
First Molecular Phylogenetic Identification and Report of Pseudocercospora cannabina Causing Leaf Spot Disease on Cannabis sativa in Thailand
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. DNA Extraction and PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
2.4. Pathogenicity Test
3. Results
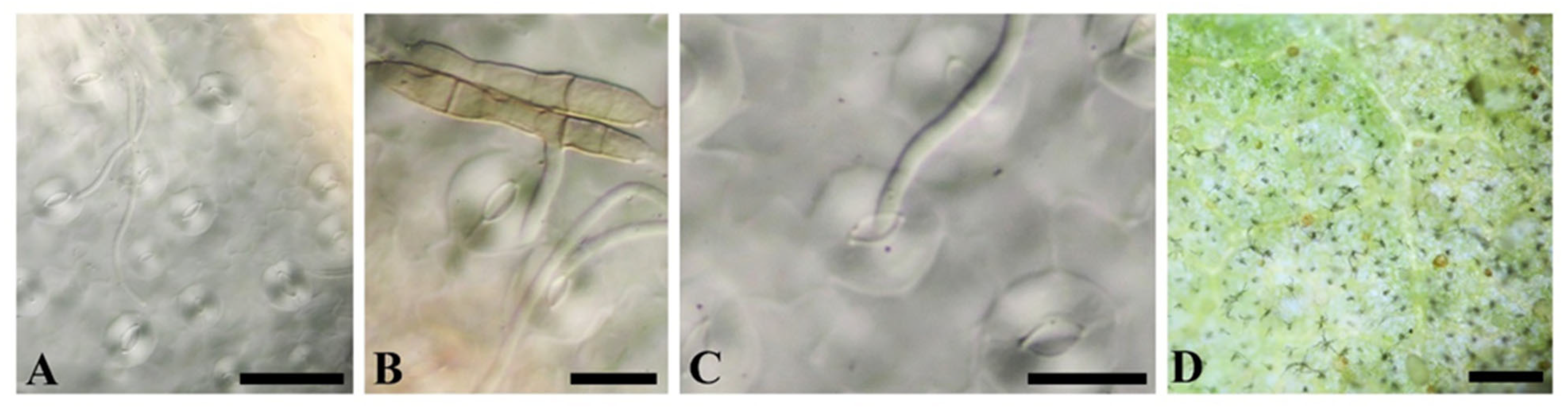
3.1. Taxonomy
Taxonomic Description
3.2. Pathogenicity Test
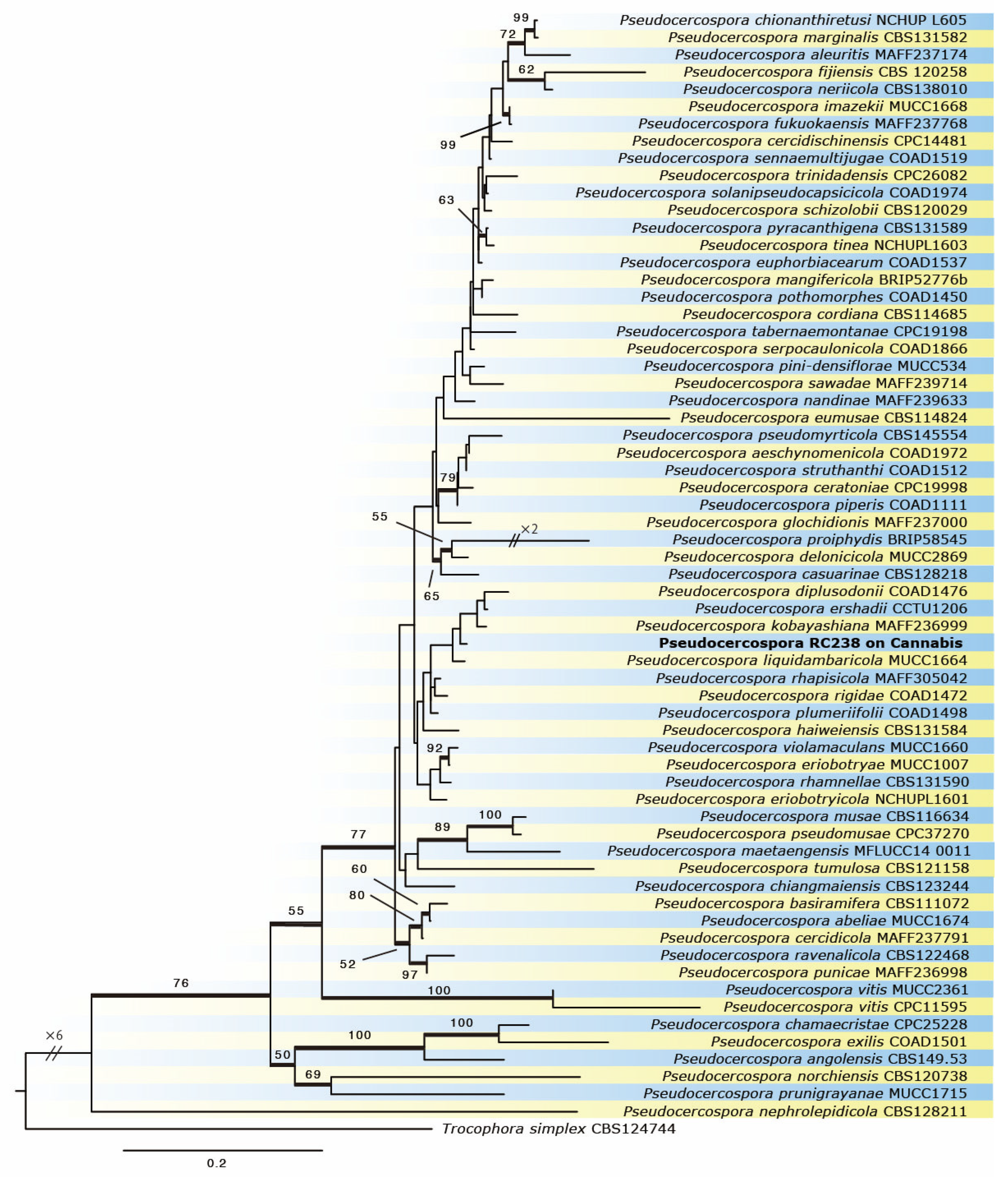
3.3. Phylogeny
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salentijn, E.M.J.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New developments in fiber hemp (Cannabis sativa L.) breeding. Ind. Crops Prod. 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Struik, P.C.; Amaducci, S.; Bullard, M.J.; Stutterheim, N.C. Agronomy of fibre hemp (Cannabis sativa L.) in Europe. Ind. Crops Prod. 2000, 11, 107–118. [Google Scholar] [CrossRef]
- Tsaliki, E.; Kalivas, A.; Jankauskiene, Z.; Irakli, M.; Cook, C.; Grigoriadis, I.; Panoras, I.; Vasilakoglou, I.; Dhima, K. Fibre and Seed Productivity of Industrial Hemp (Cannabis sativa L.) Varieties under Mediterranean Conditions. Agronomy 2021, 11, 171. [Google Scholar] [CrossRef]
- Rupasinghe, V.H.P.; Davis, A.; Kumar, S.K.; Murray, B.; Zheljazkov, V.Z. Industrial hemp (Cannabis sativa subsp. sativa) as an emerging source for value-added functional food ingredients and nutraceuticals. Molecules 2020, 25, 4078. [Google Scholar] [CrossRef]
- Gwinn, K.D.; Hansen, Z.; Kelly, H.; Ownley, B.H. Diseases of Cannabis sativa Caused by Diverse Fusarium Species. Front Agronomy 2022, 3, 796062. [Google Scholar] [CrossRef]
- Honig, L. National Hemp Report Agricultural Statistics Board Briefing, USDA. 2022. Available online: www.nass.usda.gov (accessed on 25 May 2023).
