
Fungal and Fungal-like Diseases of Halophytes in the Mediterranean Basin: A State-of-the-Art Review

1
Department of Agriculture, Food and Environment, University of Pisa, Via del Borghetto 80, 56124 Pisa, Italy
2
Interdepartmental Research Center, Nutraceuticals and Food for Health, University of Pisa, Via del Borghetto 80, 56124 Pisa, Italy
*
Authors to whom correspondence should be addressed.
Horticulturae 2024, 10(4), 313; https://doi.org/10.3390/horticulturae10040313
Submission received: 16 February 2024
/
Revised: 18 March 2024
/
Accepted: 21 March 2024
/
Published: 23 March 2024
(This article belongs to the Special Issue Fungal Diseases and Pathogens Affecting Horticultural Crops: New Challenges)
Abstract
:Halophytes are salt-tolerant plants growing in saline ecosystems and are spread throughout the Mediterranean area. Recently, there has been a renewed interest in agricultural exploitation of halophytes, but poor attention has been given to pest and disease management of these species. The objective of this review is to assess the extent and distribution of pathogenic fungal and fungal-like (Oomycota) organisms on major Mediterranean halophytes. We retrieved 92 references spanning over 100 years, with more than half published since 2000. Ascomycota is the best-represented phylum, and the order Pleosporales has the highest species diversity. The order Pucciniales prevails in Basidiomycota, whereas Peronosporales is the richest order within Oomycota. Most of the pathogenic species have been isolated from aboveground organs, especially from leaves. Portulaca oleracea is the species with the most associated pathogens (16) and records (28). Leveillula powdery mildew, caused by Leveillula taurica, is the most common disease among the selected species. Cakile maritima was found to be vulnerable to different mycotoxigenic Alternaria species. Strengthening the research on diseases of halophytes is essential to successfully grow these species and to evaluate the risks related to the presence of mycotoxigenic species, which is crucial for the effective exploitation of halophytes as crops.
1. Halophytes for Saline Agriculture
The Mediterranean Basin is considered one of the major hotspots for plant biodiversity in the world, counting more than 25,000 plant species [1]. Hot and dry summers and mild and rainy winters characterize the Mediterranean climate. The Mediterranean Region is particularly vulnerable to climate change [2,3], and coastal areas are particularly prone to saltwater intrusion and tidal inundations [4]. The combination of these extreme events, together with inadequate irrigation practices, makes the Mediterranean soils among the most affected by salinity worldwide [5,6].
Halophytes are salt-adapted plant species that can complete their life cycle in high salt conditions; they can tolerate levels of salinity that salt-sensitive plants, otherwise referred to as glycophytes, cannot tolerate. Various morphological, physiological, and biochemical properties enable halophytes to grow in highly saline environments [7]. The main mechanisms of salt tolerance in halophytes include the development of specialized organs like salt glands, regulation of Na+ and Cl− uptake and excretion, and protection against osmotic stresses [8]. Salt glands originate from the epidermis and consist of multiple vacuolated cells, in which the salt is sequestered in response to salt stress [8]. Through the salt glands, different cations and anions are secreted with a high selectivity for Na+ and Cl− [9]. Furthermore, an important strategy employed by halophytes is the use of specialized transporters to sequester toxic ions, such as Na+ and Cl−, within cellular compartments [8]. Finally, osmotic adjustment, through the accumulation of osmolytes in the cytoplasm, is a salt tolerance mechanism utilized by halophytes. Low-molecular-weight compounds such as proline, sugar alcohols, sorbitol, and quaternary ammonium compounds act as osmolytes [10]. Additionally, halophytes often exhibit cross-tolerance between salinity and other abiotic stresses [11].
Halophytes play also a crucial ecological role for their ability to grow in salt-affected soils, and the increasing concern for soil salinization in the Mediterranean Basin has revived the interest in these species. Halophytes can restore damaged and underused environments and transform them into productive land for agriculture, improving agrobiodiversity and preventing soil degradation [12].
The Mediterranean halophytes mainly reside in the Amaranthaceae family, which counts several extremely salt-tolerant species [7]. Other representative families are Brassicaceae, Poaceae, and Portulacaceae. They have been traditionally collected in the wild for human consumption and medicinal uses [13,14]. However, plant harvest in the wild (foraging) presents many risks due to possible microbiological and chemical contamination and appears to be incompatible with the quality and safety standards required by the current food market. Furthermore, halophytic plants are often present in protected areas and their collection would therefore be illegal [15]. Some species are protected in Europe, such as Salicornia veneta, which is included in the European red list of threatened species as a vulnerable plant [16]. Therefore, the production of these species must be entrusted to professional growers capable of cultivating these plants in controlled conditions and with extensive guarantees regarding sanitary safety and more generally, quality [7].
Moreover, halophytes display a broad range of potentialities that are not limited to human consumption. Many species are widely used as forage, especially Atriplex spp. which has proved to provide a better feeding value when cultivated than the spontaneous counterpart [17]. Furthermore, these plants contain many antioxidant metabolites, which contribute to salt tolerance and nutraceutical value [18]. A relatively high content of phenols has been found, among other species, in Suaeda fruticosa (shrubby sea-blite) [19], Atriplex littoralis (grassleaf orache) [20], and Portulaca oleracea (common purslane) [21]. Interestingly, in other species such as Salicornia perennans (glasswort), Lepidium latifolium (broadleaved pepperweed), and Suaeda maritima (sea-blite), the phenolic fraction can be tailored to obtain phenolic-enriched products through saline cultivation [22]. For these reasons, halophytes are now being rediscovered as promising functional food, with notable market perspectives [23].
Some Mediterranean halophytic species are of economic interest. Salicornia is probably the most widely cultivated halophyte. Agricultural exploitation of Salicornia spp. plant is led by Europe and the Near East, generating hundreds of tons per year in export [24]. In Italy and Spain, Soda inermis (Monk’s beard or agretti in Italian) is a crop of interest for both poor arable areas and greenhouse operations, while Crithmum maritimum (sea fennel) and Cakile maritima (sea rocket) are being rediscovered as valuable crops in the Mediterranean Basin [25,26].
To fulfill market demand, year-round production must be guaranteed and, in this scenario, greenhouse cultivation, either in soil or in soilless systems, is the most promising cropping technology for these plants [7]. Soil-less cultivation systems have proved to be very suitable for halophytes, and several species have been investigated for their potential to grow hydroponically with considerable yields (reviewed in [7]). Moreover, due to their intrinsic tolerance to high salinity, halophytes are excellent candidates for integrated marine aquaculture systems (i.e., aquaponics), where they can act in the recovery of saline aquaculture effluents [27]. In recent years, many efforts have been carried out to unravel the suitability of these crops in aquaponic systems [28,29,30,31] but some crucial aspects are still under-investigated, such as the occurrence and control of plant pathogens. The constant presence of water is an ideal condition for a plethora of plant pathogens and the best adapted are probably those belonging to the phylum Oomycota, which includes genera like Globisporangium and Phytophthora. Other plant pathogenic fungi can be found in hydroponic systems, e.g., Fusarium spp., Colletotrichum spp., Rhizoctonia spp., and Verticillium spp. [32].
Similar to glycophytes, halophytes are susceptible to infectious diseases that can occur either in native environments or throughout the entire production process, thus leading to yield loss or foodstuff contamination. Recently, Calabon et al. [33] have extensively reviewed the fungal biodiversity associated with halophytes in salt marshes worldwide; however, most of these fungal species are saprobic.
Notwithstanding the growing interest in the cultivation of halophytes, the research on pathogenic fungal and fungal-like (Oomycota) organisms of these species is very limited. To fill this knowledge gap, this review aims to systematically analyze and sort the state-of-the-art about infectious diseases of major Mediterranean halophytes. A classification of the pathogenic species, at the order level, is provided together with their distribution among hosts. In addition, for each plant genus or species, a dedicated section will deepen the most well-studied pathosystems, examining the major findings about symptomatology, disease development, and the current taxonomic status of the pathogenic species. The work also intends to provide a critical revision of the records, in conformity with the current taxonomy of Fungi and Oomycota.
The current literature on the topic addressed by this review is very limited. Some halophytic species are still considered weeds and the studies have mainly focused on their eradication with biological methods. For species that can be used as food or non-food crops, however, little or no attention is paid to phytopathological problems, and the case studies are extremely limited. Therefore, due to the lack of information, it was not possible to carry out a comprehensive evaluation including specific aspects (e.g., disease severity in response to growing technique or salinity). The case studies reporting data in this regard have been explored in depth in the appropriate sections of this manuscript.
2. Bibliographic Research and Resources Accessed
Fungus–host associations were retrieved using the United States Department of Agriculture Fungal Database [34] as a starting point, and the references were looked at therein. It should be noted that many reports derive from checklists (e.g., lists and indexes of diseases or species annotated in a particular geographic area) and provide no or brief information about disease. Other studies only aimed at addressing species redisposition within a genus and include herbarium specimens previously collected. Therefore, different literature databases (i.e., JSTOR, Google Scholar, SCOPUS, and Web of Science) were searched for research papers regarding pathosystems in halophytic plants, without any temporal limitations. In SCOPUS, we used this searching strategy: TITLE-ABS-KEY (plant species/genus*) AND TITLE-ABS-KEY (disease* OR host* OR pathosystem*) AND TITLE-ABS-KEY (fungal* OR fungal-like*). When no results were found, the input was modified to the more specific string: TITLE-ABS-KEY (plant species/genus* AND pathogenic species/genus*). Synonym names for both hosts and pathogens were considered. Similarly, the other databases were searched using plant species/genus and pathogenic species/genus as input words.
We only accepted records that included: (i) disease confirmed through artificial inoculations; (ii) sampling of symptomatic plants followed by the isolation of the putative agent of disease (such as herbarium specimens); and (iii) records of biotrophic pathogens (i.e., rusts, powdery and downy mildews). Furthermore, to provide a study as complete as possible, we included records without any geographical limitations. The plant species in this study refer to recent classifications of the main Mediterranean halophytes [7] and were selected for their distribution and environmental significance in the Mediterranean area. Other halophytic species were included based on their geographical distribution according to the Global Biodiversity Information Facility database [35] and if they were listed in the salt-tolerant plant database eHALOPH [36]. Table 1 summarizes the plant species considered in this review. The taxonomic nomenclature for fungi and fungal-like organisms and for plant species is based, respectively, on Index Fungorum [37] and [7]. Studies on the taxonomic status of pathogens were regarded as research papers.
3. Plant Pathogens Associated with Halophytes
Across all the consulted databases, we retrieved a total of 92 references reporting the association of fungal and fungal-like microorganisms that are pathogenic, or potentially pathogenic, to halophytic species. The references cover a time frame from 1911 to 2023, with more than half (48/92, 52.17%) of the documents published since the year 2000. Research papers account for 22.7% (10/44) of the total records before 2000 and 64.6% (31/48) in the following years, thus proving a revived interest in fungal and fungal-like diseases of halophytes (Figure 1).
Overall, 62 different species residing in 16 distinctive orders and three Phyla (Ascomycota, Basidiomycota, and Oomycota) were retrieved (Figure 2). Ascomycota is the most represented and diverse phylum, with the highest number of orders (9) and species (37), followed by Basidiomycota (five different orders and 11 species), and Oomycota (two different orders and 14 species). Among the phylum Ascomycota, the order Pleosporales is the richest one, counting 12 different species, and the genus Alternaria is the most represented with five species (A. alternata, A. arborescens, A. brassicae, A. brassicicola, and A. mali). The order Pleosporales is the largest within the class Dothideomycetes and includes 23 families, 332 genera, and more than 4700 species [38].
Several studies were mostly restricted to endophyte communities and reported Pleosporales as the main order associated with halophytes worldwide. Calabon et al. [33] found that Pleosporales was the most recurring order among the fungi inhabiting salt marshes, with 178 taxa recorded.
Similarly, Pleosporales accounted for 28.9% of the total operational taxonomic units found in endophytic and rhizospheric fungal communities associated with roots of Limbarda crithmoides (golden samphire) [39], interestingly, the fraction of Pleosporales increases when moving from the upper dune to the salt marsh. Pleosporales may prevail in the endophytic community regardless of the location. The genera Phoma (order Pleosporales) and Fusarium (order Hypocreales) were reported to be the most frequent in both coastal and inland sites in the province of Alicante (Southeast Spain) [40]. In the nearby region of Andalusia, more than half of the isolates (208 out of 349) collected from different hosts belonged to Pleosporales, with Neocamarosporium, Alternaria, and Preussia being the most frequently isolated genera [41]. Finally, A. alternata was one of the most frequently isolated species from the roots of halophytic species in the Red Sea coast of Egypt [42].
The second most represented order within the phylum Ascomycota is Mycosphaerellales, which belongs to the class Dothideomycetes and includes important plant pathogens typically causing leaf spots. Within this order, the most represented genera are Cercospora and Passalora, both counting two species (C. beticola and C. bizzozeriana, and P. dubia, and P. spegazzinii, respectively)
In the phylum Basidiomycota, the order counting the highest number of species (seven) is Pucciniales, represented by two genera: Puccinia (two species, P. aristidae, and P. isiacae) and Uromyces (five species, U. betae, U. chenopodii, U. peckianus, U. salicorniae, and U. salsolae). Members of the Pucciniales are widely known as rusts; they can infect a broad range of economically important crops and display a biotrophic lifestyle.
Finally, the Oomycota, a phylum of fungus-like organisms in the Kingdom Straminipila, counts 14 different species of two different orders (Albuginales, and Peronosporales). The genus Peronospora is the most represented one, with five different species: P. atriplicis-hastatae, P. atriplicis-hortensis, P. minor, P. litoralis, and P. schachtii. This genus consists of species mainly responsible for downy mildew, a foliar disease that is promoted by wet weather conditions and is characterized by whitish patches on the underside of leaves and discolorations on the upper leaf surface.
4. Diversity of Plant Pathogens among Halophytes of the Mediterranean Basin
Overall, we considered 11 plant genera commonly found in the Mediterranean Basin, accounting for 22 different plant species belonging to four families: Amaranthaceae, Apiaceae, Brassicaceae, and Portulacaceae. Amaranthaceae is the most represented family, with six genera: Atriplex, Beta, Salicornia, Salsola, Soda, and Suaeda. In the Apiaceae family, Crithmum and Eryngium are the genera examined, whereas Brassicaceae counts two different genera: Cakile and Lepidium. Finally, Portulaca oleracea (purslane) was considered in the Portulacaceae family. The diversity of plant pathogen orders among the halophytic plant genera is reported in Figure 3.
The order Pucciniales is the most common among all the plant genera, as it is described in seven out of eleven genera, although hosts only belong to Amaranthaceae and Brassicaceae (restricted to the genus Lepidium). Interestingly, these biotrophic pathogens were found in every examined plant genus in Amaranthaceae.
Similarly, other orders including biotrophic species are highly common among the plants. Species in Peronosporales (apart from Globisporangium intermedium and G. irregulare, which do not display a biotrophic lifestyle) and Erysiphales (which includes species causing powdery mildew) are reported, respectively, on five and six plant genera. They are described on Atriplex, Beta, Salicornia (Amaranthaceae), Cakile, Lepidium (Brassicaceae), and Beta, Salsola, Soda (Amaranthaceae), Cakile, Lepidium (Brassicaceae), and Crithmum (Apiaceae).
Leaf spot agents in the order Mycosphaerellales are reported on plants belonging to Amaranthaceae (Atriplex and Beta), Apiaceae (Eryngium), and Brassicaceae (Cakile and Lepidium). Surprisingly, despite being the richest order in terms of species diversity, fungal species in the Pleosporales are only recorded on four plant genera: Atriplex, Beta, Cakile, and Portulaca. Genera Cakile and Portulaca count the highest number (four) of distinctive species within this order.
Members of the Diaporthales are present on Amaranthaceae (Atriplex and Salsola), Apiaceae (Eryngium), and Brassicaceae (Lepidium), while species belonging to Glomerellales are present on Amaranthaceae (Salsola), Brassicaceae (Lepidium) and Portulacaceae.
White rusts in the Albuginales are exclusive to the Brassicaceae and Portulacaceae families, while species within the order Helotiales are reported on genera Eryngium and Portulaca. Finally, the remaining orders (Botryosphaeriales, Hypocreales, Microascales—phylum Ascomycota—Cantharellales, Entylomatales, Thelephorales, and Urocystidiales—phylum Basidiomycota) are exclusive to only one host species.
Table 2 shows the association between a pathogenic (or potentially pathogenic) species and its host, reporting the diseases caused by the pathogen and thus providing an overview of the diseases. For those reports providing no information about the pathosystems, the putative disease was inferred by the pathogen itself.
4.1. Atriplex spp.
In the genus Atriplex (Amaranthaceae), five different species are considered: A. hortensis (garden orache), A. littoralis (grassleaf orache), A. nummularia (oldman saltbush), A. patula (common orache), and A. prostrata (hastate orache). Except for A. nummularia, all the species are native to the Mediterranean area. A. nummularia was introduced in Tunisia from Australia and rapidly became one of the most distributed exotic Atriplex species in the whole Mediterranean Region [137]. Fourteen different species are reported on this genus, belonging to the orders Diaporthales, Mycosphaerellales, Peronosporales, Pleosporales, and Pucciniales. The order Peronosporales is the richest one, with four different species described among the selected hosts, each of them resulting to be species-specific.
Interestingly, the different species within the Peronosporales order reside in the genus Peronospora. This genus is the largest in the Oomycota phylum, comprises about 400 species [43], and is responsible for downy mildew on several crops. Atriplex spp. are particularly susceptible to downy mildew. The species separation within the genus Peronospora has undergone several revisions during the decades. Fries [138] first published in Systema Mycologicum the name Botrytis farinosa to indicate an organism found on an undefined species of Atriplex in Sweden before moving it to the genus Peronospora, and therefore establishing the name P. farinosa Fr. [139]. Later, Gaumann [140] described P. litoralis Gäum. on A. littoralis and P. minor Gäum. on A. patula. In 1932, Săvulescu and Rayss [141] described P. atriplicis-hastatae Săvul. & Rayss on A. prostrata (indicated as A. hastata) and P. atriplicis-hortensis Săvul. & Rayss on A. hortensis. Constantinescu [43] kept their original names when annotating an exhaustive list of species in Peronosporales and described 11 species on Atriplex spp. However, Yerkes and Shaw [142] gathered the different Peronospora species, including the parasitic species on other plant hosts in the Amaranthaceae, under the synonym P. farinosa, which was erroneously considered as the sole downy mildew agent on Amaranthaceae. Nonetheless, molecular phylogenetics demonstrated that downy mildews are extremely host-specific and that species from different plants represent different lineages according to their host genus or species [143,144,145]. Taxonomic revisions can lead to the reclassification of particular species and their placement in different taxonomic groups. This affects how pathogenic species are identified. By accurately classifying species within their taxonomic groups, evolutionary history, transmission dynamics, and host range of pathogens can be better defined.
