 | [36] | |
| Phragmidium yangii | BJFC:R00338 * | Rosa sp. | China | — | NG_060138 | [62] | |
| Phragmidium zangdongii | BJFC:R02447 * | Rosa tibetica | Tibet, China | MH128372 | NG_064490 | [59] | |
| Phragmidium zhouquense | BJFC:R01516 * | Rosa omeiensis | China | — | NG_060139 | [62] | |
| Trachyspora alchemillae | WM 1019 | Alchemilla vulgaris | — | — | AF426220 | [61] | |
| Trachyspora alchemillae | BPI 843828 | Alchemilla vulgaris | Switzerland | — | DQ354550 | [8] | |
| Xenodochus carbonarius | PUR N15566 | Sanguisorba officinalis | — | — | MW049289 | [40] | |
| Pucciniastraceae | Melampsorella elatina | WM 1092 | Abies alba | — | — | AF426232 | [61] |
| Melampsorella elatina | PUR 82 | Cerastium fontanum | Minnesota, America | — | MG907233 | [10] | |
| Pucciniastrum circaeae | RB 2098 | Circaea lutetiana | — | — | AF426227 | [61] | |
| Pucciniastrum corni | TSH-R13510 | Cornus kuosa | Tottori, Japan | AB221437 | AB221409 | [67] | |
| Pucciniastrum epilobii | PUR N11088(MCA5308) | Epilobium angustifolium | — | — | MW147083 | [40] | |
| Pucciniastrum fagi | TSH-R10724 | Fagus crenata | Tochigi, Japan | AB221425 | AB221378 | [67] | |
| Pucciniastrum hikosanense | TSH-R4287 (IBA2565) | Acer rufinerve | Yamanashi Japan | AB221441 | AB221388 | [67] | |
| Pucciniastrum hydrangeae | MCA2837 | Hydrangea arborescens | North Carolina, America | — | MG907240 | [10] | |
| Pucciniastrum kusanoi | HiroU21509 | Clethra barbinervis | Miyazaki, Japan | AB221426 | AB221402 | [67] | |
| Pucciniastrum yoshinagai | TSH-R4272 IBA8430 | Stewartia monadelpha | Nara, Yoshino-gun, Japan | AB221434 | AB221411 | [67] | |
| Thekopsora minima | MCA2988 | Vaccinium angustifolium | Maryland, America | — | MG907243 | [10] | |
| Sphaerophragmiaceae | Austropuccinia psidii | BRIP:58164 | Rhodamnia angustifolia | Brisbane, Queensland. Australia | — | KF318449 | [67] |
| Dasyspora amazonica | BPI US0116382 * | Xylopia amazonica | Brazil | JF263460 | JF263460 | [68] | |
| Dasyspora gregaria | ZT Myc 3397 | Xylopia cayennensis | French Guiana | JF263477 | JF263477 | [68] | |
| Dasyspora guianensis | ZT Myc 3413 * | Xylopia benthamii | French Guiana | JF263479 | JF263479 | [68] | |
| Dasyspora mesoamericana | PUR 42390 * | Xylopia frutescens | Panama | JF263480 | JF263480 | [68] | |
| Dasyspora segregaria | PMA MP4941 | Xylopia aromatica | Panama | JF263488 | JF263488 | [68] | |
| Dasyspora winteri | S F30078 | Xylopia sericea | Brazil | JF263492 | JF263492 | [68] | |
| Puccorchidium polyalthiae | ZT HeRB 251 | Polyalthia longifolia | India | JF263493 | JF263493 | [68] | |
| Sphaerophragmium acaciae | BRIP:56910 | Albizzia sp. | Kununurra, Western Australia, Australia | — | KJ862350 | [69] | |
| Sphaerophragmium longicorne | PUR N16513 | Dalbergia hostilis | — | — | MW147053 | [40] | |
| Sphenorchidium xylopiae | PC 0096711 | Xylopia aethiopica | western Central Africa. | KM217355 | KM217355 | [70] | |
| Uredinineae incertae sedis | Allodus podophylli | BPI 842277 | Podophyllum peltatum | Maryland, America | DQ354543 | DQ354543 | [67] |
| Neopuccinia bursa | RB 75707 | Protium heptaphyllum | Brazil | — | MH047186 | — | |
| Nyssopsora altissima | GMB0103 * | Ailanthus altissima | Guizhou, China | OQ067089 | OQ067529 | This study | |
| Nyssopsora echinata | KR0012164 | Meum athamanticum | — | — | MW049272 | [40] | |
| Nyssopsora koelreuteriae | BBSW-1 | Koelreuteria bipinnata | — | KT750965 | KT750965 | [71] | |
| Nyssopsora koelreuteriae | GMB0105 | Koelreuteria bipinnata | Guizhou, China | OQ067088 | — | This study | |
| Nyssopsora thwaitesii | AMH:9528 | Schefflera wallichiana | India | KF550283 | — | [72] | |
Notes: —: no data available; *: type specimens.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, Q.; He, M.; Liu, T.; Hu, H.; Liu, L.; Zhao, P.; Li, Q. Rust Fungi on Medicinal Plants in Guizhou Province with Descriptions of Three New Species. J. Fungi 2023, 9, 953. https://doi.org/10.3390/jof9090953
AMA Style
Wu Q, He M, Liu T, Hu H, Liu L, Zhao P, Li Q. Rust Fungi on Medicinal Plants in Guizhou Province with Descriptions of Three New Species. Journal of Fungi. 2023; 9(9):953. https://doi.org/10.3390/jof9090953
Chicago/Turabian StyleWu, Qianzhen, Minghui He, Tiezhi Liu, Hongmin Hu, Lili Liu, Peng Zhao, and Qirui Li. 2023. "Rust Fungi on Medicinal Plants in Guizhou Province with Descriptions of Three New Species" Journal of Fungi 9, no. 9: 953. https://doi.org/10.3390/jof9090953
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.