
Soil Conditioning and Neighbor Identity Influence on Cycas Seedling Performance


1
Cycad Specialist Group, International Union for Conservation of Nature Species Survival Commission, Rue Mauverney 28, 1196 Gland, Switzerland
2
Bagong Kaalaman Botanikal Institute, 15 Rizal Street, Barangay Malabañas, Angeles City 2009, Philippines
Ecologies 2023, 4(4), 731-748; https://doi.org/10.3390/ecologies4040048
Submission received: 9 September 2023
/
Revised: 18 November 2023
/
Accepted: 20 November 2023
/
Published: 23 November 2023
Abstract
:Translocation of endangered plant species and facilitating in situ regeneration require knowledge of the factors that define suitable habitat characteristics. Four approaches were employed to define how antecedent and contemporary plant identity influenced Cycas seedling growth and survival in Guam, Yap, and the Philippines. Cycas micronesica K.D. Hill or Cycas nitida K.D. Hill & A. Lindstr. seedlings growing beneath the canopy of the maternal parent tree reached 100% mortality in 4 to 8 year. Cycas micronesica seedlings planted beneath a conspecific male adult tree reached 68% mortality in two years while seedlings planted away from a conspecific individual exhibited 100% survival. Cycas micronesica seedlings planted beneath monostands of invasive Leucaena leucocephala (Lam.) de Wit or Vitex parviflora Juss. reached 40% mortality in two years while seedlings planted in adjacent native forest cover exhibited 100% survival. Cycas micronesica seedlings planted in soil conditioned for 9 months by conspecifics, heterospecific native species, Carica papaya L., L. leucocephala, or V. parviflora exhibited the greatest growth in the soils conditioned by heterospecific natives and the least growth in the soils conditioned by the three invasive species. The seedlings growing in soil conditioned by conspecifics exhibited the most coralloid roots and the greatest shoot:root ratio. These studies confirmed that the identity of antecedent and competitive plants strongly influenced the longevity and growth of Cycas seedlings. Conservationists may improve outcomes by avoiding degraded forests that have experienced invasive plant species when positioning translocation projects. Planting Cycas seedlings in biodiverse native forest but away from a conspecific individual may lead to the greatest success in population recovery.
1. Introduction
Plants alter many factors of the soils in which they grow, and some of these factors can influence the performance of other plants that grow at a later date in the same soils. The extent of this effect varies with the identity of the plants before and during seedling growth. These time-lag issues are discussed extensively in the considerable literature described as land-use legacy, soil legacy effects, allelopathy, and invasive plant species [1,2,3,4,5,6,7,8]. For controlled studies, soil conditioning is often accomplished experimentally by growing the initial plants in container cultures, then using the container medium for growing subsequent plants [9,10]. For ecological studies, these issues are often studied as part of the underlying factors that support the Janzen–Connell hypothesis [11,12], where seedling proximity to and competition with conspecific plants may negatively influence growth and survival.
Knowledge and appreciation of how historical plant cover influences contemporary plant performance may inform conservation decisions of imperiled plant species. This may be especially true for non-recruiting species characterized by persisting mature individuals with no active regeneration for years, species that Janzen refers to as the “living dead” [13,14]. The Cycas micronesica populations on the island of Guam are currently among the living dead [15]. This status has been created by the 2003 invasion of Guam by the specialist armored scale Aulacaspis yasumatsui Takagi [16,17]. The ubiquitous A. yasumatsui pressures have caused failures in the recruitment of new C. micronesica seedlings for two decades. A conservative estimate indicated that 70 year of demographic depth had been culled from these forests by 2020 [18]. Clearly, one critical component of a conservation plan designed to enable C. micronesica recovery involves assisting the re-establishment of regeneration and recruitment. Yet, research that directly informs this action is lacking.
The objectives of this investigation were to use four observational and experimental approaches to determine how soils that were conditioned by conspecific, sympatric native heterospecific, or invasive tree species influenced seedling growth and survival of two Cycas species in Guam, Yap, and the Philippines. The results will be of value to conservation planners and practitioners to make more informed restoration and species recovery decisions.
2. Materials and Methods
Four islands in the western Pacific were used to more fully understand plant identity factors involved in Cycas seedling survival (Figure 1).