Pathogenic species described on Atriplex spp. are not related to other pathogens revealed on important crops in the Amaranthaceae, such as spinach, beet, and quinoa [146]. Different names were proposed to indicate downy mildew agents on these crops: P. effusa (Grev.) Rabenh. on spinach [144], P. schachtii Fuckel on beet [47], and P. variabilis Gäum. on quinoa [147]. The name P. farinosa was proposed for rejection [148], and the proposal was accepted in 2017 by the Nomenclature Committee on Fungi and the General Committee on Nomenclature [149,150]. Consequently, reports annotating the occurrence of P. farinosa on Atriplex were not included in this study. Through multi-locus phylogenetic analysis, Choi et al. [47] were able to discriminate different downy mildew species associated with Amaranthaceae. The authors discriminated different Peronospora species (including three species on Atriplex spp.) consistently with the narrow species concept, that a downy mildew species is specific to a host plant genus or species. On Atriplex, two distinct lineages were found representing P. litoralis and P. minor/P. atriplicis-hastatae. The study found that the isolates from Atriplex were closely related but received low support for monophyly. They were divided into three clades (i.e., monophyletic groups, comprising members descending from a single ancestor), corresponding to a specific Atriplex species, namely A. littoralis, A. prostrata, and A. patula. Therefore, we only included records aligned with these findings. It should be noted that the identity of P. atriplicis-hortensis was not investigated and thus we have kept the record on A. hortensis [43]. According to the current nomenclature of Index Fungorum, P. minor is not considered a distinct species from P. arborescens. However, following [47], who have deepened the taxonomic status of Peronospora species on Atriplex, we have kept the record as P. minor.
Against this background, we report P. atriplicis-hortensis on A. hortensis [43]; P. litoralis on A. littoralis [43,47]; P. minor on A. patula [43,47,49,50,51,52,53]; and P. atriplicis-hastatae on A. prostrata [43,47].
The Atriplex/Peronospora binomial has a long history of records. The species described on Atriplex seem to be extremely specialized and exclusive of one host species, further confirming the narrow species concept in downy mildews. Molecular phylogenetics is a powerful tool to infer the evolutionary history and relation of different species and is fundamental for understanding the taxonomy, host range, and specificity of closely related organisms. These findings are essential for disentangling the host–pathogen interactions and the complexity of host range in downy mildews. In the optic of agricultural use of this halophyte, these works are a breakthrough in developing accurate diagnosis and pest control protocols.
Another important pathogen frequently associated with Atriplex spp. is Ascochyta caulina (Pleosporales). van der Aa and van Kesteren [44] examined several specimens collected all over Europe and grouped different species names under the synonym A. caulina. However, these authors stated that the isolates showed different conidial shapes depending on the host organ: conidia found on leaves and associated with leaf spots were bigger than those found on stems. Moreover, leaf conidia were hyaline to yellowish green and often 1-septate, occasionally 2- and 3-septate, while those on the stem were yellowish to pale brown and only 1-septate. Thus, due to their different phenotypes, the authors claimed that they may be different species. A. caulina is the causal agent of leaf spot and stem necrosis, and it was reported on stem and leaves of A. prostrata (indicated as A. hastata), stem and leaves of A. hortensis, and leaves of A. littoralis and A. patula [44]. In artificial inoculation trials, the application of A. caulina conidia successfully resulted in leaf necrosis on A. patula and A. prostrata within one week [151].
Later, Andrianova and Minter [46] described another fungal pathogen, Stagonospora atriplicis, as a leaf spot agent on A. hortensis, A. littoralis, A. patula, and A. prostrata. They described the disease as the presence of numerous, irregular, and small spots starting from central or basal leaves on the stem, mainly occurring during the summer. According to Andrianova and Minter [46], the name S. atriplicis and its synonyms (Septoria atriplicis and Phyllosticta atriplicis) have been misunderstood in many studies and are listed among other synonyms of A. caulina [44,152]. The authors stated that differences based on the conidial shape (multi-septate, holoblastically produced from annellated conidiogenous cells in S. atriplicis; 1-septate, enteroblastically produced from phialides in A. caulina) are key features in differentiating the two species. Interestingly, by analyzing two isolates (CBS 246.79 and CBS 344.78) of A. caulina from A. prostrata (reported as A. hastata), de Guyter et al. [153], through molecular phylogeny based on the analysis of actin sequence data, proposed a final identification of Pleospora calvescens for the A. caulina isolate CBS 246.79 and Pleospora chenopodii for the isolate CBS 344.78. However, in a recent taxonomic revision based on a multi-gene phylogenetic analysis, both isolates were designated as Neocamarosporium chenopodii (anamorph A. caulina) [154]. Since both A. caulina and S. atriplicis are still accepted taxa, we have kept the original names provided by the authors. In addition, some authors noted Ascochyta atriplicis as another leaf spot agent on A. prostrata [54,62].
Other pathogenic species known to cause leaf spots on Atriplex are Passalora spegazzinii (Mycosphaerellales) on A. hortensis [45] and A. patula [55,59], and Passalora dubia (Mycosphaerellales) on A. patula [54,55,56,57,58].
Among the fungi belonging to the phylum Basidiomycota, Uromyces peckianus (Pucciniales) is described on A. patula in Canada as the agent of rust [45]. The author, by annotating an index of plant diseases, referred to a study by Fraser [61], where aecia of U. peckianus were collected and successfully inoculated on A. patula plants. Finally, A. nummularia was found to be susceptible to shoot blight caused by Diaporthe incospicua (Diaporthales), which is the only disease annotated on this species. [48].
4.2. Beta vulgaris subsp. maritima
Beta vulgaris subsp. maritima (Amaranthaceae, sea beet) is the only species considered in the genus Beta, for which we retrieved five fungus–host associations. Overall, all the pathogens belong to different orders and cause foliar diseases, although with a different array of symptoms. The most frequently reported species is the rust agent Uromyces betae (Pucciniales). This biotroph was described twice on B. vulgaris subsp. maritima by the Norwegian botanist and mycologist Halvor B. Gjærum in the Madeira archipelago, Portugal [65,66].
Moreover, like cultivated varieties of B. vulgaris, sea beet is susceptible to beet downy mildew (BDM) caused by Peronospora schachtii. BDM is a widespread disease, and it is commonly found on beet [47]. Gustavsson [53] described P. schachtii on this species in Spain and Portugal.
Another important pathogen is Cercospora beticola (Mycosphaerellales), a leaf spot agent. Despite being a very common pathogen of beet, we retrieved only one record [63]. Cercospora leaf spot is considered one of the most devastating foliar diseases on this crop. Dark and grey spots on leaves and petioles characterize this disease, which, at an advanced stage, can cause the collapse of the whole leaf [155].
Powdery mildew caused by Leveillula taurica (Erysiphales) is another foliar disease recorded on sea beet in Italy and Corsica (France) [64]. The genus Leveillula comprises obligate parasites with a broad range of hosts, described on approximately one thousand different plant species, especially members of Solanaceae [156]. Morphologically, Leveillula species are very close to each other, and this has led to confusion in the species definition. It was proved that L. taurica s. lat. is a species-complex and not a monophyletic group [123]. Heluta and Minter [157] advocated that the isolates of L. taurica s. lat. described by Amano [64] on sea beet are probably L. cylindrospora. Leveillula powdery mildew is commonly spread in Central and West Asia and in the Mediterranean area [158]. Typical symptoms are observed on the lower side of affected leaves, which are characterized by abundant and white sporulation, whereas, on the upper side, the infection can result in chlorotic spots that eventually coalesce leading to leaf shredding.
4.3. Cakile maritima
Cakile maritima (Brassicaceae, sea rocket), the sole species considered in the genus Cakile, is an annual, succulent herb that thrives in sand dune habitats [159]. On this halophyte, five different orders have been noted (Albuginales, Erysiphales, Mycosphaerellales, Peronosporales, and Pleosporales) for a total of eight different species.
In general, C. maritima is highly susceptible to Alternaria spp. infections, as four different species are described on this host (A. arborescens, A. alternata, A. brassicicola, and A. mali). The Alternaria spp.-C. maritima pathosystems have been extensively investigated [67,68,69,70]. The best-known and most widely investigated species associated with this halophyte is Alternaria brassicicola (Pleosporales) which causes black necrotic lesions on many parts of the host. On the surface of infected tissues, the pathogen produces spores that are airborne or dispersed by water splash [70]. Alternaria brassicicola has been successfully demonstrated for vertical transmission of the disease from one generation of C. maritima to another, as it was frequently isolated from seeds and fruit coats [67]. Seed dispersal seems to play a key role in the spread of this pathogen [70]. This pathogenic species has also been isolated from flowers, although attacks on fruits have been shown to provide a higher fraction of pathogen inoculum on seeds. Sampling of C. maritima fruits yielded 30% of infected seeds, while up to 60% of seedlings were found to be infected when germinating from attacked fruits [67]. Moreover, plant age and geographical location strongly influence the incidence of the disease. The fraction of infected plants is the highest in adult plants and gradually decreases in young plants and seedlings. In addition, the disease incidence was found to be higher when moving from the sea toward the beaches and the surrounding areas. Thus, a positive relationship between the distance from the sea and disease incidence has been demonstrated [70]. Another pathogenic Alternaria species that was frequently recovered from the Australian population of C. maritima is A. alternata [67]. Nevertheless, this species accounts only for a small fraction of the infected plants, thus resulting in a lower incidence of the disease compared with A. brassicicola. In glasshouse inoculation trials, A. brassicicola was able to infect both adult plants and seedlings, whereas A. alternata was not able to develop any symptoms in seedlings. Moreover, when both species were inoculated on fruits, the fraction of seeds infected by A. brassicicola was significantly higher than those infected by A. alternata. Therefore, A. brassicicola displayed higher colonization fitness and was able to exclude A. alternata [67]. The aggressiveness of A. brassicicola towards its host is so high that disease incidence can reach 100% by the end of the growing season. Although initial disease spread mainly relies on infected overwintering adults or plant debris, since Alternaria is a polycyclic pathogen, outbreaks can reach critical levels during the growing season. The occurrence and distribution of the diseases are deeply dependent on the behavior of pathogenic species. The polycyclic nature of the pathogen characterizes the epidemiology of the disease with several extinction and colonization events [70]. Furthermore, the Australian population of A. brassicicola infecting C. maritima is genetically highly variable. From five different populations of C. maritima, 222 isolates were found representing 202 unique genotypes, and this high level of variability may suggest the occurrence of a sexual recombination process in A. brassicicola [160].
On the other hand, the Tunisian population of Alternaria infecting C. maritima is different. Field surveys of symptomatic plants revealed that the species most frequently associated with black lesions was A. alternata, followed by A. arborescens and A. mali, whereas no A. brassicicola isolates were found [68]. All the tested isolates were proved to be pathogenic after artificial inoculation trials by application of a conidia suspension on leaves [68]; the isolates tested for pathogenicity resulted mycotoxigenic.
Mycotoxins are secondary metabolites produced by several fungal species and represent a threat to food security as exposure can cause health complications. The main mycotoxins produced by Alternaria species are: alternariol (AOH); alternariol monomethyl ether (AME); altuene (ALT), which belong to the group of dibenzopyrone derivatives; alterotoxins (ATXs); and tenuazonic acid (TeA). Some fungal species can even produce host-specific mycotoxins [161]. The Alternaria species pathogenic on C. maritima were positive for AOH, AME, ALT, and TeA biosynthesis with a different spectrum of production [68].
In artificial inoculation trials in saline conditions, the presence of salt (200 mM NaCl) was associated with a lower susceptibility to A. alternata infection [69]. This is the sole study where a pathosystem involving a halophytic species was investigated in the presence of salt. Interestingly, when both saline and biotic stress were applied, plants exhibited a higher growth rate and photosynthesis compared to single-stress application. The key to the alleviation of salt stress over fungal attack might reside in the intrinsic ability of halophytes to tolerate salinity and hence, after exposure to the stress, the defense mechanisms implemented by the plants (e.g., improved scavenging of reactive oxygen species) might act towards other stresses [69]. On the other hand, the susceptibility to salinity of fungal and fungal-like species must be considered as some species can tolerate this stress better than others (see Section 6 for a discussion of salt stress in fungal and fungal-like organisms).
Furthermore, the phylum Oomycota comprises other important pathogens described on C. maritima. Occurrences of Albugo candida (Albuginales) on sea rocket were reported in Egypt [73], Wales [50], Spain, and Portugal [74]. A. candida is a biotrophic plant pathogen widely known as the causal agent of white rust or white blister on Brassicaceae, with an impressive range of hosts. The disease is characterized by the formation of white blisters (or sori) on the infected tissues, usually leaves, representing the zoosporangia bearing zoospores, which are dispersed either by air or water splash [162]. It should be noted that A. candida was proposed to be a species-complex with at least two different clades [163,164]; however, no specimens of A. candida on C. maritima were investigated. Hyaloperonospora parasitica (Peronosporales) is another member of the phylum Oomycota, which was found to cause downy mildew on C. maritima in Egypt and therein reported as Peronospora parasitica [71]. This species was accommodated into the novel genus Hyaloperonospora by Constantinescu and Fatehi [165].
4.4. Crithmum maritimum
Crithmum maritimum (Apiaceae) is a perennial herb that thrives on the Mediterranean coasts. It is a facultative halophyte that is gaining a lot of interest in recent years for both nutrition and medicinal use, due to its high content of essential oils [166,167].
Leveillula powdery mildew is the only disease noted on this species, and the causal agents are L. lanuginosa and L. taurica. Powdery mildew caused by L. taurica was recorded in France [64,76] and Israel [75]. The same authors also reported the presence of L. lanuginosa as a powdery mildew agent on C. maritimum [75].
4.5. Eryngium maritimum
Eryngium maritimum (Apiaceae, sea holly or sea eryngo) is a perennial species of flowering plants native to most European coastlines whose leaves and roots can be used as a food source. On this halophyte, four different fungal orders are recorded (Diaporthales, Entylomatales, Helotiales, and Mycosphaerellales). Each order includes only one species, with Entylomatales (comprising the sole species Entyloma eryngii-maritimi) being unique to this species [79].
The genus Entyloma (phylum Basidiomycota) counts more than 160 species, responsible for leaf smut. This disease is characterized by the presence of spots, which represent sori bearing teliospores. Eryngium spp. are particularly susceptible to Entyloma leaf smut, and the fungal species are highly host-specific [79,168]. Four host-specific species of Entyloma were described by Savchenko et al. [79], including E. eryngii-maritimii. This novel species was established by analyzing specimens collected on E. maritimum in France and Wales. A few distinctive features of E. eryngii-maritimii make this species easily distinguishable from the others: the production of small-size spores (the smallest within the genus) and greenish gray sori.
Diaporthe angelicae (Diaporthales) is noted as the causal agent of stem necrosis on E. maritimum [80,81,82]. This species is a common pathogen described in the family Apiaceae [169]. In both the above-mentioned publications, the fungus was reported with a different name, i.e., as Diaporthopsis angelicae by Ellis and Ellis [80], and as Diaporthe nigrella by Unamuno [81], which are now both considered synonyms of D. angelicae [169,170].
Finally, two leaf spot agents are described on this halophyte: Cylindrosporium eryngii (Helotiales) [77], which was first described by Ellis and Kellerman [171] as a leaf spot agent on Eryngium yuccifolium Michx., and Pseudocercosporella eryngii (Mycosphaerellales) [54,78] reported as Septoria eryngii (now considered a synonym).
4.6. Lepidium spp.
In the genus Lepidium (Brassicaceae), four different species have been considered: L. latifolium (broadleaved pepperweed), L. perfoliatum (clasping pepperweed), L. ruderale (stinking pepperweed), and L. subulatum. Fourteen different pathogenic species, accounting for nine different orders (Albuginales, Diaporthales, Erysiphales, Glomerellales, Mycosphaerellales, Peronosporales, Pleosporales, Pucciniales, and Urocystidiales) were retrieved. This genus includes several species that have a worldwide distribution and generally are weeds [172]. However, some species, such as L. latifolium, have edible leaves with a high content of nutraceutical compounds [173,174].
Overall, all four plant species are susceptible to downy mildew. On L. subulatum, Perofascia lepidii is described as the causal agent of the disease [53]; it is reported as Peronospora lepidii, subsequently accommodated into the Perofascia genus by Constantinescu and Fatehi [166], the sole pathogen described on this species. P. lepidii is also described on L. latifolium [83] and L. ruderale [51,83]. On the latter species, Hyaloperonospora parasitica is another downy mildew agent [51], whereas on L. perfoliatum, Hyaloperonospora lepidii-perfoliati is reported as the causal agent of this disease [43].
Powdery mildew is another common disease of Lepidium spp; this disease is described in three of the Lepidium species considered, and the main causal agent is Erysiphe cruciferarum. This plant pathogen was found on L. latifolium [54,62,64,76,86,87] L. perfoliatum [64,76] and L. ruderale [54,64,76,93]. Powdery mildew caused by E. cruciferarum commonly occurs on many Brassicaceae crops and is characterized by the presence of a powdery whitish coat on the upper side of infected leaves, representing conidiophores bearing conidia which are air-dispersed [175]. Leveillula taurica is reported as another powdery mildew agent on L. latifolium [64].
In addition, the white rust agent Albugo candida is reported on L. latifolium [74,89], and L. ruderale [54]. This disease is widespread in Europe and, through artificial inoculations, Koike et al. [89] showed that the A. candida isolate they found was highly host-specific. Indeed, while the first symptoms of white blister appeared on L. latifolium within 14 days, no disease was noted on other economically important crops such as rocket, mustard, cauliflower, and broccoli. Therefore, Koike et al. [89] concluded that the isolate does not belong to any of the previously described races of the pathogen. Another evidence of the high host specificity of the white rust agents Albugo spp. is provided by Rector et al. [90]. These authors characterized a pathogenic isolate of A. lepidii that was able to infect L. latifolium while not causing any symptoms on rocket, cauliflower, broccoli, pak choi, and radish. The isolate has been proposed as a bioherbicide to control the spread of broad-leaved pepperweed. Bioherbicides are herbicidal agents, derived from living organisms or natural sources, specifically targeted to control and regulate the growth of weeds in agricultural and ecological environments [176]
Within the phylum Basidiomycota, we report the leaf smut agent Urocystis coralloides (Urocystidiales) on L. perfoliatum and L. ruderale [91] and two rusts (Pucciniales), Uredo isiacae on L. latifolium [88] and Puccinia aristidae, on L. perfoliatum [59].
Lepidium perfoliatum is also reported as a host of Verticillium dahliae (Glomerellales) [92], while on L. latifolium leaf spot agents such as Alternaria brassicae [84] and Cercospora bizzozeriana (Mycosphaerellales) [85], and the canker and dieback agent Cytospora ribis (Diaporthales) [82] are described.