- Martin, M.A. Ethnobotanical Aspects of Cannabis in Southeast Asia. In Cannabis and Culture; Rubin, V., Ed.; Mouton Publishers: Berlin, Germany, 1975; pp. 63–76. ISBN 9027976694. [Google Scholar]
- Cerino, P.; Buonerba, C.; Cannazza, G.; D’Auria, J.; Ottoni, E.; Fulgione, A.; Di Stasio, A.; Pierri, B.; Gallo, A. A Review of Hemp as Food and Nutritional Supplement. Cannabis Cannabinoid Res. 2021, 6, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, H.N.; Zhang, Z. The use of hemp fibers as reinforcements in composites. In Biofiber Reinforcements in Composite Materials; Woodhead Publishing: Sawston, UK, 2015; pp. 86–103. ISBN 9781782421221. [Google Scholar] [CrossRef]
- Punja, Z.K. Emerging diseases of Cannabis sativa and sustainable management. Pest Manag. Sci. 2021, 9, 3857–3870. [Google Scholar] [CrossRef]
- Finley, S.J.; Javan, C.T.; Green, R.L. Bridging Disciplines: Applications of Forensic Science and Industrial Hemp. Front. Microbiol. 2022, 13, 760374. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Collyer, D.; Scott, C.; Lung, S.; Holmes, J.; Sutton, D. Pathogens and Molds Affecting Production and Quality of Cannabis sativa L. Front. Plant Sci. 2019, 10, 1120. [Google Scholar] [CrossRef] [PubMed]
- Jerushalmi, S.; Maymon, M.; Dombrovsky, A. Fungal Pathogens Affecting the Production and Quality of Medical Cannabis in Israel. Plants 2020, 9, 882. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Xu, J.; Balasuriya, A.; Hyde, K.D.; Stępień, Ł.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in fungal plant pathology: Beyond the species level. Fungal Divers. 2021, 109, 267–282. [Google Scholar] [CrossRef]
- McPartland, J.M.; Hillig, K.W. White Leaf Spot. J. Ind. Hemp 2006, 11, 43–50. [Google Scholar] [CrossRef]
- Bakro, F.; Wielgusz, K.; Bunalski, M.; Jedryczka, M. An overview of pathogen and insect threats to fibre and oilseed hemp (Cannabis sativa L.) and methods for their biocontrol. In Proceedings of the Meeting 17th IOBC Working Group Meeting on Integrated Control in Oilseeds (ICOC), International Organization for Biological and Integrated Control of Noxious Animals and Plants, West Palearctic Regional Section (IOBC-WPRS), Zagreb, Croatia, 18–20 September 2018; Volume 136, pp. 9–20. [Google Scholar]
- Santo, D. Australian Industrial Hemp Industry Pesticide Review and Evaluation Program. AgriFutures Australia. Publication No. 22-060. 2022. Available online: https://agrifutures.com.au/wp-content/uploads/2022/06/22-060.pdf (accessed on 25 May 2023).
- Rodriguez, G.; Kibler, A.; Campbell, P.; Punja, Z.K. Fungal diseases of Cannabis sativa in British Columbia, Canada. In Proceedings of the American Phytopathological Society Annual Meeting, Pasadena, CA, USA, 1–5 August 2015. Poster 529-P. [Google Scholar]
- Ward-Gauthier, N.; Beale, J.; Amsden, B.; Dixon, E. Greenhouse hemp in Kentucky exhibits many common diseases. In Proceedings of the American Phytopathological Society Annual Meeting, Pasadena, CA, USA, 1–5 August 2015. Poster 502-P. [Google Scholar]