4.7. Portulaca oleracea
Portulaca oleracea (Portulacaceae) is the one with the most associated records. We retrieved the highest number (nine) of different fungal and fungal-like orders recorded on this species (Albuginales, Botryosphaeriales, Cantharellales, Glomerellales, Helotiales, Microascales, Pleosporales, Peronosporales and Thelephorales) resulting in 16 different pathogenic species. The orders Botryosphaeriales, Microascales (phylum Ascomycota), Cantharellales, and Thelephorales (phylum Basidiomycota) are unique to this species.
This abundance of records can be attributed to the widespread recognition of P. oleracea as a major noxious weed for various important crops [177]. Nonetheless, this species has succulent stems and leaves with a high content of omega-3 fatty acids and can be consumed moderately as an alternative source of this important fatty acid [178].
The most detailed and well-documented disease on P. oleracea is the white rust caused by Wilsoniana portulacae (Albuginales). By analyzing different specimens through scanning electron microscopy, light microscopy, and ITS phylogeny, the genus Wilsoniana, together with the genus Pustula, was proposed as a new combination apart from the genus Albugo, within the order Albuginales and subclass Albuginomycetidae [179]. White rust of P. oleracea has been widely documented [51,74,77,98,103,112,113,114,115,116,117]. Leaves are the main organs where the infection occurs, although some characteristic symptoms can be occasionally observed on stems. A typical symptom on the upper side of leaves is the presence of white blister-like pustules, which correspond to irregular yellowish chlorotic areas on the lower side. White pustules can be seen on stems as well, but more rarely. If systemic infection occurs, plants display swelled stems and reduced leaf size, which eventually drops [114,115,117]. In addition, it has been shown that, in species of the Albuginaceae family, the development of pustules directly involves the enzymatic lysis of host cell walls, rather than a mechanical process [116]. Lytic enzymes detach the mesophyll from the epidermis, which is then disrupted by the primary sporangia enabling the dispersal of spores from the secondary sporangia [116]
Dichotomophthora lutea is known as a foliar pathogen causing leaf spot. de Hoog and van Oorschot [101] introduced this new species, previously included in the genus Dactylaria, by morphologically analyzing specimens collected from various plant species, including P. oleracea. The identity of the Dichotomophthora species was later confirmed by molecular phylogeny [102]. The multi-locus molecular phylogeny suggested that the genus Dichotomophthora belongs to the order Pleosporales and is closely related to the genus Curvularia [102]. On P. oleracea, D. lutea causes spots that are oval to irregular in shape and pale brown with dark borders [100]. Since 2015, leaf spot outbreaks have been reported in Iran, where this plant is commonly cultivated. The pathogenicity of isolates was confirmed by artificial inoculation trials on 2-month-old purslane plants [102]. Other foliar pathogens occasionally described on P. oleracea, are Phyllosticta portulacae (Botryosphaeriales) [103] and Stemphylium vesicarium [99].
Another member of the genus Dichotomophthora, D. portulacae (Pleosporales), is frequently associated with stem necrosis on P. oleracea. This fungus was first described by Mehrlich and Fitzpatrick [106] in Hawaii (USA) on stems and leaves. The occurrence of this disease was recently recorded in Iraq [107]. A detailed description of symptoms is provided by Klisiewciz et al. [109]. The disease is characterized by dark lesions and constrictions on the main stem and occasionally on lateral branches; from the main stem, lesions can progress and hit the root system, with damaged tissues showing an abundance of sclerotia. The disease is particularly severe in young plants and seedlings. In artificial inoculation trials, the leaves of 4-week-old plants showed necrotic spots within 24 h. The disease progressed to the main stem and branches and eventually resulted in dieback, which was promoted by high air humidity; affected leaves displayed a yellow color and eventually fell off the plants. Klisiewciz et al. [109] found 35 isolates of D. portulacae in California (USA). In addition, Mitchell [108] designated black stem rot as the main symptom of D. portulacae on P. oleracea in Texas (USA). In Turkey, D. portulacae was associated with crown rot and root rot on cultivated purslane [105]. Here, in artificial inoculation trials, damping-off of 3-week-old seedlings was evident within eight days after inoculation.
Curvularia portulacae (Pleosporales) is described as another pathogenic fungus causing stem rot on purslane. The disease was described by Rader [110], who proposed a new species, namely Helminthosporium portulacae that was subsequently accommodated into the genus Curvularia Boedijn [180]. Rader [110] described the presence of water-soaked lesions along the stem which coalesce and cover the whole plant organ; abundance of conidia on stems and leaves was also reported.
Furthermore, three different species of Colletotrichum (Glomerellales) are reported: C. spinaciae [95], C. fructicola, and C. gloeosporioides [94]. Purslane is described as a host of C. fructicola by the European Food Safety Authority as well [181]. Since the Simmonds [94] specimen has not been further studied, there is little chance that is a record of C. gloeosporioides sensu stricto, while it is probably a C. gloeosporioides species complex. In any case, this fungal genus is responsible for anthracnose, a disease characterized by the presence of sunken, dark brown lesions representing fruiting bodies (known as acervuli) bearing conidia. This disease mainly hits leaves and fruits; however, no information about the pathosystems is provided in the examined literature.
Notably, P. oleracea is particularly susceptible to rot agents. Other pathogens causing rots on various tissues occasionally documented on purslane are Berkeleyomyces basicola (Microascales) causing black root rot [96], reported as Thielaviopsis basicola, and accommodated into the genus Berkeleyomyces by Nel et al. [182]; Globisporangium intermedium and G. irregulare (Peronosporales) causing root dieback [104]; Sclerotinia sclerotiorum (Helotiales) causing crown rot [98]; and Rhizoctonia solani (Cantharellales) [97] and Sclerotium rolfsii (Thelephorales) [94], both causing basal stem rot.
Portulaca oleracea is indicated as a host of Verticillium dahliae (Glomerellales) [92]. Inoculated purslane plants displayed mild symptoms such as stunting, reduced plant height and root development, lower pigment content in the stem, and brown discoloration of the tap root. The author stated that no clear Verticillium symptoms, like wilt or chlorosis, were visible. It is known that purslane can represent a source of inoculum for some important pathogens without being damaged. Important pathogens like Phytophthora capsici [183], Globisporangium sylvaticum, and G. ultimum [184] were associated with roots of P. oleracea. In both studies, artificial inoculation trials failed to cause any visible symptoms. These case studies provide important insights into pathogenicity and virulence. Pathogenicity is defined as the ability of a pathogen to cause disease on a particular host and is regarded as a qualitative trait. It mainly depends on the ability of the pathogen to invade and colonize the host. Virulence is the extent of damage caused to the host; thus it is a quantitative property [185]. The above-mentioned studies concerned species that were pathogenic (i.e., able to colonize the host tissues) but caused no or mild symptoms (i.e., no or weakly virulent). In this scenario, pathogenic organisms can successfully establish and survive within a particular host, potentially acting as a means of spreading to other crops.
4.8. Salicornia spp.
In this review, three different species within the genus Salicornia (Amaranthaceae) have been considered: S. perennans (glasswort), S. perennis (perennial glasswort), and S. ramosissima (twiggy glasswort). Despite its success as a halophytic crop, only a few documents were retrieved, accounting for two pathogenic species, belonging to as many orders. Salicornia spp. can be exploited for human consumption as a source of pharmaceutical compounds, for biodiesel production, for the remediation of saline soils, and for the treatment of saline effluents from aquaculture [186,187]. This genus includes branched plants with succulent and articulated stems [7].
Uromyces salicorniae (Pucciniales) is a rust agent affecting all three Salicornia species [121]. On S. perennans, this rust was reported several times [54,77,119,120,121]. In addition, on S. perennans (reported with its synonym S. europaea), a strain of Phytophthora inundata was isolated in the Netherlands from leaves showing dark lesions [118].
4.9. Salsola spp.
The genus Salsola (Amaranthaceae) comprises several halophytic annual herbs, characterized by spinous leaves which thrive in arid and semi-arid regions [188]. S. kali (common saltwort) and S. tragus (Russian thistle) are the two species considered in this review. Nine pathogenic species, residing in five orders (Diaporthales, Erysiphales, Glomerellales, Hypocreales, and Pucciniales), were found. Although considered noxious weeds, both S. kali and S. tragus can be agronomically exploited, especially as forage for cattle and sheep [189]. Most of the studies describing the occurrence of plant pathogens on such species have explored the potential of causal agents to be employed as bioherbicides [130,131,132,190]. For instance, the potentiality of the anthracnose agent Colletotrichum salsolae (Glomerellales) has been widely investigated for the control of S. tragus [130,131]. The pathogen was first described by Schwarczinger et al. [128] in Hungary as C. gloeosporioides, isolated from leaves and stems showing the typical symptoms of anthracnose, i.e., chlorotic spots with acervuli developing on the center of the lesions. Pathogenicity was confirmed through artificial inoculations, which led to the death of inoculated plants within two weeks [128]. The disease was also reported in Greece [129] and Russia [130]. Berner et al. [191] designated the pathogenic strain of Colletrotichum as C. gloeosporioides f. sp. salsolae. Later, the resolution of the C. gloeosporioides species complex led to the establishment of the novel and highly host-specific species C. salsolae [192]. The application of C. salsolae as a biocontrol agent against Russian thistle was tested in open fields in Greece and Russia [131]. The authors monitored the development of the disease over two years and concluded that the treatment of infested fields with C. salsolae could eliminate the weed. In addition, they found a strong relationship between disease incidence and rainfall, suggesting that the pathogen mainly spreads through rain splash and high moisture conditions promote the disease.
The stem canker agent Diaporthe eres (Diaporthales) was also assessed as a mycoherbicide [132]. It was isolated in Russia from dark necrotic lesions at the base of the stem of symptomatic S. tragus plants. In artificial inoculation trials, spraying of conidia suspension onto leaves and stems led to a disease incidence of 100% at 70 days after inoculation [132].
Furthermore, the rust agent Uromyces salsolae is under examination as a potential bioherbicide for the control of S. tragus [190]. Interestingly, the same species is reported as a rust agent on S. kali [126,127].
On S. kali, two additional members of Pucciniales were noted: Uromyces chenopodii [66,98,125] and Puccinia aristidae [124]. This species was also indicated as a host of two powdery mildew agents, Leveillula cylindrospora [123] and L. taurica [64], V. dahliae [92], and Dactylonectria macrodidyma (Hypocreales) [122].
4.10. Soda inermis
Soda inermis (=Syn.: Salsola soda, Amaranthaceae, Monk’s beard) is an annual shrub widely spread in the Mediterranean Region. This crop can tolerate high levels of salinity and displays a marked ability to accumulate sodium from the soil [193,194]. Although this crop is highly consumed and cultivated in commercial greenhouses in Mediterranean countries, there are only a few reports about diseases caused by pathogens. We retrieved two records on this halophyte: Leveillula cylindrospora (Erysiphales) as a powdery mildew agent [76] and Uromyces salsolae (Pucciniales) as a rust agent [133].
4.11. Suaeda spp.
The genus Suaeda (Amaranthaceae) comprises several halophytic species, which can be either annual or perennial. In this study, two species have been considered: S. maritima (sea-blite) and S. vera (shrubby sea-blite) for which we retrieved only one association. Both species have edible leaves; S. maritima is particularly rich in proteins, minerals, and vitamins [195], while S. vera seeds can be employed as a source of edible oil [196]. On both species, the rust Uromyces chenopodii (Pucciniales) is the only fungal pathogen reported in the literature, with several records [66,77,103,120,134,135,136].
5. Host Range of Pathogens of the Mediterranean Halophytes
A plant disease is the result of the interaction between a susceptible host plant, one or more virulent pathogens, and the environment. The plant–pathogen interaction is a complex process involving pathogen- and plant-derived molecules and represents a dynamic situation in which a co-evolutionary arms race takes place. As one side advances, strong selective pressure arises on the other to increase or modify its specific response [197].
Plant–pathogen characteristics evolve and vary both in space and time and are strongly associated with differences in the level of losses caused by the disease [197]. Therefore, host–pathogen interactions are usually the outcome of evolutionistic mechanisms by which the two components of the system evolve synergistically, and this is emphasized in the case of highly specific pathogens. Nonetheless, besides those exclusive to one host species, there are many cases of pathogenic species with a broad range of hosts, both at genus and species levels. Pathogenic species reported on two or more different halophytic plant hosts are summarized in Table 3.
At host genus level, L. taurica is the most heterogeneous species as it is documented on four different host genera, residing in three different families. L. taurica powdery mildew was reported to affect members of the family Amaranthaceae, in the host plants B. vulgaris subsp. maritima and S. kali [64]; Apiaceae, in C. maritimum [64,75,76]; and Brassicaceae, in L. latifolium [64]. Similarly, the wilt pathogen V. dahliae was described on species of three different families, corresponding to as many plant host genera. Occurrences of this pathogen were reported on L. perfoliatum, P. oleracea, and S. kali [92].
The rust P. aristidae is the only member of this group of pathogens that was described on hosts belonging to two different families, corresponding to as many host genera, namely Amaranthaceae, in S. kali [124], and Brassicaceae, in L. perfoliatum [59]. Interestingly, besides two records on Brassicaceae [59,88] (Table 2), all the other members of the order Pucciniales were revealed solely on species in the family Amaranthaceae.
Other pathogens with two or more host species are exclusively described on members of the same family, e.g., A. candida, E. cruciferarum, H. parasitica, L. cylindrospora, U. chenopodii, and U. salsolae, whereas others are limited to one host genus, namely A. caulina, P. lepidii, P. spegazzinii, S. atriplicis, U. coralloides, and U. salicorniae (Table 3). Interestingly, the pathogenic species found on Atriplex spp. are not shared among any other host taken under consideration. This is also the case for Salicornia spp., albeit very few records were retrieved for this halophyte and thus it is more likely to be due to insufficient information.
Finally, it is worth noting that, in this study, many of the listed pathogenic species (e.g., A. alternata, G. irregulare, R. solani, and S. sclerotiorum) are only reported once despite having an extremely broad host range. This is further proof of evidence on how research on this topic should be encouraged to widen the current knowledge about the range of hosts and distribution of these economically important pathogens.
6. How Fungi and Fungal-like Organisms Deal with Osmotic Stress
Like plants, microorganisms inhabiting saline ecosystems must adapt to live in such extreme environments. Osmotic stress response, either hypo- or hyperosmotic, varies among the kingdoms and fungal and fungal-like organisms are not an exception. Salinity can negatively or positively affect plant pathogens.
In the first case, which is the most common, high salt concentrations reduce the production of mycelia and conidial formation because of negative osmotic potential along with toxic and nutritional effects [198]. The growth of wilt fungi, V. dahliae and Verticillium albo-atrum, on potato dextrose agar (PDA) or Dox media declined with the increase in NaCl concentration [199,200]. Bhai et al. [201] also made similar findings in Phytophthora capsici. In Fusarium oxysporum the synthesis of gibberellic acid (GA) and indole acetic acid (IAA) showed a drastic decline when NaCl concentration was higher than 1% [202].
On the other hand, salinity can promote the growth of some plant pathogens, which is reflected in enhanced pathogen activity through increased enzymatic and cellular metabolic activities. There are some reports on the positive effects of salinity on plant pathogens. Cercospora leaf spot was favored by soil salinity [203] and a significant increase in mycelium growth of Fusarium solani by sea salt and NaCl was also observed by Firdous and Shahzad [204]. Chlamydospores of F. oxysporum f.sp. vasinfectum exhibited better growth and greater virulence on cotton in a saline medium [205]. Sclerotium rolfsii produced more xylanase and galactanase with increased salt concentrations [206]. The NaCl concentration required to reduce the growth of Pythium aphanidermatum isolates by 50% ranged from 23 to 62 dS m−1 (≃13.5–36.2 g L−1), with an average of 46 dS m−1 (≃26.9 g L−1) [207]. Russo et al. [208] observed the growth of Globisporangium irregulare even at 32 g L−1 of NaCl, which is almost the salt concentration of seawater. In general, the growth of plant pathogens under NaCl conditions leads to increased virulence and may result in adaptation to salinity over time causing the appearance of halophytic strains.
To the best of our knowledge, only a few studies have examined the physiological response of Oomycota to osmotic stress, but sensing mechanisms are not addressed.
A central role in osmoregulation was attributed to proline as a compatible solute in Phytophthora nicotianae [209]. High concentrations of proline were detected in sporulating hyphae, with the content being much lower in vegetative hyphae and germlings and almost null in zoospores. Incipient zoospores accumulate proline as long as they are in the sporangium lumen (a hyperosmotic environment) and release this solute as soon as they are discharged from the sporangium into a hypoosmotic environment [209]. Moreover, in Phytophthora cinnamomi, the switch from a hypoosmotic condition to a hyperosmotic condition resulted in an increase in proline content in hyphae [210]. Recently, Lacalendola et al. [211] have demonstrated the ability of encysted zoospores of Achlya bisexualis to regulate cell turgor in hyperosmotic conditions by evaluating their biomechanical responses.
On the other hand, the mechanisms of osmotic stress sensing and response are well documented in Fungi. The most detailed mechanisms of sensing are those described in yeasts and major insights are provided by the halosensitive Saccharomyces cerevisiae and the halotolerant black yeast Hortaea werneckii. This heavily melanized yeast thrives in hypersaline habitats, making it a model organism for osmotic stress, particularly salt stress, in fungi [212]. In S. cerevisiae, salt adaptation is mediated by the osmosensor proteins Sho1p and the histidine-kinase Sln1p [213]. In H. werneckii, a central role has been attributed to the homolog putative osmosensor protein HwSho1p [214] and the histidine-kinase (HK)-like osmosensor HwHK7p (S. cerevisiae Sln1p homolog) [215]. After sensing, cells respond to hyperosmotic changes by activating the high osmolarity glycerol (HOG) pathway, a mitogen-activated protein kinase (MAPK) pathway. The activation of the osmosensors triggers a kinase cascade, which consists of three MAPKs (MAPKKK, MAPKK, MAPK) activated by sequential phosphorylations. The last MAPK activated is Hog1p. Thus, once Hog1p is phosphorylated, it migrates into the nucleus where it triggers several genes involved in osmotic stress response [215].
In filamentous fungi, sensing and response to osmotic stress are different. In Aspergillus nidulans, for instance, ShoAp (Sho1p homolog) may not be involved in the activation of HogAp (Hog1p homolog) MAPK and, hence, in osmostress response [216]. This is due to the lack of a Pro-rich domain in the downstream MAPKK, and the activation of the HOG pathway depends on the two-component signaling (TcsBp) histidine kinase [216]. Similar findings were identified in Fusarium graminearum mutants lacking both FgSHO1 and FgSLN1, which did not show greater sensitivity to osmotic stress compared to the wild type [217]. The same authors reported a central role for FgSHO1 in conidiation, virulence, and deoxynivalenol biosynthesis. Similarly, in V. dahliae, VdSHO1 mutants were not sensitive to osmotic stress but lost the ability for host penetration and melanin biosynthesis [218]. Conversely, in Magnaporthe oryzae, the causal agent of rice blast, mutants lacking MoSLN1 showed a greater susceptibility to osmotic and oxidative stress together with a loss of pathogenicity [219]. The central role of Hog1p in osmostress response has been confirmed in several plant pathogenic fungi such as F. graminearum [220], A. alternata [221], V. dahliae [222], and Botrytis cinerea [223].
Regardless of differences in sensing and signaling, physiological and biochemical responses to osmostress are shared to a certain extent among the fungal kingdom. The main mechanisms include ion homeostasis, biosynthesis of compatible solutes, and modifications of plasma membrane fluidity. Turgor adjustment requires the maintenance of balanced ion concentration, which is achieved primarily through active transport. In addition, Na+ ions are toxic to cells, even at low concentrations. The extremely halotolerant H. werneckii adopts a “Na+ exclusion” strategy as it can maintain very low intracellular levels of both Na+ and K+ ions [224]. This strategy is accomplished by active transport of Na+ outside the cell, mainly through Na+/H+ antiporter and Na+-exporting P-type ATPases [225,226]. Cell turgor is also maintained through the biosynthesis of compatible solutes, in particular, glycerol. In S. cerevisiae, glycerol biosynthetic gene GPD1 is the main target of phosphorylated Hog1p [214]. In H. werneckii, two homologs of GPD1 were identified, HwGPD1A and HwGPD1B, which are both induced by salt stress [227]. Glycerol is mainly accumulated in the exponential phase of growth, together with erythritol, arabitol, and mannitol, which instead are synthesized principally during the stationary phase [228]. The crucial role of glycerol in osmotic stress response is supported by its integration in all the above-mentioned mechanisms. For instance, S. cerevisiae can uptake glycerol through the H+/glycerol symporter Stl1p [229]. Genes encoding for this symporter were also found in M. oryzae [230]. Furthermore, due to its chemical properties, glycerol is easily leaked from the plasma membrane, forcing cells to adopt strategies to keep proper fluidity while avoiding excessive leaking. To do this, H. werneckii embeds melanin into the cell wall, increasing its ability to retain this compatible solute [231]. The main modifications in plasma membrane composition are a higher unsaturation level of the lipidic bilayer and the promotion of phosphatidylcholine and phosphatidylethanolamine biosynthesis [232]. Contrary to what was found in H. werneckii, salt-exposed S. cerevisiae cells increase the sterol-to-lipid ratio in the membrane [233].
7. Conclusions
In the examined bibliography on major Mediterranean halophytes, only a few studies have investigated host–pathogen associations in cultivated plants (apart from artificial inoculation trials), and, in most documents, host plants were collected in the wild, as part of checklists, or they represented hosts for type specimens of pathogenic species. Most of the pathogenic species have been isolated from above-ground organs, especially leaves. In particular, in genera Atriplex, Beta, Cakile, Crithmum, Salicornia, Soda, and Suaeda, only foliar diseases were recorded.
It is probable that several other pathogens have been neglected, as plants can be exposed to different pathogens when they grow in the wild or are cultivated. Apart from Salicornia spp. and S. inermis, the consumption of other halophytic species is mainly linked to ethnobotanical traditions, and this explains the limited bibliography on the topic. However, if we intend to turn these species into market-oriented crops, to eliminate the risks of foraging, the responsibility of production must lie with experienced growers. Hence, growing protocols must be implemented, including pest and disease management. This aspect is highlighted by those cases in which highly specific pathogens have been isolated (for example, the pathosystems Peronospora spp./Atriplex spp. and A. candida/Lepidium spp.), which are not related to major crop pathogens. They are genus- or even species-specific pathogens that can represent a reservoir of new diseases to which greater attention should be paid. In the optic of agricultural exploitation of these species, it is fundamental to assess the occurrence of pathogens to develop accurate control strategies. On the other hand, halophytes can be secondary hosts for pathogenic species and represent a significant source of inoculum for important pathogens of important crops. Moreover, some species can promote the spread of pathogens through the dissemination of infected organs (e.g., infected seeds as in Alternaria sp./C. maritima). In this regard, the pathosystem A. alternata/C. maritima is of pivotal importance, as it is the only host/pathogen binomial investigated in salinity conditions. In the study, the presence of salt was associated with reduced susceptibility of the host toward the pathogen. The role of salt in disease development is a key aspect that needs to be deepened as this abiotic factor can affect the virulence and the production of secondary metabolites in plant pathogenic species. Therefore, boosting research on plant diseases of halophytic plants is crucial to identifying novel diseases and better understanding the spread of pathogenic species.
Finally, if halophytic plants are intended for consumption, it is essential to evaluate the risks associated with the presence of mycotoxigenic species.
Author Contributions
Conceptualization, E.D.C., A.P. and S.P.; writing—original draft, E.D.C.; review and editing, A.P. and S.P.; supervision, A.P. and S.P.; funding acquisition, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was conducted in the framework of the project entitled “HALOphytes grown in saline Water for the production of INnovative ready-to- eat salad-HALOWIN” and funded by the University of Pisa, “PRA 2020”, grant number PRA_2020_43.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Dataset available on request from the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lopez-Alvarado, J.; Farris, E. Ecology and Evolution of Plants in the Mediterranean Basin: Perspectives and Challenges. Plants 2022, 11, 1584. [Google Scholar] [CrossRef] [PubMed]
- Gualdi, S.; Somot, S.; Li, L.; Artale, V.; Adani, M.; Bellucci, A.; Braun, A.; Calmanti, S.; Carillo, A.; Dell’Aquila, A.; et al. The CIRCE Simulations: Regional Climate Change Projections with Realistic Representation of the Mediterranean Sea. Bull. Am. Meteorol. Soc. 2013, 94, 65–81. [Google Scholar] [CrossRef]
- Luterbacher, J.; Xoplaki, E.; Casty, C.; Wanner, H.; Pauling, A.; Küttel, M.; Rutishauser, T.; Brönnimann, S.; Fischer, E.; Fleitmann, D.; et al. Chapter 1 Mediterranean Climate Variability over the Last Centuries: A Review. In Developments in Earth and Environmental Sciences; Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Eds.; Mediterranean; Elsevier: Amsterdam, The Netherlands, 2006; Volume 4, pp. 27–148. [Google Scholar]
- Rizzo, A.; Vandelli, V.; Gauci, C.; Buhagiar, G.; Micallef, A.S.; Soldati, M. Potential Sea Level Rise Inundation in the Mediterranean: From Susceptibility Assessment to Risk Scenarios for Policy Action. Water 2022, 14, 416. [Google Scholar] [CrossRef]
- Ferreira, C.S.S.; Seifollahi-Aghmiuni, S.; Destouni, G.; Ghajarnia, N.; Kalantari, Z. Soil Degradation in the European Mediterranean Region: Processes, Status and Consequences. Sci. Total Environ. 2022, 805, 150106. [Google Scholar] [CrossRef] [PubMed]
- Stolte, J.; Tesfai, M.; Øygarden, L.; Kværnø, S.; Keizer, J.; Verheijen, F.; Panagos, P.; Ballabio, C.; Rudi Hessel, R. Soil Threats in Europe; EUR 27607 EN; EU Publications: Luxembourg, 2015. [Google Scholar] [CrossRef]
- Lombardi, T.; Bertacchi, A.; Pistelli, L.; Pardossi, A.; Pecchia, S.; Toffanin, A.; Sanmartin, C. Biological and Agronomic Traits of the Main Halophytes Widespread in the Mediterranean Region as Potential New Vegetable Crops. Horticulturae 2022, 8, 195. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Sui, N. Mechanisms of Salt Tolerance in Halophytes: Current Understanding and Recent Advances. Open Life Sci. 2018, 13, 149–154. [Google Scholar] [CrossRef]
- Feng, Z.T.; Deng, Y.Q.; Liang, X.; Yuan, F.; Hao, J.L.; Zhang, J.C.; Sun, F.S.; Wang, B.S. K+ accumulation in the cytoplasm and nucleus of the salt gland cells of Limonium bicolor accompanies increased rates of salt secretion under NaCl treatment using NanoSIMS. Plant Sci. 2015, 238, 286–296. [Google Scholar] [CrossRef]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Shiri, M.; Rabhi, M.; El Amrani, A.; Abdelly, C. Cross-tolerance to Abiotic Stresses in Halophytes: Application for Phytoremediation of Organic Pollutants. Acta Physiol. Plant. 2015, 37, 209. [Google Scholar] [CrossRef]
- Duarte, B.; Caçador, I. Iberian Halophytes as Agroecological Solutions for Degraded Lands and Biosaline Agriculture. Sustainability 2021, 13, 1005. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible Halophytes of the Mediterranean Basin: Potential Candidates for Novel Food Products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef]
- Lombardi, T.; Ventura, I.; Bertacchi, A. Floristic Inventory of Ethnobotanically Important Halophytes of North-Western Mediterranean Coastal Brackish Areas, Tuscany, Italy. Agronomy 2023, 13, 615. [Google Scholar] [CrossRef]
- Strinati, M. Salicornia, the Resilient Sea Asparagus. 2024. Available online: https://www.greatitalianfoodtrade.it/en/markets/salicornia-lasparagus-resilient-sea-fish/ (accessed on 14 March 2024).
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011; ISBN 978-92-79-20199-8. [Google Scholar] [CrossRef]
- Papanastasis, V.P.; Yiakoulaki, M.D.; Decandia, M.; Dini-Papanastasi, O. Integrating Woody Species into Livestock Feeding in the Mediterranean Areas of Europe. Anim. Feed. Sci. Technol. 2008, 140, 1–17. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal Halophytes: Potent Source of Health Promoting Biomolecules with Medical, Nutraceutical and Food Applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Qasim, M.; Abideen, Z.; Adnan, M.Y.; Gulzar, S.; Gul, B.; Rasheed, M.; Khan, M.A. Antioxidant Properties, Phenolic Composition, Bioactive Compounds and Nutritive Value of Medicinal Halophytes Commonly Used as Herbal Teas. South Afr. J. Bot. 2017, 110, 240–250. [Google Scholar] [CrossRef]
- Stanković, M.S.; Petrović, M.; Godjevac, D.; Stevanović, Z.D. Screening Inland Halophytes from the Central Balkan for Their Antioxidant Activity in Relation to Total Phenolic Compounds and Flavonoids: Are There Any Prospective Medicinal Plants? J. Arid. Environ. 2015, 120, 26–32. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Quah, E.P.L. Antioxidant Properties of Different Cultivars of Portulaca oleracea. Food Chem. 2007, 103, 734–740. [Google Scholar] [CrossRef]
- Boestfleisch, C.; Wagenseil, N.B.; Buhmann, A.K.; Seal, C.E.; Wade, E.M.; Muscolo, A.; Papenbrock, J. Manipulating the Antioxidant Capacity of Halophytes to Increase Their Cultural and Economic Value through Saline Cultivation. AoB Plants 2014, 6, plu046. [Google Scholar] [CrossRef]
- Lopes, M.; Castilho, M.d.C.; Sanches-Silva, A.; Freitas, A.; Barbosa, J.; Gonçalves, M.J.; Cavaleiro, C.; Ramos, F. Evaluation of the Mycotoxins Content of Salicornia spp.: A Gourmet Plant Alternative to Salt. Food Addit. Contam. Part B 2020, 13, 162–170. [Google Scholar] [CrossRef]
- Gunning, D. Cultivating Salicornia Europaea (Marsh Samphire); BIM: Laoghaire, Ireland, 2016. [Google Scholar]
- Accogli, R.; Tomaselli, V.; Direnzo, P.; Perrino, E.V.; Albanese, G.; Urbano, M.; Laghetti, G. Edible Halophytes and Halo-Tolerant Species in Apulia Region (Southeastern Italy): Biogeography, Traditional Food Use and Potential Sustainable Crops. Plants 2023, 12, 549. [Google Scholar] [CrossRef]
- Kraouia, M.; Nartea, A.; Maoloni, A.; Osimani, A.; Garofalo, C.; Fanesi, B.; Ismaiel, L.; Aquilanti, L.; Pacetti, D. Sea Fennel (Crithmum maritimum L.) as an Emerging Crop for the Manufacturing of Innovative Foods and Nutraceuticals. Molecules 2023, 28, 4741. [Google Scholar] [CrossRef]
- Custódio, M.; Lillebø, A.I.; Calado, R.; Villasante, S. Halophytes as Novel Marine Products—A Consumers’ Perspective in Portugal and Policy Implications. Mar. Policy 2021, 133, 104731. [Google Scholar] [CrossRef]
- Maciel, E.; Domingues, P.; Domingues, M.R.M.; Calado, R.; Lillebø, A. Halophyte Plants from Sustainable Marine Aquaponics Are a Valuable Source of Omega-3 Polar Lipids. Food Chem. 2020, 320, 126560. [Google Scholar] [CrossRef] [PubMed]
- Marques, B.; Maciel, E.; Domingues, M.R.; Calado, R.; Lillebø, A.I. Halophyte Plants Cultured in Aquaponics Hold the Same Potential for Valorization as Wild Conspecifics from Donor Sites. Appl. Sci. 2021, 11, 11586. [Google Scholar] [CrossRef]
- Oliveira, V.; Martins, P.; Marques, B.; Cleary, D.F.R.; Lillebø, A.I.; Calado, R. Aquaponics Using a Fish Farm Effluent Shifts Bacterial Communities Profile in Halophytes Rhizosphere and Endosphere. Sci. Rep. 2020, 10, 10023. [Google Scholar] [CrossRef] [PubMed]
- Spradlin, A.; Saha, S. Saline Aquaponics: A Review of Challenges, Opportunities, Components, and System Design. Aquaculture 2022, 555, 738173. [Google Scholar] [CrossRef]
- Stouvenakers, G.; Dapprich, P.; Massart, S.; Jijakli, M.H. Plant Pathogens and Control Strategies in Aquaponics. In Aquaponics Food Production Systems: Combined Aquaculture and Hydroponic Production Technologies for the Future; Goddek, S., Joyce, A., Kotzen, B., Burnell, G.M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 353–378. ISBN 978-3-030-15943-6. [Google Scholar]
- Calabon, M.S.; Jones, E.B.G.; Promputtha, I.; Hyde, K.D. Fungal Biodiversity in Salt Marsh Ecosystems. J. Fungi 2021, 7, 648. [Google Scholar] [CrossRef] [PubMed]
- Farr and Rossman Fungal Databases—Fungus-Host Distribution. Available online: https://fungi.ars.usda.gov/ (accessed on 14 January 2024).
- GBIF: The Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 9 January 2024).
- eHALOPH Halophyte Database v4.65. Available online: https://ehaloph.uc.pt/ (accessed on 9 January 2024).
- Index Fungorum Homepage. Available online: www.indexfungorum.org (accessed on 22 January 2024).