- Crous, P.W.; Braun, U.; Hunter, G.C.; Wingfield, M.J.; Verkley, G.J.M.; Shin, H.-D.; Nakashima, C.; Groenewald, J.Z. Phylogenetic lineages in Pseudocercospora. Stud. Mycol. 2013, 75, 37–114. [Google Scholar] [CrossRef] [PubMed]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, E.B.G.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Chen, Q.; Bakhshi, M.; Balci, Y.; Broders, K.; Cheewangkoon, R.; Chen, S.; Fan, X.; Gramaje, D.; Halleen, F.; Jung, M.H.; et al. Genera of phytopathogenic fungi: GOPHY 4. Stud. Mycol. 2022, 101, 417–564. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, C.; Motohashi, K.; Chen, C.Y.; Groenewald, J.Z.; Crous, P.W. Species diversity of Pseudocercospora from Far East Asia. Mycol. Prog. 2016, 15, 1093–1117. [Google Scholar] [CrossRef]
- Aslake, M.; Sintayehu, A.; Alem, T.; Abera, M. Distribution and Status of Citrus Leaf and Fruit Spot (Pseudocercospora angolensis) Disease in North Western Ethiopia. Open J. Ecol. 2022, 12, 733–741. [Google Scholar] [CrossRef]
- Silva, K.J.D.E.; De Souza, E.A.; Sartorato, A.; De Souza, F. Pathogenic Variability of Isolates of Pseudocercospora griseola, the Cause of Common Bean Angular Leaf Spot, and its Implications for Resistance Breeding. J. Phytopathol. 2008, 156, 602–606. [Google Scholar] [CrossRef]
- Evans, H.C. The genus Mycosphaerella and its anamorphs Cercoseptoria, Dothistroma and Lecanosticta on pines. Mycol. Pap. 1984, 153, 1–102. [Google Scholar]
- Braun, U.; Nakashima, C.; Crous, P.W. Cercosporoid fungi (Mycosphaerellaceae) 1. Species on other fungi, Pteridophyta and Gymnospermae. IMA Fungus 2013, 4, 265–345. [Google Scholar] [CrossRef]
- Videira, S.I.R.; Groenewald, J.Z.; Nakashima, C.; Braun, U.; Barreto, R.W.; de Wit, P.J.G.M.; Crous, P.W. Mycosphaerellaceae—Chaos or clarity? Stud. Mycol. 2017, 87, 257–421. [Google Scholar] [CrossRef] [PubMed]
- To-Anun, C.; Hidayat, I.; Meeboon, J. Genus Cercospora in Thailand: Taxonomy and Phylogeny (with a dichotomous key to species). Plant Pathol. Quar. 2011, 1, 11–87. [Google Scholar] [CrossRef]
- White, T.J.; Burns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships Among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1998, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Sautua, F.J.; Doyle, V.P.; Price, P.P.; Porfiri, A.; Fernandez, P.; Scandiani, M.M.; Carmona, M.A. Fungicide resistance in Cercospora species causing Cercospora leaf blight and purple seed stain of soybean in Argentina. Plant Pathol. 2020, 69, 1678–1694. [Google Scholar] [CrossRef]
- Wakefield, E.M. Fungi Exotici: XXIII. Bull. Misc. Inf. R. Bot. Gard. Kew. 1917, 1917, 308–314. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y.; Castlebury, L.A. United States National Fungus Collections Fungus-Host Dataset. Ag Data Commons. 2021. Available online: https://data.nal.usda.gov/dataset/united-states-national-fungus-collections-fungus-host-dataset (accessed on 13 November 2022).
- Chupp, C. A monograph of the Fungus genus Cercospora. by the Author Plant Pathology Department, Cornell University, Ithaca, N. Y. AIBS Bull. 1954, 4, 11. [Google Scholar] [CrossRef]
- Crous, P.W.; Braun, U. Mycosphaerella and its Anamorphs 1: Names Published in Cercospora and Passalora; CBS Biodiversity Series 1; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2003; ISBN 90-70351-49-8. [Google Scholar]