- Kirk, P.; Cannon, P.; Stalpers, J.; Minter, D.W. Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Maciá-Vicente, J.G.; Ferraro, V.; Burruano, S.; Lopez-Llorca, L.V. Fungal Assemblages Associated with Roots of Halophytic and Non-Halophytic Plant Species Vary Differentially Along a Salinity Gradient. Microb. Ecol. 2012, 64, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Maciá-Vicente, J.G.; Jansson, H.-B.; Abdullah, S.K.; Descals, E.; Salinas, J.; Lopez-Llorca, L.V. Fungal Root Endophytes from Natural Vegetation in Mediterranean Environments with Special Reference to Fusarium spp. FEMS Microbiol. Ecol. 2008, 64, 90–105. [Google Scholar] [CrossRef]
- González-Menéndez, V.; Crespo, G.; de Pedro, N.; Diaz, C.; Martín, J.; Serrano, R.; Mackenzie, T.A.; Justicia, C.; González-Tejero, M.R.; Casares, M.; et al. Fungal Endophytes from Arid Areas of Andalusia: High Potential Sources for Antifungal and Antitumoral Agents. Sci. Rep. 2018, 8, 9729. [Google Scholar] [CrossRef]
- El-Morsy, E.-M. Fungi Isolated from the Endorhizosphere of Halophytic Plants from the Red Sea Coast of Egypt. Fungal Divers. 2000, 5, 43–54. [Google Scholar]
- Constaninescu, O. An Annotated List of Peronospora Names: Thunbergia 15. Mycologia 1992, 84, 953. [Google Scholar] [CrossRef]
- van der Aa, H.; van Kesteren, H.A. Some Pycnidial Fungi Occurring on Atriplex and Chenopodium. Persoonia Mol. Phylogeny Evol. Fungi 1979, 10, 267–276. [Google Scholar]
- Conners, I. An Annotated Index of Plant Diseases in Canada and Fungi Recorded on Plants in Alaska, Canada and Greenland. Aspen Bibliogr. 1967, 60, 222. [Google Scholar]
- Andrianova, T.V.; Minter, D.W. Stagonospora atriplicis. [Descriptions of Fungi and Bacteria]. Descr. Fungi Bact. 2005, Sheet 1630. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Klosterman, S.J.; Kummer, V.; Voglmayr, H.; Shin, H.-D.; Thines, M. Multi-Locus Tree and Species Tree Approaches toward Resolving a Complex Clade of Downy Mildews (Straminipila, Oomycota), Including Pathogens of Beet and Spinach. Mol. Phylogenetics Evol. 2015, 86, 24–34. [Google Scholar] [CrossRef]
- Queiroz, J.; Santos, M.A.; Santos, A.M.J.; Michereff, S.; Freire, M. First Report of Diaporthe inconspicua Associated with Shoot Blight of Atriplex nummularia in Brazil. J. Plant Pathol. 2017, 99, 542. [Google Scholar] [CrossRef]
- Blagoveshchenskaya, E.Y. Peronosporales of Skadovsky Zvenigorod Biological Station (Moscow Region). Nov. Sist. Nizshikh Rastenii 2018, 52, 91–100. [Google Scholar] [CrossRef]
- Chater, A.O.; Woods, R.; Stringer, R.; Evans, D.; Smith, P. Downy Mildews (Peronosporaceae) and White Blister-Rusts (Albuginaceae) of Wales; A.O. Chater: Aberystwyth, UK, 2020. [Google Scholar]
- Müller, J.; Kokeš, P. Erweitertes Verzeichnis Der Falschen Mehltaupilze Mährens Und Tschechisch Schlesiens. Czech Mycol. 2008, 60, 91–104. [Google Scholar] [CrossRef]
- Francis, S.M.; Waterhouse, G.M. List of Peronosporaceae Reported from the British Isles. Trans. Br. Mycol. Soc. 1988, 91, 1–62. [Google Scholar] [CrossRef]
- Gustavsson, A. The Genus “Peronospora” in the Iberian Peninsula, Especially in Northern Spain and Andorra. An. Jardín Botánico Madr. 1991, 49, 3–38. [Google Scholar]
- Mułenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A Preliminary Checklist of Micromycetes in Poland = Wstępna Lista Grzybów Mikroskopijnych Polski; Biodiversity of Poland; W. Szafer Institute of Botany Polish Academy of Sciences: Kraków, Poland, 2008; ISBN 978-83-89648-75-4. [Google Scholar]
- Pollack, F.G. An Annotated Compilation of Cercospora Names. Mycol. Mem. 1987, 12, 1–212. [Google Scholar]
- Adamska, I. Microscopic Fungus-like Organisms and Fungi of the Słowiński National Park. II. (NW PoIand). Acta Mycol. 2001, 36, 31–65. [Google Scholar] [CrossRef]
- Braun, U. Miscellaneous Notes on Phytopathogenic Hyphomycetes (II). Mycotaxon 1995, 55, 223–241. [Google Scholar]
- McKenzie, E.H.C. New Plant Disease Records in New Zealand: Miscellaneous Fungal Pathogens II. N. Z. J. Crop Hortic. Sci. 1990, 18, 65–73. [Google Scholar] [CrossRef]
- Shaw, C. Host Fungus Index for the Pacific Northwest-1. Hosts. Wash. State Univ. Agric. Exp. Stn. Bull. 1973, 765, 1–121. [Google Scholar]
- Priest, M.J. Fungi of Australia: Septoria; Australian Biological Resources Study/CSIRO Publishing: Clayton, Australia, 2006. [Google Scholar]
- Fraser, W.P. Cultures of Some Heteroecious Rusts. Mycologia 1911, 3, 67–74. [Google Scholar] [CrossRef]
- Piątek, M.; Wołczańska, A. Some Phytopathogenic Fungi Rare or New to Poland. Pol. Bot. J. 2004, 49, 67–72. [Google Scholar]
- Pirnia, M.; Zare, R.; Zamanizadeh, H.R.; Khodaparast, A. New Records of Cercosporoid Hyphomycetes from Iran. Mycotaxon 2012, 120, 157–169. [Google Scholar] [CrossRef]
- Amano, K. Host Range and Geographical Distribution of the Powdery Mildew Fungi., 2nd ed.; Japan Scientific Societies Press: Tokyo, Japan, 1986; ISBN 978-4-7622-9486-0. [Google Scholar]
- Gjærum, H.B. Rust Fungi from Madeira. Bol. Mus. Munic. Funchal 1982, 34, 5–22. [Google Scholar]
- Gjærum, H.B.; Sunding, P. Flora of Macaronesia. Checklist of rust fungi (Uredinales). Sommerfeltia 1986, 4, 1–46. [Google Scholar] [CrossRef]
- Oliver, E.J.; Thrall, P.H.; Burdon, J.J.; Ash, J.E. Vertical Disease Transmission in the Cakile-Alternaria Host-Pathogen Interaction. Aust. J. Bot. 2001, 49, 561–569. [Google Scholar] [CrossRef]
- Chalbi, A.; Sghaier-Hammami, B.; Meca, G.; Quiles, J.M.; Abdelly, C.; Marangi, C.; Logrieco, A.F.; Moretti, A.; Masiello, M. Characterization of Mycotoxigenic Alternaria Species Isolated from the Tunisian Halophyte Cakile maritima. Phytopathol. Mediterr. 2020, 59, 107–108. [Google Scholar] [CrossRef]
- Chalbi, A.; Sghaier-Hammami, B.; Baazaoui, N.; Hammami, S.B.M.; Ben-Jouira, H.; García-Caparrós, P.; Djébali, N.; Regaya, I.; Debez, A.; Jorrín-Novo, J.V.; et al. Comparative Study of the Effect of Salt Stress, Alternaria alternata Attack or Combined Stress on the Cakile maritima Growth and Physiological Performance. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12446. [Google Scholar] [CrossRef]
- Thrall, P.H.; Burdon, J.J.; Bock, C.H. Short-Term Epidemic Dynamics in the Cakile maritima–Alternaria brassicicola Host–Pathogen Association. J. Ecol. 2001, 89, 723–735. [Google Scholar] [CrossRef]
- Baka, Z.A.M. A Fine-structural Study of the Downy Mildew Fungus Peronospora parasitica in the New Host Cakile maritima. Arch. Phytopathol. Plant Prot. 1995, 30, 41–51. [Google Scholar] [CrossRef]
- Mckenzie, E.H.C.; Dingley, J.M. New Plant Disease Records in New Zealand: Miscellaneous Fungal Pathogens III. N. Z. J. Bot. 1996, 34, 263–272. [Google Scholar] [CrossRef]
- Aldesuquy, H.S.; Baka, Z.A.M. Physiological and Biochemical Changes in Host Leaf Tissues Associated with the Growth of Two Biotrophic Fungi Growing in Egypt. Phyton 1991, 32, 129–142. [Google Scholar]
- García-Blázquez, G.; Constantinescu, O.; Telleria, M.; Martín, M. Preliminary Check List of Albuginales and Peronosporales (Chromista) Reported from the Iberian Peninsula and Balearic Islands. Mycotaxon 2006, 98, 185–188. [Google Scholar]
- Voytyuk, S.; Heluta, V.; Wasser, S.; Nevo, E.; Takamatsu, S. Biodiversity of the Powdery Mildew Fungi (Erysiphales, Ascomycota) of Israel (Biodiversity of Cyanoprocaryotes, Algae and Fungi of Israel); Ganter Verlag: Ruggell, Liechtenstein, 2009; ISBN 978-3-906166-74-2. [Google Scholar]
- Braun, U. The Powdery Mildews (Erysiphales) of Europe; Gustav Fischer Verlag: Jena, Germany, 1995; ISBN 978-3-334-60994-1. [Google Scholar]
- Pantidou, M.E. Fungus-Host Index for Greece; Benaki Phytopathological Institute: Kiphissia, Athens, 1973. [Google Scholar]
- Braun, U. A Monograph of Cercosporella, Ramularia and Allied Genera (Phytopathogenic Hyphomycetes); IHW-Verlag: Eching, Germany, 1995; Volume 1, ISBN 978-3-930167-11-1. [Google Scholar]
- Savchenko, K.G.; Carris, L.M.; Castlebury, L.A.; Heluta, V.P.; Wasser, S.P.; Nevo, E. Revision of Entyloma (Entylomatales, Exobasidiomycetes) on Eryngium. Mycologia 2014, 106, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.B.; Ellis, J.P. Microfungi on Land Plants: An Identification Handbook; Richmond Publishing: London, UK, 1997. [Google Scholar]
- Unamuno, L.M. Enumeración y distribución geográfica de los Ascomicetos de la Península Ibérica y de las Islas Baleares. Mem. Real. Acad. Ci Exact. Madr. Ser. Ci Nat. 1941, 8, 1–403. [Google Scholar]
- Fotouhifar, K.-B.; Hedjaroude, G.-A.; Leuchtmann, A. ITS rDNA Phylogeny of Iranian Strains of Cytospora and Associated Teleomorphs. Mycologia 2010, 102, 1369–1382. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Thines, M.; Choi, I.-Y.; Shin, H.-D. Perofascia Is Not Monotypic: The Description of the Second Taxon Affecting the South American Crop Maca (Lepidium meyenii). Mycol. Prog. 2017, 16, 857–864. [Google Scholar] [CrossRef]
- Gannibal, P.; Gasich, E.L. Causal Agents of the Alternariosis of Cruciferous Plants in Russia: Species Composition, Geography and Ecology. Mikol. I Fitopatol. 2009, 43, 447–456. [Google Scholar]
- Bakhshi, M.; Arzanlou, M.; Babai-ahari, A.; Groenewald, J.Z.; Crous, P.W. Novel Primers Improve Species Delimitation in Cercospora. IMA Fungus 2018, 9, 299–332. [Google Scholar] [CrossRef]
- Gautam, A.; Avasthi, S. Diversity of Powdery Mildew Fungi from North Western Himalayan Region of Himachal Pradesh—A Checklist. Plant Pathol. Quar. 2018, 8, 78–99. [Google Scholar] [CrossRef]
- Kabaktepe, S.; Heluta, V.; Akata, I. Checklist of Powdery Mildews (Erysiphales) in Turkey. Biol. Divers. Conserv. 2015, 8, 128–146. [Google Scholar]
- Ramazanova, S.S.; Faizieva, F.K.; Sagdullaeva, M.S.; Kirgizbaeva, K.M.; Gaponenko, N.I. Fungal Flora of Uzbekistan, Rustfungi. Fan Publ. Task. 1986, 3, 1–229. [Google Scholar]
- Koike, S.T.; Sullivan, M.J.; Southwick, C.; Feng, C.; Correll, J.C. Characterization of White Rust of Perennial Pepperweed Caused by Albugo candida in California. Plant Dis. 2011, 95, 876. [Google Scholar] [CrossRef] [PubMed]
- Rector, B.G.; Wang, S.; Choi, Y.-J.; Thines, M. First Report of Albugo lepidii Causing White Rust on Broadleaved Pepperweed (Lepidium latifolium) in Nevada and California. Plant Dis. 2016, 100, 229. [Google Scholar] [CrossRef]
- Scholtz, H.; Scholtz, I. Die Brandpilze Deutschlands (Ustilaginales); Englera: La Jolla, CA, USA, 1988; Volume 8. [Google Scholar]
- Woolliams, G.E. Host Range and Symptomatology of Verticillium dahliae in Economic, Weed, and Native Plants in Interior British Columbia. Can. J. Plant Sci. 1966, 46, 661–669. [Google Scholar] [CrossRef]
- Ruszkiewicz-Michalska, M.; Michalski, M. Phytopathogenic Micromycetes in Central Poland. I. Peronosporales and Erysiphales. Acta Mycol. 2005, 40, 223–250. [Google Scholar] [CrossRef]
- Simmonds, J.H. Host Index of Plant Diseases in Queensland; Queensland Department of Primary Industries: Brisbane, Australia, 1966.
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum Species with Curved Conidia from Herbaceous Hosts. Fungal Divers. 2009, 39, 45–87. [Google Scholar]
- Gayed, S.K. Host Range and Persistence of Thielaviopsis basicola in Tobacco Soil. Can. J. Plant Sci. 1972, 52, 869–873. [Google Scholar] [CrossRef]
- Alfieri, S.A., Jr.; Lanngdon, K.R.; Wehlburg, C.; Kimbrough, J.W. Index of Plant Diseases in Florida; Bulletin 11; Florida Department of Agriculture and Consumer Services, Division of Plant Industry: Gainesville, FL, USA, 1984.
- Shivas, R.G. Fungal and Bacterial Diseases of Plants in Western Australia. J. R Soc. West. Aust. 1989, 72, 1–62. [Google Scholar]
- Mcdonald, M.R.; Stricker, S.; Gossen, B.D. Weed Hosts and Winter Survival of Stemphylium vesicarium on Onion in Ontario, Canada. Can. J. Plant Pathol. 2023, 45, 134–139. [Google Scholar] [CrossRef]
- Heidari, K.; Farokhinejad, R.; Mehrabi-Koushki, M. Occurrence of Purslane Leaf Spot Caused by Dichotomophthora lutea in Iran. Australas. Plant Dis. Notes 2018, 13, 33. [Google Scholar] [CrossRef]
- de Hoog, G.S.; van Oorschot, C.A.N. Taxonomy of the Dactylaria Complex I: Notes on the Genus Dichotomophthora. Proc. Van De K. Ned. Akad. Van Wet. Sect. C 1983, 86, 55–61. [Google Scholar]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F.; et al. Genera of Phytopathogenic Fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef]
- Venturella, G. A checklist of Sicilian Fungi. Bocconea 1991, 2, 5–221. [Google Scholar]
- Kalu, N.N.; Sutton, J.C.; Vaartaja, O. Pythium spp. Associated with Root Dieback of Carrot in Ontario. Can. J. Plant Sci. 1976, 56, 555–561. [Google Scholar] [CrossRef]
- Eken, C. Dichotomophthora portulacae on Portulaca oleracea in Turkey. Mycotaxon 2003, 87, 153–156. [Google Scholar]
- Mehrlich, F.P.; Fitzpatrick, H.M. Dichotomophthora portulacae, A Pathogen of Portulaca oleracea. Mycologia 1935, 27, 543–550. [Google Scholar] [CrossRef]
- Abdulla, Z.K. Dichotomophthora portulacae, a New Record from Iraq. Mycotaxon 2021, 136, 215–218. [Google Scholar] [CrossRef]
- Mitchell, J.K. Dichotomophthora portulacae Causing Black Stem Rot on Common Purslane in Texas. Plant Dis. 1986, 70, 603b. [Google Scholar] [CrossRef]
- Klisiewicz, J.M. Black Stem: A Fungal Disease of Common Purslane in California. Plant Dis. 1983, 67, 1162. [Google Scholar] [CrossRef]
- Rader, W.E. Helminthosporium portulacae a New Pathogen of Portulaca oleracea L. Mycologia 1948, 40, 342–346. [Google Scholar] [CrossRef]
- Savile, D.B.O.; Parmelee, J.A. Some Fungal Parasites of Portulacaceae. Mycologia 1956, 48, 573–590. [Google Scholar] [CrossRef]
- El-Buni, A.M.; Rattan, S.S. Check List of Libyan Fungi; Flora of Libya; Al Faateh University Faculty of Science Department of Botany: Tripoli, Libya, 1981. [Google Scholar]
- Riethmüller, A.; Voglmayr, H.; Göker, M.; Weiß, M.; Oberwinkler, F. Phylogenetic Relationships of the Downy Mildews (Peronosporales) and Related Groups Based on Nuclear Large Subunit Ribosomal DNA Sequences. Mycologia 2002, 94, 834–849. [Google Scholar] [CrossRef]
- Vrandecic, K.; Jerković, D.; Cosic, J.; Poštić, J.; Bijelić, Z. White Blister Species (Albuginaceae) on Weeds. Poljoprivreda 2011, 17, 47–51. [Google Scholar]
- Mahmoudi, E.; Ahmadi, A. White Rust Disease of Portulaca oleracea Caused by Wilsoniana portulacae, First Report in Isfahan. In Proceedings of the 1st International Conference on New Ideas in Agriculture, Islamic Azad University Khorasgan Branch, Isfahan, Iran, 26–27 January 2014. [Google Scholar]
- Heller, A.; Thines, M. Evidence for the Importance of Enzymatic Digestion of Epidermal Walls during Subepidermal Sporulation and Pustule Opening in White Blister Rusts (Albuginaceae). Mycol. Res. 2009, 113, 657–667. [Google Scholar] [CrossRef]
- Tóth, T.; Kövics, G. White Rust Species (Chromista, Peronosporomycetes, Albuginales, Albuginaceae) on Common Weeds in Hungary. Acta Agrar. Debreceniensis 2015, 66, 30–33. [Google Scholar] [CrossRef]
- Man in ’t Veld, W.A.; Rosendahl, K.C.H.M.; van Rijswick, P.C.J.; Meffert, J.P.; Boer, E.; Westenberg, M.; van der Heide, T.; Govers, L.L. Multiple Halophytophthora spp. and Phytophthora spp. Including P. gemini, P. inundata and P. chesapeakensis sp. nov. Isolated from the Seagrass Zostera marina in the Northern Hemisphere. Eur. J. Plant Pathol. 2019, 153, 341–357. [Google Scholar] [CrossRef]
- Braun, U. Die Rostpilze (Uredinales) der Deutschen Demokratischen Republik. Feddes Repert. 1982, 93, 213–333. [Google Scholar] [CrossRef]
- Henderson, D.M. A Checklist of the Rust Fungi of the British Isles; British Mycological Society: Manchester, UK, 2000; ISBN 978-0-9527704-4-2. [Google Scholar]
- Woods, R.; Stringer, R.; Evans, D.; Chater, A. Rust Fungus Red Data List and Census Catalogue for Wales; A.O. Chater: Aberystwyth, UK, 2015. [Google Scholar]
- Agustí-Brisach, C.; Gramaje, D.; León, M.; García-Jiménez, J.; Armengol, J. Evaluation of Vineyard Weeds as Potential Hosts of Black-Foot and Petri Disease Pathogens. Plant Dis. 2011, 95, 803–810. [Google Scholar] [CrossRef]
- Khodaparast, S.A.; Takamatsu, S.; Hedjaroude, G.-A. Phylogenetic Structure of the Genus Leveillula (Erysiphales: Erysiphaceae) Inferred from the Nucleotide Sequences of the rDNA ITS Region with Special Reference to the L. taurica Species Complex. Mycol. Res. 2001, 105, 909–918. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; McHenry, J. Check List and Host Index for Arizona Rust Fungi. Bull. Univ. Ariz. Agric. Exp. Stn. 1969, 186, 1–40. [Google Scholar]
- Jørstad, I. Uredinales of the Canary Islands. Skr. Utg. Av. Det. Nor. Vidensk. Akad. I Oslo. 1958, 2, 1–182. [Google Scholar]
- Hasan, S.; Sobhian, R.; Hérard, F. Biology, Impact and Preliminary Host-Specificity Testing of the Rust Fungus, Uromyces salsolae, a Potential Biological Control Agent for Salsola kali in the USA. Biocontrol Sci. Technol. 2001, 11, 677–689. [Google Scholar] [CrossRef]
- Richiteanu, A. Uredinales from North Algeria. Rev. Roum. Biol. 1960, 35, 33–44. [Google Scholar]
- Schwarczinger, I.; Vajna, L.; Bruckart, W.L. First Report of Colletotrichum gloeosporioides on Russian-Thistle. Plant Dis. 1998, 82, 1405. [Google Scholar] [CrossRef] [PubMed]
- Berner, D.K.; Cavin, C.A.; McMahon, M.B.; Loumbourdis, I. First Report of Anthracnose of Salsola tragus Caused by Colletotrichum gloeosporioides in Greece. Plant Dis. 2006, 90, 971. [Google Scholar] [CrossRef] [PubMed]
- Kolomiets, T.; Skatenok, O.; Alexandrova, A.; Mukhina, Z.; Matveeva, T.; Bogomaz, D.; Berner, D.K.; Cavin, C.A. First Report of Anthracnose of Salsola tragus Caused by Colletotrichum gloeosporioides in Russia. Plant Dis. 2008, 92, 1366. [Google Scholar] [CrossRef] [PubMed]
- Berner, D.; Lagopodi, A.L.; Kashefi, J.; Mukhina, Z.; Kolomiets, T.; Pankratova, L.; Kassanelly, D.; Cavin, C.; Smallwood, E. Field Assessment, in Greece and Russia, of the Facultative Saprophytic Fungus, Colletotrichum salsolae, for Biological Control of Russian Thistle (Salsola tragus). Biol. Control 2014, 76, 114–123. [Google Scholar] [CrossRef]
- Kolomiets, T.; Mukhina, Z.; Matveeva, T.; Bogomaz, D.; Berner, D.K.; Cavin, C.A.; Castlebury, L.A. First Report of Stem Canker of Salsola tragus Caused by Diaporthe eres in Russia. Plant Dis. 2009, 93, 110. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, N. The Rust Flora of Japan; Tsukuba Shuppankai: Tsukuba, Japan, 1992; ISBN 978-4-924753-11-2. [Google Scholar]
- Oner, M.; Dizbay, M.; Ekmekci, S. An Investigation of Some Leaf Rust, Smuts, Powdery Mildews and Leaf Spots Occurring of the Natural Flora of Southern Egean Region. Bitki Derg. 1974, 1, 426–431. [Google Scholar]
- Gjærum, H.B. Two Rust Species from the Salvage Islands. Bocagiana Mus. Munic. Funchal 1975, 41, 1–3. [Google Scholar]
- Dias, M.R.d.S.; Lucas, M.T. Fungi of Madeira and Selvagem Islands. Bol. Soc. Broteriana 1980, 53, 469–473. [Google Scholar]
- Le Houérou, H.N. The Role of Saltbushes (Atriplex spp.) in Arid Land Rehabilitation in the Mediterranean Basin: A Review. Agroforest. Syst. 1992, 18, 107–148. [Google Scholar] [CrossRef]
- Fries, E.M. Epicrisis Systematis Mycologici; E Typographia Academica, Upsaliae: Uppsala, Sweden, 1836. [Google Scholar]
- Fries, E.M. Summa Vegetabilium Scandinaviae; E Typographia Academica, Upsaliae: Uppsala, Sweden, 1845. [Google Scholar]
- Gäumann, E. Zur Kenntnis Der Chenopodiaceen Bewohnenden Peronospora-Arten. Mitteilungen Naturforschenden Ges. Bern. 1918, 45–66. [Google Scholar] [CrossRef]
- Săvulescu, T.; Rayss, T. Contribution a La Connaissance Des Peronsporacees de Roumanie. Ann. Mycol. 1932, 28, 297–320. [Google Scholar]
- Yerkes, W.D.; Shaw, C.G. Taxonomy of the Peronospora Species on Cruciferae and Chenopodiaceae. Phytopathology 1959, 49, 499–507. [Google Scholar]
- Voglmayr, H. Phylogenetic Relationships of Peronospora and Related Genera Based on Nuclear Ribosomal ITS Sequences. Mycol. Res. 2003, 107, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Hong, S.-B.; Shin, H.-D. Re-Consideration of Peronospora farinosa Infecting Spinacia oleracea as Distinct Species, Peronospora effusa. Mycol. Res. 2007, 111, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Denchev, C.M.; Shin, H.-D. Morphological and Molecular Analyses Support the Existence of Host-Specific Peronospora Species Infecting Chenopodium. Mycopathologia 2008, 165, 155–164. [Google Scholar] [CrossRef]
- Thines, M.; Choi, Y.-J. Evolution, Diversity, and Taxonomy of the Peronosporaceae, with Focus on the Genus Peronospora. Phytopathology 2016, 106, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Danielsen, S.; Lübeck, M.; Hong, S.-B.; Delhey, R.; Shin, H.-D. Morphological and Molecular Characterization of the Causal Agent of Downy Mildew on Quinoa (Chenopodium quinoa). Mycopathologia 2010, 169, 403–412. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Thines, M. (2288) Proposal to Reject the Name Botrytis farinosa (Peronospora farinosa) (Peronosporaceae: Oomycetes). Taxon 2014, 63, 675–676. [Google Scholar] [CrossRef]
- May, T.W. Report of the Nomenclature Committee for Fungi—20. IMA Fungus 2017, 8, 189–203. [Google Scholar] [CrossRef]
- Wilson, K.L. Report of the General Committee: 18. Taxon 2017, 66, 742–744. [Google Scholar] [CrossRef]
- Kempenaar, C.; Horsten, P.J.F.M.; Scheepens, P.C. Spore Germination and Disease Development after Application of Pycnidiospores of Ascochyta caulina to Chenopodium album Plants. Eur. J. Plant Pathol. 1996, 102, 143–153. [Google Scholar] [CrossRef]
- van der Aa, H.; Vanev, S. A Revision of the Species Described in Phyllosticta; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002. [Google Scholar]
- de Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Redisposition of Phoma-like Anamorphs in Pleosporales. Stud. Mycol. 2013, 75, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Wanasinghe, D.N.; Hyde, K.D.; Jeewon, R.; Crous, P.W.; Wijayawardene, N.N.; Jones, E.B.G.; Bhat, D.J.; Phillips, A.J.L.; Groenewald, J.Z.; Dayarathne, M.C.; et al. Phylogenetic Revision of Camarosporium (Pleosporineae, Dothideomycetes) and Allied Genera. Stud. Mycol. 2017, 87, 207–256. [Google Scholar] [CrossRef]
- Weiland, J.; Koch, G. Sugarbeet Leaf Spot Disease (Cercospora beticola Sacc.). Mol. Plant Pathol. 2004, 5, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Palti, J. The Leveillula Mildews. Bot. Rev. 1988, 54, 423–535. [Google Scholar] [CrossRef]
- Heluta, V.P.; Minter, D.W. Leveillula cylindrospora. [Descriptions of Fungi and Bacteria]. Descr. Fungi Bact. 1998, Sheet 1373. [Google Scholar] [CrossRef]
- Takamatsu, S. Origin and Evolution of the Powdery Mildews (Ascomycota, Erysiphales). Mycoscience 2012, 54, 75–86. [Google Scholar] [CrossRef]
- Davy, A.J.; Scott, R.; Cordazzo, C.V. Biological Flora of the British Isles: Cakile maritima Scop. J. Ecol. 2006, 94, 695–711. [Google Scholar] [CrossRef]
- Bock, C.H.; Thrall, P.H.; Burdon, J.J. Genetic Structure of Populations of Alternaria brassicicola Suggests the Occurrence of Sexual Recombination. Mycol. Res. 2005, 109, 227–236. [Google Scholar] [CrossRef]
- Pinto, V.E.F.; Patriarca, A. Alternaria Species and Their Associated Mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; pp. 13–32. ISBN 978-1-4939-6707-0. [Google Scholar]
- Holub, E.B. Phenotypic and Genotypic Variation in the Interaction between Arabidopsis thaliana and Albugo candida. Mol. Plant-Microbe Interact. MPMI 1995, 8, 916. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Hong, S.-B.; Shin, H.-D. Genetic Diversity within the Albugo candida Complex (Peronosporales, Oomycota) Inferred from Phylogenetic Analysis of ITS rDNA and COX2 mtDNA Sequences. Mol. Phylogenet. Evol. 2006, 40, 400–409. [Google Scholar] [CrossRef]
- Voglmayr, H.; Riethmüller, A. Phylogenetic Relationships of Albugo Species (White Blister Rusts) Based on LSU rDNA Sequence and Oospore Data. Mycol. Res. 2006, 110, 75–85. [Google Scholar] [CrossRef]
- Constantinescu, O.; Fatehi, J. Peronospora-like Fungi (Chromista, Peronosporales) Parasitic on Brassicaceae and Related Hosts. Nova Hedwig. 2002, 74, 291–338. [Google Scholar] [CrossRef]
- Abdallah, A.; Zouhaier, B.; Rabhi, M.; Abdelly, C.; Smaoui, A. Environmental Eco-Physiology and Economical Potential of the Halophyte Crithmum maritimum L. (Apiaceae). J. Med. Plant Res. 2011, 5, 3564–3571. [Google Scholar]
- Renna, M. Reviewing the Prospects of Sea Fennel (Crithmum maritimum L.) as Emerging Vegetable Crop. Plants 2018, 7, 92. [Google Scholar] [CrossRef]
- Vánky, K. Taxonomic Studies on Ustilaginomycetes—29. Mycotaxon 2009, 110, 289–324. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A Genus of Endophytic, Saprobic and Plant Pathogenic Fungi. Persoonia Mol. Phylogeny Evol. Fungi 2013, 31, 1–41. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Farr, D.F.; Rossman, A.Y.; Jaklitsch, W. Diaporthe angelicae comb. nov., a Modern Description and Placement of Diaporthopsis in Diaporthe. Mycoscience 2003, 44, 203–208. [Google Scholar] [CrossRef]
- Ellis, J.B.; Kellerman, W.A. New Kansas Fungi. J. Mycol. 1887, 3, 102–105. [Google Scholar] [CrossRef]
- Young, J.A.; Palmquist, D.E.; Blank, R.R. The Ecology and Control of Perennial Pepperweed (Lepidium latifolium L.). Weed Technol. 1998, 12, 402–405. [Google Scholar] [CrossRef]
- Kaur, T.; Hussain, K.; Koul, S.; Vishwakarma, R.; Vyas, D. Evaluation of Nutritional and Antioxidant Status of Lepidium latifolium Linn.: A Novel Phytofood from Ladakh. PLoS ONE 2013, 8, e69112. [Google Scholar] [CrossRef]
- Conde-Rioll, M.; Gajate, C.; Fernández, J.J.; Villa-Pulgarin, J.A.; Napolitano, J.G.; Norte, M.; Mollinedo, F. Antitumor Activity of Lepidium latifolium and Identification of the Epithionitrile 1-Cyano-2,3-Epithiopropane as Its Major Active Component. Mol. Carcinog. 2018, 57, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Manganiello, G.; Nicastro, N.; Cardi, T.; Carotenuto, F. Powdery Mildew Caused by Erysiphe cruciferarum on Wild Rocket (Diplotaxis tenuifolia): Hyperspectral Imaging and Machine Learning Modeling for Non-Destructive Disease Detection. Agriculture 2021, 11, 337. [Google Scholar] [CrossRef]
- Hasan, M.; Ahmad-Hamdani, M.S.; Rosli, A.M.; Hamdan, H. Bioherbicides: An Eco-Friendly Tool for Sustainable Weed Management. Plants 2021, 10, 1212. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Johnson, D.E. Seed Germination Ecology of Portulaca oleracea L.: An Important Weed of Rice and Upland Crops. Ann. Appl. Biol. 2009, 155, 61–69. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Karkanis, A.; Fernandes, Â.; Barros, L.; Ferreira, I.C.F.R.; Ntatsi, G.; Petrotos, K.; Lykas, C.; Khah, E. Chemical Composition and Yield of Six Genotypes of Common Purslane (Portulaca oleracea L.): An Alternative Source of Omega-3 Fatty Acids. Plant Foods Hum. Nutr. 2015, 70, 420–426. [Google Scholar] [CrossRef]
- Thines, M.; Spring, O. A Revision of Albugo (Chromista, Peronosporomycetes). Mycotaxon 2005, 92, 443–458. [Google Scholar]
- Tan, Y.P.; Madrid, H.; Crous, P.W.; Shivas, R.G. Johnalcornia gen. et. comb. nov., and Nine New Combinations in Curvularia Based on Molecular Phylogenetic Analysis. Australas. Plant Pathol. 2014, 43, 589–603. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH); Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.-A.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; et al. Pest Categorisation of Colletotrichum fructicola. EFSA J. 2021, 19, e06803. [Google Scholar] [CrossRef] [PubMed]
- Nel, W.J.; Duong, T.A.; Wingfield, B.D.; Wingfield, M.J.; de Beer, Z.W. A New Genus and Species for the Globally Important, Multihost Root Pathogen Thielaviopsis basicola. Plant Pathol. 2018, 67, 871–882. [Google Scholar] [CrossRef]
- French-Monar, R.D.; Jones, J.B.; Roberts, P.D. Characterization of Phytophthora capsici Associated with Roots of Weeds on Florida Vegetable Farms. Plant Dis. 2006, 90, 345–350. [Google Scholar] [CrossRef]
- Barboza, E.A.; Cabral, C.S.; Rossato, M.; Martins, F.H.S.R.; Reis, A. Pythium and Phytopythium Species Associated with Weeds Collected in Vegetable Production Fields in Brazil. Lett. Appl. Microbiol. 2022, 74, 796–808. [Google Scholar] [CrossRef]