- Chen, Y.J.; Jayawardena, R.S.; Bhunjun, C.S.; Harishchandra, D.L.; Hyde, K.D. Pseudocercospora dypsidis sp. Nov. (Mycosphaerellaceae) on Dypsis lutescens leaves in Thailand. Phytotaxa 2020, 474, 218–234. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Culture Collection Number | ITS | act | tef | rpb2 |
|---|---|---|---|---|---|
| Pseudocercospora abeliae | MUCC1674 * | LC599330 | LC599407 | LC599448 | LC599587 |
| Ps. aeschynomenicola | COAD 1972 * | KT290146 | KT313501 | KT290200 | NA |
| Ps. aleuritis | MAFF237174 * | LC599331 | LC599408 | LC599449 | LC599588 |
| Ps. angolensis | CBS 149.53 * | JQ324975 | JQ325011 | JQ324988 | NA |
| Ps. basiramifera | CBS 111072 * | GU269661 | GU320368 | DQ211677 | NA |
| Ps. cannabina | SDBR-CMU372 * | OR101673 | OR344083 | NA | OR344085 |
| Ps. cannabina | SDBR-CMU373 | OR101674 | OR344084 | NA | OR344086 |
| Ps. casuarinae | CBS 128218 * | HQ599603 | LC599413 | LC599454 | NA |
| Ps. ceratoniae | CBS 147386 = CPC19998 * | LC599335 | LC599414 | LC599455 | LC599592 |
| Ps. cercidicola | MAFF237791 * | GU269671 | GU320377 | GU384388 | KX462618 |
| Ps. cercidis-chinensis | CBS 132109 = CPC14481 * | GU269670 | GU320376 | GU384387 | LC599593 |
| Ps. chamaecristae | CPC 25228 = COAD 1973 * | KT290147 | KT313502 | KT290201 | NA |
| Ps. chiangmaiensis | CBS 123244 * | EU882113 | KF903544 | KF903177 | NA |
| Ps. chionanthi-retusi | NCHUP L605 * | KX462585 | KX462552 | KX462671 | KX462620 |
| Ps. cordiana | CBS 114685 * | AF362054 | GU320387 | GU384398 | NA |
| Ps. delonicicola | MUCC2869 * | LC599341 | LC599421 | LC599463 | LC599601 |
| Ps. diplusodonii | CPC 25179 = COAD 1476 * | KT290135 | KT313490 | KT290189 | NA |
| Ps. eriobotryae | MUCC1007 * | KX462589 | KX462557 | KX462676 | KX462628 |
| Ps. eriobotryicola | NCHUP L1601 * | KX462590 | KX462558 | KX462677 | KX462629 |
| Ps. ershadii | CBS 136114 = CCTU1206 * | KM452867 | KM452844 | KM452889 | MN786459 |
| Ps. eumusae | CBS 114824 * | EU514238 | LFZN0100 0053 | LFZN0100 0037 | NA |
| Ps. euphorbiacearum | COAD 1537 * | KT290145 | KT313500 | KT290199 | NA |
| Ps. exilis | COAD 1501* | KT290139 | KT313494 | KT290193 | NA |
| Ps. fijiensis | CBS 120258 = CIRAD 86 * | EU514248 | NW006921533 | NW006921532 | NW006921535 |
| Ps. fukuokaensis | MAFF237768 * | GU269714 | GU320418 | GU384430 | KX462632 |
| Ps. glochidionis | MAFF237000 * | LC599348 | LC599428 | LC599470 | LC599608 |
| Ps. haiweiensis | CBS 131584 * | GU269803 | GU320506 | GU384514 | KX462634 |
| Ps. imazekii | MUCC1668 * | KX462596 | KX462564 | KX462683 | KX462638 |
| Ps. kobayashiana | MAFF236999 * | LC511998 | LC512004 | LC515780 | LC515791 |
| Ps. liquidambaricola | MUCC1664 * | LC599352 | LC599432 | LC599474 | LC599611 |
| Ps. maetaengensis | MFLUCC 14-0011 * | GU188048 | NA | NA | NA |
| Ps. mangifericola | BRIP 52776b * | GU188048 | NA | NA | NA |
| Ps. marginalis | CBS 131582 * | GU269794 | GU320495 | GU384504 | NA |
| Ps. musae | CBS 116634 * | GU269747 | GU320449 | GU384459 | NA |
| Ps. nandinae | MAFF239633 * | KX462600 | KX462568 | KX462687 | KX462645 |
| Ps. neriicola | CBS 138010 * | KJ869165 | KJ869231 | KJ869240 | KX462647 |
| Ps. norchiensis | CBS 120738 * | EF394859 | GU320455 | GU384464 | KX462648 |
| Ps. pini-densiflorae | MUCC534 * | LC599354 | LC599434 | LC599478 | LC599615 |