- Sacristán, S.; García-Arenal, F. The evolution of virulence and pathogenicity in plant pathogen populations. Mol. Plant Pathol. 2008, 9, 369–384. [Google Scholar] [CrossRef]
- Ventura, Y.; Sagi, M. Halophyte Crop Cultivation: The Case for Salicornia and Sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar] [CrossRef]
- Cárdenas-Pérez, S.; Piernik, A.; Chanona-Pérez, J.J.; Grigore, M.N.; Perea-Flores, M.J. An Overview of the Emerging Trends of the Salicornia L. Genus as a Sustainable Crop. Environ. Exp. Bot. 2021, 191, 104606. [Google Scholar] [CrossRef]
- Mosyakin, S.L. A Taxonomic Synopsis Of The Genus Salsola (Chenopodiaceae) In North America. Ann. Mo. Bot. Gard. 1996, 83, 387–395. [Google Scholar] [CrossRef]
- Altay, V.; Ozturk, M. The Genera Salsola and Suaeda (Amaranthaceae) and Their Value as Fodder. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture; Grigore, M.-N., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 2333–2344. ISBN 978-3-030-57635-6. [Google Scholar]
- Bruckart, W.L.; Thomas, J.L.; Frederick, R.D.; Aime, M.C.; Abbasi, M. First Report of a Rust Disease Caused by Uromyces sp. on Suaeda californica in California. Plant Dis. 2019, 103, 1784. [Google Scholar] [CrossRef]
- Berner, D.K.; Bruckart, W.L.; Cavin, C.A.; Michael, J.L.; Carter, M.L.; Luster, D.G. Best Linear Unbiased Prediction of Host-Range of the Facultative Parasite Colletotrichum gloeosporioides f. sp. salsolae, a Potential Biological Control Agent of Russian Thistle. Biol. Control 2009, 51, 158–168. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides Species Complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef]
- Centofanti, T.; Bañuelos, G. Evaluation of the Halophyte Salsola soda as an Alternative Crop for Saline Soils High in Selenium and Boron. J. Environ. Manag. 2015, 157, 96–102. [Google Scholar] [CrossRef]
- Iannuzzi, A.M.; Moschini, R.; De Leo, M.; Pineschi, C.; Balestri, F.; Cappiello, M.; Braca, A.; Del-Corso, A. Chemical Profile and Nutraceutical Features of Salsola soda (Agretti): Anti-Inflammatory and Antidiabetic Potential of Its Flavonoids. Food Biosci. 2020, 37, 100713. [Google Scholar] [CrossRef]
- Castañeda-Loaiza, V.; Oliveira, M.; Santos, T.; Schüler, L.; Lima, A.R.; Gama, F.; Salazar, M.; Neng, N.R.; Nogueira, J.M.F.; Varela, J.; et al. Wild vs Cultivated Halophytes: Nutritional and Functional Differences. Food Chem. 2020, 333, 127536. [Google Scholar] [CrossRef] [PubMed]
- Shahi, M.; Saaghari, M.; Esfahan, E.Z.; Jaimand, K. Investigation on Potential of Suaeda fruticosa as a Source of Edible Oil. J. Bio Environ. Sci. 2013, 3, 101–107. [Google Scholar]
- Anderson, J.P.; Gleason, C.A.; Foley, R.C.; Thrall, P.H.; Burdon, J.B.; Singh, K.B. Plants versus pathogens: An evolutionary arms race. Funct. Plant Biol. 2010, 37, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Dikilitas, M.; Karakas, S. Chapter 8—Crop Plants under Saline-Adapted Fungal Pathogens: An Overview. In Emerging Technologies and Management of Crop Stress Tolerance; Ahmad, P., Rasool, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 173–192. ISBN 978-0-12-800875-1. [Google Scholar]
- Danti, S.; Broggio, M. Impact of Salinity Stress on the Biometric Responses of an Isolate of Verticillium dahliae Kleb. from Potato. In Proceedings of the Seventh International Verticillium Symposium, Cape Sounion, Athens, Greece, 6–10 October 1997; p. 53. [Google Scholar]
- Dikilitas, M. Effect of Salinity and Its Interactions with Verticillium albo-atrum on the Disease Develoment in Tomato (Lycopersicon esculentum Mill.) and Lucerne (Medicago sativa & M. Media) Plants. Ph.D. Thesis, University of Wales, Swansea, UK, 2003. [Google Scholar]
- Bhai, S.; Muthuswamy, A.; Srinivasan, V. Validation of Farmer’s Practice of Using Sodium Chloride for Controlling Foot Rot Disease of Black Pepper (Piper nigrum). Indian J. Agric. Sci. 2009, 79, 722–726. [Google Scholar]
- Hasan, H. Gibberellin and Auxin-Indole Production by Plant Root-Fungi and Their Biosynthesis under Salinity-Calcium Interaction. Acta Microbiol. Immunol. Hung. 2005, 49, 105–118. [Google Scholar] [CrossRef]
- Porter, D.M.; Adamsen, F.J. Effect of Sodic Water and Irrigation on Sodium Levels and the Development of Early Leaf Spot in Peanuts. Plant Dis. 1993, 77, 480. [Google Scholar] [CrossRef]
- Firdous, H.; Shahzad, S. Effect of Some Salts on in Vitro Growth of Fusarium solani. Pak. J. Bot. 2001, 33, 117–124. [Google Scholar]
- Ragazzi, A.; Vecchio, V.; Dellavalle, I.; Cucchi, A.; Mancini, F. Variations in the Pathogenicity of Fusarium oxysporum f. sp. vasinfectum in Relation to the Salinity of the Nutrient Medium. J. Plant Dis. Prot. 1994, 101, 263–266. [Google Scholar]
- El-Abyad, M.S.; Abu-Taleb, A.M.; Khalil, M.S. Impact of Salinity Stress on Soil-Borne Fungi of Sugarbeet. Plant Soil. 1992, 143, 75–83. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Al-Masoudi, R.S.; Al-Habsi, N.; Al-Said, F.A.; Al-Rawahy, S.A.; Ahmed, M.; Deadman, M.L. Effect of Salinity on Pythium Damping-off of Cucumber and on the Tolerance of Pythium aphanidermatum. Plant Pathol. 2010, 59, 112–120. [Google Scholar] [CrossRef]
- Russo, G.L.; Langellotti, A.L.; Martín-García, B.; Verardo, V.; Romano, R.; Sacchi, R.; Masi, P. New Biotechnological Production of EPA by Pythium irregulare Using Alternative Sustainable Media Obtained from Food Industry By-Products and Waste. Sustainability 2023, 15, 1147. [Google Scholar] [CrossRef]
- Ambikapathy, J.; Marshall, J.S.; Hocart, C.H.; Hardham, A.R. The Role of Proline in Osmoregulation in Phytophthora nicotianae. Fungal Genet. Biol. 2002, 35, 287–299. [Google Scholar] [CrossRef]
- Luard, E.J. Growth and Accumulation of Solutes by Phytophthora cinnamomi and Other Lower Fungi in Response to Changes in External Osmotic Potential. Microbiology 1982, 128, 2583–2590. [Google Scholar] [CrossRef]
- Lacalendola, N.; Tayagui, A.; Ting, M.; Malmstrom, J.; Nock, V.; Willmott, G.R.; Garrill, A. Biomechanical Responses of Encysted Zoospores of the Oomycete Achlya bisexualis to Hyperosmotic Stress Are Consistent with an Ability to Turgor Regulate. Fungal Genet. Biol. 2022, 159, 103676. [Google Scholar] [CrossRef]
- Gostinčar, C.; Lenassi, M.; Gunde-Cimerman, N.; Plemenitaš, A. Chapter 3—Fungal Adaptation to Extremely High Salt Concentrations. In Advances in Applied Microbiology; Laskin, A.I., Sariaslani, S., Gadd, G.M., Eds.; Academic Press: New York, NY, USA, 2011; Volume 77, pp. 71–96. [Google Scholar]
- Hohmann, S. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef]
- Fettich, M.; Lenassi, M.; Veranič, P.; Gunde-Cimerman, N.; Plemenitaš, A. Identification and Characterization of Putative Osmosensors, HwSho1A and HwSho1B, from the Extremely Halotolerant Black Yeast Hortaea werneckii. Fungal Genet. Biol. 2011, 48, 475–484. [Google Scholar] [CrossRef]
- Lenassi, M.; Plemenitaš, A. Novel Group VII Histidine Kinase HwHhk7B from the Halophilic Fungi Hortaea werneckii Has a Putative Role in Osmosensing. Curr. Genet. 2007, 51, 393–405. [Google Scholar] [CrossRef]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus nidulans HOG Pathway Is Activated Only by Two-Component Signaling Pathway in Response to Osmotic Stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Chen, Y.; Liu, Y.; Zhang, C.; Ma, Z. The Transmembrane Protein FgSho1 Regulates Fungal Development and Pathogenicity via the MAPK Module Ste50-Ste11-Ste7 in Fusarium graminearum. New Phytol. 2015, 206, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-J.; Zhou, L.; Yin, C.-M.; Zhang, D.-D.; Klosterman, S.J.; Wang, B.-L.; Song, J.; Wang, D.; Hu, X.-P.; Subbarao, K.V.; et al. The Verticillium dahliae Sho1-MAPK Pathway Regulates Melanin Biosynthesis and Is Required for Cotton Infection. Environ. Microbiol. 2019, 21, 4852–4874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, K.; Zhang, X.; Song, W.; Zhao, Q.; Dong, Y.; Guo, M.; Zheng, X.; Zhang, Z. A Two-Component Histidine Kinase, MoSLN1, Is Required for Cell Wall Integrity and Pathogenicity of the Rice Blast Fungus, Magnaporthe oryzae. Curr. Genet. 2010, 56, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Zhang, S.; Zhou, X.; Wang, C.; Xiang, P.; Zheng, Q.; Xu, J.-R. The FgHOG1 Pathway Regulates Hyphal Growth, Stress Responses, and Plant Infection in Fusarium graminearum. PLoS ONE 2012, 7, e49495. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.-L.; Chen, L.-H.; Chung, K.-R. How the Pathogenic Fungus Alternaria alternata Copes with Stress via the Response Regulators SSK1 and SHO1. PLoS ONE 2016, 11, e0149153. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Wang, Y.; Yu, J.; Xiong, D.; Zhao, H.; Tian, C. The Mitogen-Activated Protein Kinase Kinase VdPbs2 of Verticillium dahliae Regulates Microsclerotia Formation, Stress Response, and Plant Infection. Front. Microbiol. 2016, 7, 1532. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yin, D.; Yin, Y.; Cao, Y.; Ma, Z. The Response Regulator BcSkn7 Is Required for Vegetative Differentiation and Adaptation to Oxidative and Osmotic Stresses in Botrytis cinerea. Mol. Plant Pathol. 2015, 16, 276–287. [Google Scholar] [CrossRef]
- Kogej, T.; Ramos, J.; Plemenitaš, A.; Gunde-Cimerman, N. The Halophilic Fungus Hortaea werneckii and the Halotolerant Fungus Aureobasidium pullulans Maintain Low Intracellular Cation Concentrations in Hypersaline Environments. Appl. Environ. Microbiol. 2005, 71, 6600–6605. [Google Scholar] [CrossRef]
- Gorjan, A.; Plemenitaš, A. Identification and Characterization of ENA ATPases HwENA1 and HwENA2 from the Halophilic Black Yeast Hortaea werneckii. FEMS Microbiol. Lett. 2006, 265, 41–50. [Google Scholar] [CrossRef]
- Vaupotič, T.; Plemenitaš, A. Differential Gene Expression and Hog1 Interaction with Osmoresponsive Genes in the Extremely Halotolerant Black Yeast Hortaea werneckii. BMC Genom. 2007, 8, 280. [Google Scholar] [CrossRef]
- Lenassi, M.; Zajc, J.; Gostinčar, C.; Gorjan, A.; Gunde-Cimerman, N.; Plemenitaš, A. Adaptation of the Glycerol-3-Phosphate Dehydrogenase Gpd1 to High Salinities in the Extremely Halotolerant Hortaea werneckii and Halophilic Wallemia ichthyophaga. Fungal Biol. 2011, 115, 959–970. [Google Scholar] [CrossRef]
- Petrovič, U.; Gunde-Cimerman, N.; Plemenitaš, A. Cellular Responses to Environmental Salinity in the Halophilic Black Yeast Hortaea werneckii. Mol. Microbiol. 2002, 45, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; van Voorst, F.; Martins, A.; Neves, L.; Oliveira, R.; Kielland-Brandt, M.C.; Lucas, C.; Brandt, A. A Member of the Sugar Transporter Family, Stl1p Is the Glycerol/H+ Symporter in Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, S.; Antelo, L.; Grünewald, C.; Yemelin, A.; Andresen, K.; Jacob, S. Rapid Adaptation of Signaling Networks in the Fungal Pathogen Magnaporthe oryzae. BMC Genom. 2019, 20, 763. [Google Scholar] [CrossRef] [PubMed]
- Kogej, T.; Wheeler, M.H.; Lanišnik Rižner, T.; Gunde-Cimerman, N. Evidence for 1,8-Dihydroxynaphthalene Melanin in Three Halophilic Black Yeasts Grown under Saline and Non-Saline Conditions. FEMS Microbiol. Lett. 2004, 232, 203–209. [Google Scholar] [CrossRef]
- Turk, M.; Méjanelle, L.; Šentjurc, M.; Grimalt, J.O.; Gunde-Cimerman, N.; Plemenitaš, A. Salt-Induced Changes in Lipid Composition and Membrane Fluidity of Halophilic Yeast-like Melanized Fungi. Extremophiles 2004, 8, 53–61. [Google Scholar] [CrossRef]
- Turk, M.; Abramović, Z.; Plemenitaš, A.; Gunde-Cimerman, N. Salt Stress and Plasma-Membrane Fluidity in Selected Extremophilic Yeasts and Yeast-like Fungi. FEMS Yeast Res. 2007, 7, 550–557. [Google Scholar] [CrossRef]
Figure 1.
Documents published over time, distinguished in research papers and checklists on fungi or fungal-like pathogens of Mediterranean halophytic species. Documents are chronologically grouped by decades; those published before 1950 are gathered under the same period. Studies addressing the taxonomy of species were considered research papers. Data were collected using the USDA Fungal Database and different literature databases (i.e., JSTOR, Google Scholar, SCOPUS, and Web of Science).
Figure 1.
Documents published over time, distinguished in research papers and checklists on fungi or fungal-like pathogens of Mediterranean halophytic species. Documents are chronologically grouped by decades; those published before 1950 are gathered under the same period. Studies addressing the taxonomy of species were considered research papers. Data were collected using the USDA Fungal Database and different literature databases (i.e., JSTOR, Google Scholar, SCOPUS, and Web of Science).

Figure 2.
Diversity of fungal and fungal-like orders reported on major Mediterranean halophytes according to the literature search on different databases (JSTOR, Google Scholar, SCOPUS, and Web of Science). Each color corresponds to the specific phylum (orange: Ascomycota; blue: Basidiomycota; green: Oomycota). Numbers indicate the different pathogenic species within each order.
Figure 2.
Diversity of fungal and fungal-like orders reported on major Mediterranean halophytes according to the literature search on different databases (JSTOR, Google Scholar, SCOPUS, and Web of Science). Each color corresponds to the specific phylum (orange: Ascomycota; blue: Basidiomycota; green: Oomycota). Numbers indicate the different pathogenic species within each order.

Figure 3.
The distribution of fungal and fungal-like orders among different Mediterranean halophytic plant genera. Different colors indicate different orders. The fraction of each order matches the number of reported species within the same order on the specific host genus.
Figure 3.
The distribution of fungal and fungal-like orders among different Mediterranean halophytic plant genera. Different colors indicate different orders. The fraction of each order matches the number of reported species within the same order on the specific host genus.


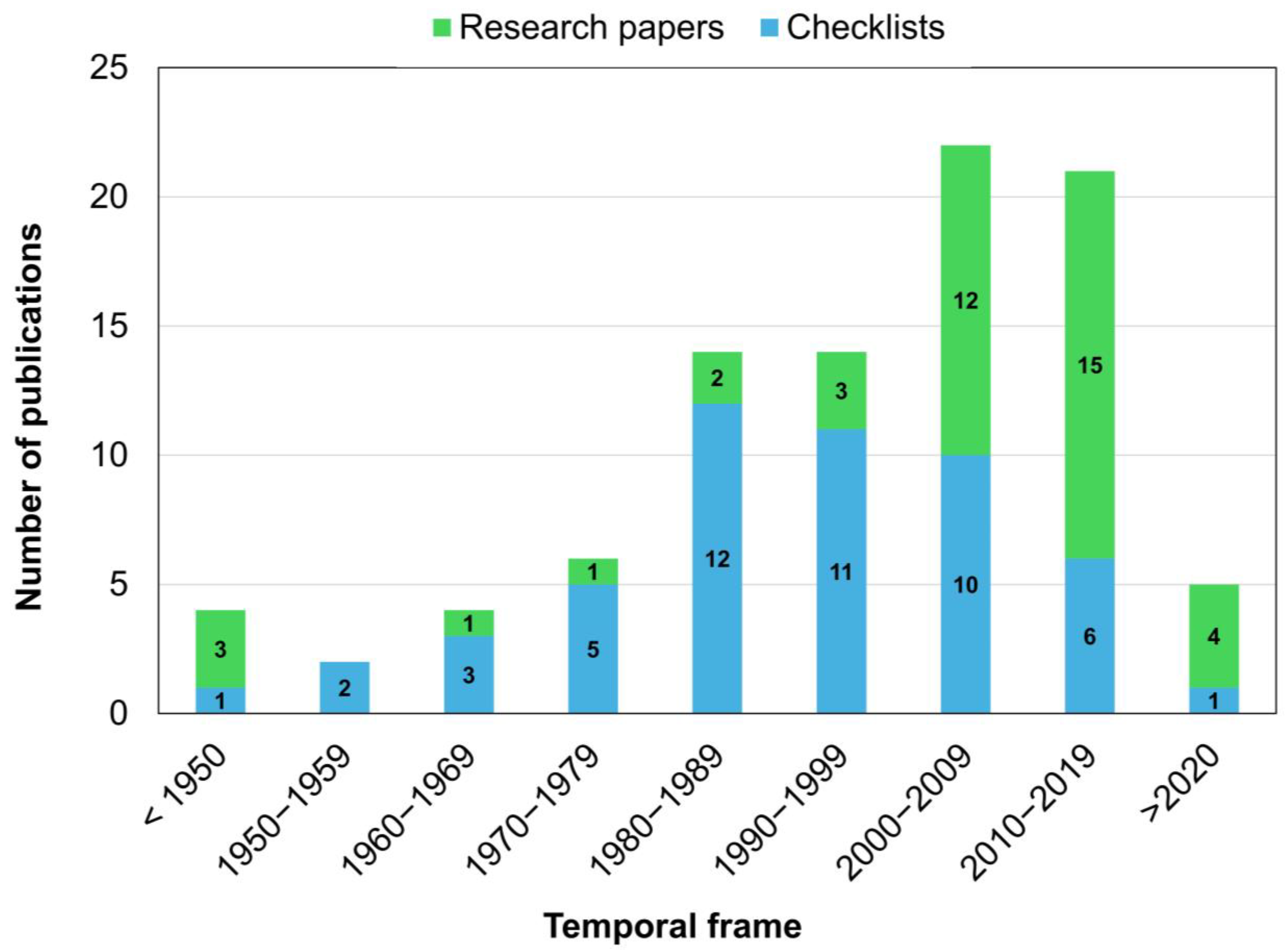{kind=link}
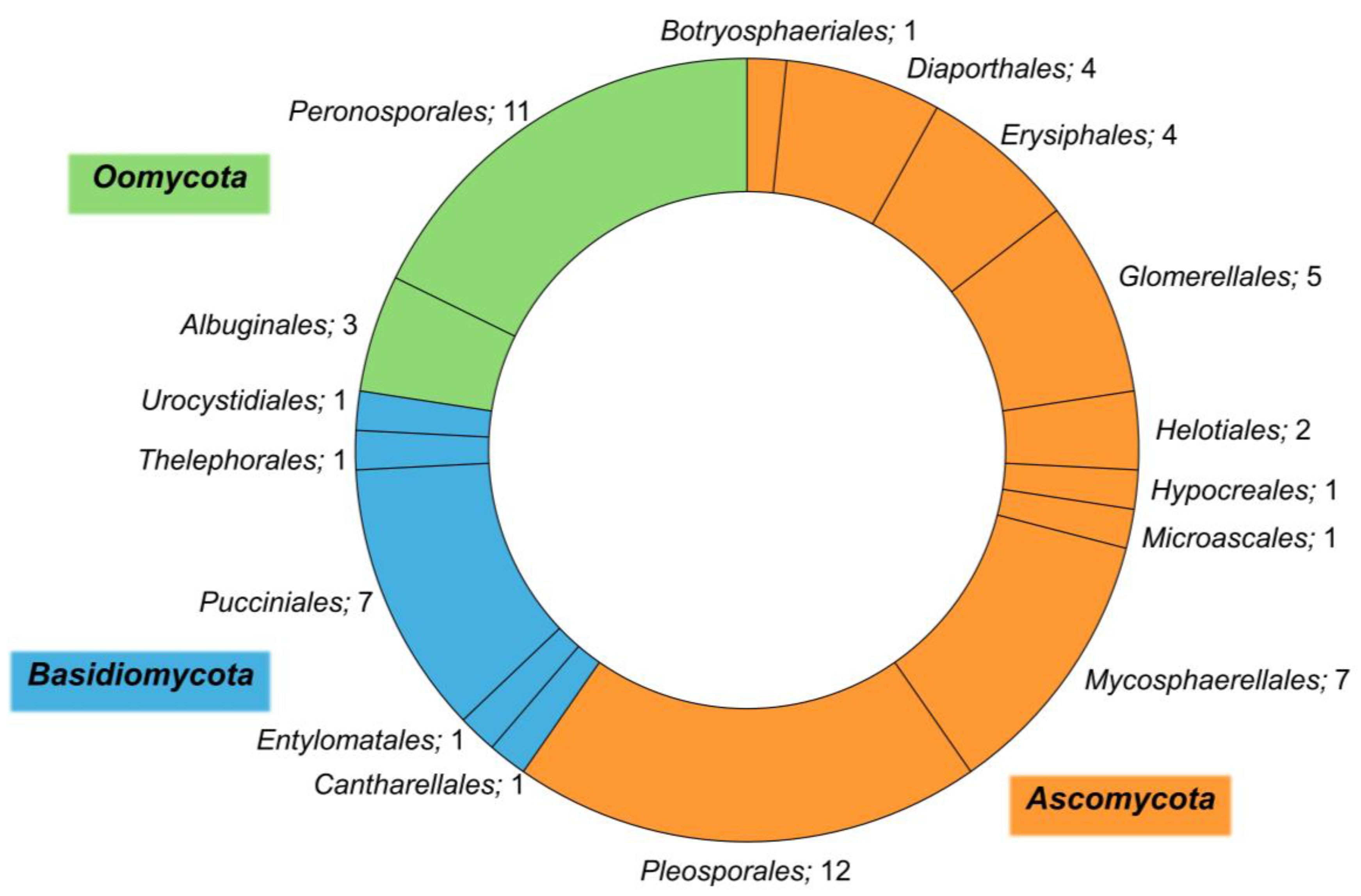{kind=link}
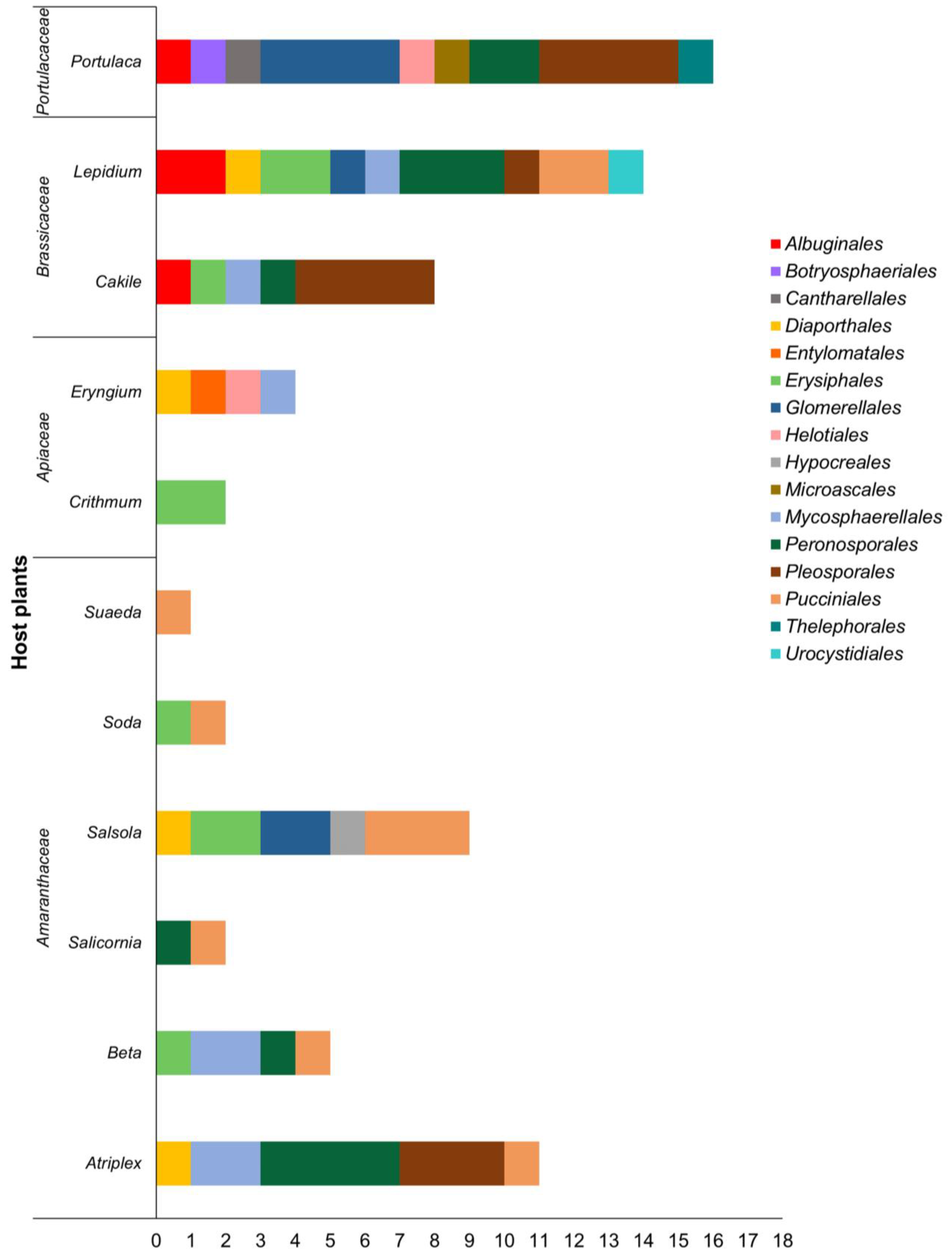{kind=link}
Table 1.
Halophytic plant species considered in the review. Plant selection was made based on the current literature; other species were added on their geographical distribution (according to the GBIF database) and if present in the eHALOPH salt-tolerant plant database.
Table 1.
Halophytic plant species considered in the review. Plant selection was made based on the current literature; other species were added on their geographical distribution (according to the GBIF database) and if present in the eHALOPH salt-tolerant plant database.
| Botanical Name | Family | Common Name |
|---|---|---|
| Atriplex hortensis L. | Amaranthaceae | Garden orache |
| Atriplex littoralis L. | Amaranthaceae | Grassleaf orache |
| Atriplex nummularia Lindl. | Amaranthaceae | Oldman saltbush |
| Atriplex patula L. | Amaranthaceae | Common orache |
| Atriplex prostrata Boucher ex DC (=Syn.: Atriplex hastata L. var. prostrata (Boucher ex DC.) Lange) | Amaranthaceae | Hastate orache |
| Beta vulgaris L. subsp. maritima (L.) Arcang. | Amaranthaceae | Sea beet |
| Cakile maritima Scop. subsp. maritima | Brassicaceae | Searocket |
| Crithmum maritimum L. | Apiaceae | Sea fennel |
| Eryngium maritimum L. | Apiaceae | Sea holly |
| Lepidium latifolium L. | Brassicaceae | Broadleaved pepperweed |
| Lepidium perfoliatum L. | Brassicaceae | Clasping pepperweed |
| Lepidium ruderale L. | Brassicaceae | Stinking pepperweed |
| Lepidium subulatum L. | Brassicaceae | - |
| Portulaca oleracea L. | Portulacaceae | Common purslane |
| Salicornia perennans Willd. subsp. perennans (=Syn.: Salicornia europaea L.) | Amaranthaceae | Glasswort |
| Salicornia perennis Mill. (=Syn.: Salicornia perennis (Mill.) A.J. Scott subsp. perennis) | Amaranthaceae | Perennial glasswort |
| Salicornia ramosissima (Hook.f.) J. Woods ex W.A. Clarke & E.S. Marshall | Amaranthaceae | Twiggy glasswort |
| Salsola kali L. (=Syn.: Kali turgidum (Dumort.) Gutermann) | Amaranthaceae | Common saltwort |
| Salsola tragus L. (=Syn.: Kali tragus subsp. tragus (L.) Scoop.) | Amaranthaceae | Russian thistle |
| Salsola soda L. (=Syn.: Soda inermis Fourr.) | Amaranthaceae | Monk’s beard |
| Suaeda maritima (L.) Dumort. (=Syn.: Chenopodium maritimum L.) | Amaranthaceae | Sea-blite |
| Suaeda vera J.F. Gmel (=Syn.: Suaeda fruticosa (L.) Forssk. subsp. vera (J.F. Gmel.) Maire & Weiller) | Amaranthaceae | Shrubby sea-blite |
Table 2.
Plant pathogenic species associated with major Mediterranean halophytes. Host species, diseases caused, origin of the specimens, and bibliographic references are reported.
Table 2.
Plant pathogenic species associated with major Mediterranean halophytes. Host species, diseases caused, origin of the specimens, and bibliographic references are reported.
| Plant Genus | Plant Species | Disease | Pathogenic Species | Order | Origin | Reference(s) |
|---|---|---|---|---|---|---|
| Atriplex | A. hortensis | Downy mildew | Peronospora atriplicis-hortensis | Peronosporales | Herbarium specimen(s) | [43] |
| Leaf spot | Ascochyta caulina | Pleosporales | Herbarium specimen(s) | [44] | ||
| Passalora spegazzinii | Mycosphaerellales | Wild | [45] | |||
| Stagonospora atriplicis | Pleosporales | Herbarium specimen(s) | [46] | |||
| A. littoralis | Downy mildew | Peronospora litoralis | Peronosporales | Herbarium specimen(s) | [43,47] | |
| Leaf spot | Ascochyta caulina | Pleosporales | Herbarium specimen(s) | [44] | ||
| Stagonospora atriplicis | Pleosporales | Herbarium specimen(s) | [46] | |||
| A. nummularia | Shoot blight | Diaporthe inconspicua | Diaporthales | Wild | [48] | |
| A. patula | Downy mildew | Peronospora minor. | Peronosporales | Herbarium specimen(s) | [43,47] | |
| Wild | [49,50,51,52,53] | |||||
| Leaf spot | Ascochyta caulina | Pleosporales | Herbarium specimen(s) | [44] | ||
| Passalora dubia | Mycosphaerellales | - | [54,55] | |||
| Wild | [56,57,58] | |||||
| Passalora spegazzinii | Mycosphaerellales | - | [55,59] | |||
| Stagonospora atriplicis | Pleosporales | Herbarium specimen(s) | [46,60] | |||
| Rust | Uromyces peckianus | Pucciniales | Wild | [61] | ||
| A. prostrata (=Syn.: A. hastata) | Downy mildew | Peronospora atriplicis-hastatae | Peronosporales | Herbarium specimen(s) | [43,47] | |
| Leaf spot | Ascochyta atriplicis | Pleosporales | - | [54] | ||
| Herbarium specimen(s) | [62] | |||||
| Ascochyta caulina | Pleosporales | Herbarium specimen(s) | [44] | |||
| Stagonospora atriplicis | Pleosporales | Herbarium specimen(s) | [46] | |||
| Beta | B. vulgaris subsp. maritima | Downy mildew | Peronospora schachtii | Peronosporales | Wild | [53] |
| Leaf blotch | Septoria betae | Mycosphaerellales | - | [60] | ||
| Leaf spot | Cercospora beticola | Mycosphaerellales | Wild | [63] | ||
| Powdery mildew | Leveillula taurica | Erysiphales | - | [64] | ||
| Rust | Uromyces betae | Pucciniales | Wild | [65,66] | ||
| Cakile | C. maritima | Black leaf spot | Alternaria alternata | Pleosporales | Wild | [67] |
| Wild and pot (artificial inoculation on detached leaves) | [68] | |||||
| Pot (artificial inoculation in salinity-NaCl 200 mM) | [69] | |||||
| Alternaria arborescens | Pleosporales | Wild and pot (artificial inoculation on detached leaves) | [68] | |||
| Alternaria brassicicola | Pleosporales | Wild | [67,70] | |||
| Alternaria mali | Pleosporales | Wild and pot (artificial inoculation on detached leaves) | [68] | |||
| Downy mildew | Hyaloperonospora parasitica | Peronosporales | Wild | [71] | ||
| Powdery mildew | Erysiphe cruciferarum | Erysiphales | - | [64] | ||
| Ring spot | Mycosphaerella brassicicola | Mycosphaerellales | Herbarium specimen(s) | [72] | ||
| White rust | Albugo candida | Albuginales | Wild | [50,73,74] | ||
| Crithmum | C. maritimum | Powdery mildew | Leveillula taurica | Erysiphales | - | [64,75] |
| Wild | [76] | |||||
| Leveillula lanuginosa | Erysiphales | - | [75] | |||
| Eryngium | E. maritimum | Leaf spot | Cylindrosporium eryngii | Helotiales | - | [77] |
| Pseudocercosporella eryngii | Mycosphaerellales | - | [54] | |||
| Wild and herbarium specimen(s) | [78] | |||||
| Leaf smut | Entyloma eryngii-maritimi | Entylomatales | Herbarium specimen(s) | [79] | ||
| Stem necrosis | Diaporthe angelicae | Diaporthales | - | [80] | ||
| Wild | [81] | |||||
| Lepidium | L. latifolium | Canker and dieback | Cytospora ribis | Diaporthales | Wild | [82] |
| Downy mildew | Perofascia lepidii | Peronosporales | Herbarium specimen(s) | [83] | ||
| Leaf spot | Alternaria brassicae | Pleosporales | Herbarium specimen(s) | [84] | ||
| Cercospora bizzozeriana | Mycosphaerellales | Herbarium specimen(s) | [85] | |||
| Powdery mildew | Erysiphe cruciferarum | Erysiphales | - | [54,64] | ||
| Wild | [62,76,86,87] | |||||
| Leveillula taurica | Erysiphales | - | [64] | |||
| Rust | Puccinia isiacae | Pucciniales | - | [88] | ||
| White rust | Albugo candida | Albuginales | Wild | [74] | ||
| Wild and pot (artificial inoculation) | [89] | |||||
| Albugo lepidii | Albuginales | Wild and growth chamber (artificial inoculation) | [90] | |||
| L. perfoliatum | Downy mildew | Hyaloperonospora lepidii-perfoliati | Peronosporales | Herbarium specimen(s) | [43] | |
| Leaf smut | Urocystis coralloides | Urocystidiales | - | [91] | ||
| Powdery mildew | Erysiphe cruciferarum | Erysiphales | - | [64] | ||
| Wild | [76] | |||||
| Rust | Puccinia aristidae | Pucciniales | - | [59] | ||
| Stunting | Verticillium dahliae | Glomerellales | Greenhouses, gardens, fields, and orchards | [92] | ||
| L. ruderale | Downy mildew | Hyaloperonospora parasitica | Peronosporales | Herbarium specimen(s) | [51] | |
| Perofascia lepidii | Peronosporales | Herbarium specimen(s) | [51,83] | |||
| Leaf smut | Urocystis coralloides | Urocystidiales | - | [91] | ||
| Powdery mildew | Erysiphe cruciferarum | Erysiphales | - | [54,64] | ||
| Wild | [76,93] | |||||
| White rust | Albugo candida | Albuginales | - | [54] | ||
| L. subulatum | Downy mildew | Perofascia lepidii | Peronosporales | Wild | [53] | |
| Portulaca | P. oleracea | Anthracnose | Colletotrichum fructicola | Glomerellales | - | [94] |
| Colletotrichum gleosporioides species complex | Glomerellales | - | [94] | |||
| Colletotrichum spinaciae | Glomerellales | Herbarium specimen(s) | [95] | |||
| Black root rot | Berkeleyomyces basicola | Microascales | Tobacco field | [96] | ||
| Basal stem rot | Rhizoctonia solani | Cantharellales | Wild | [97] | ||
| Sclerotium rolfsii | Thelephorales | - | [94] | |||
| Crown rot | Sclerotinia sclerotiorum | Helotiales | Wild | [98] | ||
| Leaf blight | Stemphylium vesicarium | Pleosporales | Onion field and growth chamber (artificial inoculation) | [99] | ||
| Leaf spot | Dichotomophthora lutea | Pleosporales | Field and pot (artificial inoculation) | [100] | ||
| Herbarium specimen(s) | [101,102] | |||||
| Phyllosticta portulacae | Botryosphaeriales | Wild | [103] | |||
| Root dieback | Globisporangium intermedium | Peronosporales | Carrot field | [104] | ||
| Globisporangium irregulare | Peronosporales | Carrot field | [104] | |||
| Stem rot | Dichotomophthora portulacae | Pleosporales | Cultivated (not specified) and pot in glasshouse (artificial inoculation) | [105] | ||
| Wild | [106,107] | |||||
| Wild and greenhouse (artificial inoculation) | [108] | |||||
| Wild and pots in growth chamber (artificial inoculation) | [109] | |||||
| Curvularia portulacae | Pleosporales | Field and greenhouse | [110] | |||
| Not specified | [111] | |||||
| Stunting | Verticillium dahliae | Glomerellales | Greenhouses, gardens, fields, and orchards | [92] | ||
| White rust | Wilsoniana portulacae | Albuginales | - | [51,77,112] | ||
| Herbarium specimen(s) | [113] | |||||
| Maize, soybean, sugar beet, and sunflower fields | [114] | |||||
| Wild | [74,98,103,115,116] | |||||
| Wild and maize and sunflower fields | [117] | |||||
| Salicornia | S. perennans (=Syn.: S. europaea) | Black lesions on leaves | Phytophthora inundata | Peronosporales | Wild | [118] |
| Rust | Uromyces salicorniae | Pucciniales | - | [54,77] | ||
| Wild | [119,120,121] | |||||
| S. perennis | Rust | Uromyces salicorniae | Pucciniales | Wild | [121] | |
| S. ramosissima | Rust | Uromyces salicorniae | Pucciniales | Wild | [121] | |
| Salsola | S. kali | Black foot | Dactylonectria macrodidyma | Hypocreales | Grapevine field | [122] |
| Powdery mildew | Leveillula cylindrospora | Erysiphales | Wild | [123] | ||
| Leveillula taurica | Erysiphales | - | [64] | |||
| Rust | Puccinia aristidae | Pucciniales | Herbarium specimen(s) | [124] | ||
| Uromyces chenopodii | Pucciniales | Wild | [66,98,125] | |||
| Uromyces salsolae | Pucciniales | Pots under greenhouse | [126] | |||
| Wild | [127] | |||||
| Stunting | Verticillium dahliae | Glomerellales | Greenhouses, gardens, fields, and orchards | [92] | ||
| S. tragus | Anthracnose | Colletotrichum salsolae | Glomerellales | Wild and greenhouse (artificial inoculation) | [128,129,130] | |
| Field | [131] | |||||
| S. tragus | Stem canker | Diaporthe eres | Diaporthales | Wild and greenhouse (artificial inoculation) | [132] | |
| Soda | S. inermis (=Syn.: Salsola soda) | Powdery mildew | Leveillula cylindrospora | Erysiphales | Wild | [76] |
| Rust | Uromyces salsolae | Pucciniales | - | [133] | ||
| Suaeda | S. maritima | Rust | Uromyces chenopodii | Pucciniales | - | [134] |
| Wild | [103,135] | |||||
| S. vera | Rust | Uromyces chenopodii | Pucciniales | - | [77,136] | |
| Wild | [66,120] |
Table 3.
Host range of pathogenic species. Only pathogenic species reported on two or more different Mediterranean halophytic host species have been considered.
Table 3.
Host range of pathogenic species. Only pathogenic species reported on two or more different Mediterranean halophytic host species have been considered.
| Pathogenic Species | Host Family | Host Species |
|---|---|---|
| Albugo candida | Brassicaceae | Cakile maritima |
| Lepidium latifolium | ||
| Lepidium ruderale | ||
| Ascochyta caulina | Amaranthaceae | Atriplex hortensis |
| Atriplex littoralis | ||
| Atriplex patula | ||
| Atriplex prostrata | ||
| Erysiphe cruciferarum | Brassicaceae | Cakile maritima |
| Lepidium latifolium | ||
| Lepidium ruderale | ||
| Lepidium perfoliatum | ||
| Hyaloperonospora parasitica | Brassicaceae | Cakile maritima |
| Lepidium ruderale | ||
| Leveillula cylindrospora | Amaranthaceae | Salsola kali |
| Soda inermis | ||
| Leveillula taurica | Amaranthaceae | Beta vulgaris subsp. maritima |
| Salsola kali | ||
| Brassicaceae | Lepidium latifolium | |
| Apiaceae | Crithmum maritimum | |
| Passalora spegazzinii | Amaranthaceae | Atriplex patula |
| Atriplex hortensis | ||
| Perofascia lepidii | Brassicaceae | Lepidium latifolium |
| Lepidium ruderale | ||
| Lepidium subulatum | ||
| Puccinia aristidae | Amaranthaceae | Salsola kali |
| Brassicaceae | Lepidium perfoliatum | |
| Stagonospora atriplicis | Amaranthaceae | Atriplex hortensis |
| Atriplex littoralis | ||
| Atriplex patula | ||
| Atriplex prostrata | ||
| Urocystis coralloides | Brassicaceae | Lepidium perfoliatum |
| Lepidium ruderale | ||
| Uromyces chenopodii | Amaranthaceae | Salsola kali |
| Suaeda maritima | ||
| Suaeda vera | ||
| Uromyces salicorniae | Amaranthaceae | Salicornia perennans |
| Salicornia perennis | ||
| Salicornia ramosissima | ||
| Uromyces salsolae | Amaranthaceae | Salsola tragus |
| Salsola kali | ||
| Soda inermis | ||
| Verticillium dahliae | Amaranthaceae | Salsola kali |
| Brassicaceae | Lepidium perfoliatum | |
| Portulacaceae | Portulaca oleracea |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Delli Compagni, E.; Pardossi, A.; Pecchia, S. Fungal and Fungal-like Diseases of Halophytes in the Mediterranean Basin: A State-of-the-Art Review. Horticulturae 2024, 10, 313. https://doi.org/10.3390/horticulturae10040313
AMA Style
Delli Compagni E, Pardossi A, Pecchia S. Fungal and Fungal-like Diseases of Halophytes in the Mediterranean Basin: A State-of-the-Art Review. Horticulturae. 2024; 10(4):313. https://doi.org/10.3390/horticulturae10040313
Chicago/Turabian StyleDelli Compagni, Emiliano, Alberto Pardossi, and Susanna Pecchia. 2024. "Fungal and Fungal-like Diseases of Halophytes in the Mediterranean Basin: A State-of-the-Art Review" Horticulturae 10, no. 4: 313. https://doi.org/10.3390/horticulturae10040313
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.