| Ps. piperis | COAD 1111 | JX875062 | NA | JX896123 | NA |
| Ps. plumeriifolii | COAD 1498 * | KT290138 | KT313493 | KT290192 | NA |
| Ps. pothomorphes | COAD 1450 * | KT290131 | KT313486 | KT290185 | NA |
| Ps. proiphydis | BRIP58545 * | KM055430 | NA | KM055437 | NA |
| Ps. pruni-grayanae | MUCC1715 * | LC599356 | NA | LC599481 | LC599618 |
| Ps. pseudomusae | CBS 147147 = CPC37270* | MW063423 | MW070772 | MW071091 | MW070919 |
| Ps. pseudomyrticola | CBS 145554 * | MK876405 | MK876461 | MK876499 | MK876490 |
| Ps. punicae | MAFF236998 | KX462606 | KX462573 | KX462692 | KX462655 |
| Ps. pyracanthigena | CBS 131589 * | GU269766 | GU320469 | GU384478 | NA |
| Ps. ravenalicola | CBS 122468 * | GU269810 | GU320513 | GU384521 | NA |
| Ps. rhamnellae | CBS 131590 * | GU269795 | GU320496 | GU384505 | NA |
| Ps. rhapisicola | MAFF305042 * | LC599357 | LC599436 | LC599483 | LC599620 |
| Ps. rigidae | COAD 1472 * | KT290134 | KT313489 | KT290188 | NA |
| Ps. sawadae | MAFF239714 | LC599359 | LC599438 | LC599485 | LC599622 |
| Ps. schizolobii | CBS 120029 * | KF251322 | KF253628 | KF253269 | NA |
| Ps. sennae-multijugae | COAD 1519 * | KT290142 | KT313497 | KT290196 | NA |
| Ps. serpocaulonicola | COAD 1866 * | KT037525 | KT037607 | KT037485 | NA |
| Ps. solani-pseudocapsicicola | COAD 1974 * | KT290148 | KT313503 | KT290202 | NA |
| Ps. struthanthi | COAD 1512 * | KT290141 | KT313496 | KT290195 | NA |
| Ps. tabernaemontanae | CPC 19198 * | LC599363 | LC599442 | NA | LC599625 |
| Ps. tinea | NCHUP L1603 * | KX462608 | KX462577 | KX462696 | KX462660 |
| Ps. trinidadensis | CPC 26082 = COAD1756 * | KT290157 | NA | KT290210 | NA |
| Ps. tumulosa | CBS 121158 * | DQ530217 | NA | NA | NA |
| Ps. violamaculans | MUCC1660 * | KX462610 | KX462579 | KX462698 | KX462662 |
| Ps. vitis | CPC 11595 | GU269829 | GU320533 | GU384541 | KX462663 |
| Ps. vitis | CBS 128211 * | LC599366 | LC599445 | LC599492 | LC599627 |
| Trochophora simplex | CBS 124744 | GU269872 | GU320568 | GU384580 | KX462666 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harishchandra, D.L.; Haituk, S.; Withee, P.; Tamakaew, N.; Nokum, N.; Kanchanomai, C.; Pusadee, T.; Nakashima, C.; Cheewangkoon, R. First Molecular Phylogenetic Identification and Report of Pseudocercospora cannabina Causing Leaf Spot Disease on Cannabis sativa in Thailand. Horticulturae 2023, 9, 1261. https://doi.org/10.3390/horticulturae9121261
Harishchandra DL, Haituk S, Withee P, Tamakaew N, Nokum N, Kanchanomai C, Pusadee T, Nakashima C, Cheewangkoon R. First Molecular Phylogenetic Identification and Report of Pseudocercospora cannabina Causing Leaf Spot Disease on Cannabis sativa in Thailand. Horticulturae. 2023; 9(12):1261. https://doi.org/10.3390/horticulturae9121261
Chicago/Turabian StyleHarishchandra, Dulanjalee Lakmali, Sukanya Haituk, Patchareeya Withee, Nisachon Tamakaew, Nittaya Nokum, Chaorai Kanchanomai, Tonapha Pusadee, Chiharu Nakashima, and Ratchadawan Cheewangkoon. 2023. "First Molecular Phylogenetic Identification and Report of Pseudocercospora cannabina Causing Leaf Spot Disease on Cannabis sativa in Thailand" Horticulturae 9, no. 12: 1261. https://doi.org/10.3390/horticulturae9121261