2.1. Preliminary Study
Resilience of C. micronesica plants following damage during the 1997 Typhoon Paka was observed from December 1997 until June 1999 [19]. Although the primary dataset focused on the recovery of mature trees that were damaged in northern Guam, we collected ancillary data from 50 female trees with high density seedlings beneath their canopies (Figure 2). The range in the initial count was 19 to 33 seedlings per tree and the total number of seedlings monitored was 1296. The number of seedlings beneath these trees was counted in December 1997, December 1998, and June 1999. The data are presented as mean ± standard deviation.
2.2. Janzen–Connell Seedling Mortality
Fifteen female C. micronesica trees with abundant one-leaf seedlings beneath the canopy were identified in northern Guam in February 2001. The karst soils formed in slope alluvium, loess, and residuum overlying limestone. Trees with older seedlings were not included in order to reduce the noise caused by the initial presence of older half-siblings. The range was 16 to 24 seedlings per tree and the total number of seedlings monitored was 301. The persistent seed sclerotesta of every seedling was painted with blue paint to distinguish the original population from all additional regeneration during the observation period. The trees were observed annually until all of the marked seedlings had died. At each annual visit, the number of remaining seedlings from the original 2001 date were counted, then the total number of seedlings beneath the canopy of each tree was counted. This latter number fluctuated and was influenced by the periodic addition of new seeds that germinated. The study was terminated in February 2005 when all of the original seedlings had died.
Ten female C. micronesica trees were identified for observation in Yap in 2010. The soils were formed in residuum derived from green, chlorite, and talc schist. The range was 22 to 32 seedlings per tree and the total number of seedlings monitored was 278. The plants were observed in 2011, 2015, 2016, and 2018 when all of the original seedlings had died. Data were collected as described above.
Ten female C. nitida trees were identified for observation in Matnog, Sorsogon, Philippines in 2011. The sandy soils were formed in water-deposited sand derived from coral reef formations. The range was 15 to 21 seedlings per tree and the total number of seedlings monitored was 179. The plants were observed in 2012, 2013, 2014, and 2016 when all of the original seedlings had died. Data were collected as described above.
Twelve female C. nitida trees were identified for observation in Calbayog, Samar, Philippines in 2013. The substrate was a halomorphic soil formed due to the salination of parent material. The range was 15 to 24 seedlings per tree and the total number of seedlings monitored was 233. The plants were observed in 2014, 2016, 2017, and 2018 when all of the original seedlings had died. Data were collected as described above.
For each of the four sites and observation dates, the number of living original seedlings and the total number of standing seedlings were presented as mean ± standard deviation. This study included a total of 991 newly-emerged Cycas seedlings that were observed until death.
2.3. Distance-Dependent Growth and Mortality
Mature seeds from a single C. micronesica female tree in northwest Guam were harvested on 7 March 2002 (n = 115). The karst soils formed in slope alluvium, loess, and residuum overlying limestone. The integuments were cleaned, then the seeds were stored in ambient conditions for nine months for embryo maturation. Seeds were sown in perlite beds and watered every other day until germination was observed. On 14 March 2003, 64 synchronized germinated seeds were removed from the germination bed and taken to the same habitat as the parent tree to be placed within in situ soils before the radicles had extended.
Eight mature male C. micronesica trees were identified to receive germinating seedlings as the “near” treatment, and a paired site for each tree was identified 10–15 m away to receive seedlings as the “away” treatment. The male C. micronesica trees were 2.8–3.6 m in stem height, and were used instead of female trees to ensure no new seedlings would be recruited during the course of the study. Sympatric native canopy species in this habitat included Ficus prolixa G.Forst., Premna obtusifolia R.Br., and Ochrosia oppositifolia (Lam.) K.Schum. Each “away” location was positioned where mature individuals of all three of these trees contributed to the litter layer. Four germinated seeds were planted at each of the 16 locations.
The methods were designed to study distance effects but exclude density effects. Therefore, the four seeds planted beneath each C. micronesica tree were planted at the cardinal positions at the edge of the canopy, which was defined in this pachycaulous tree by the length of the petiole and rachis of the compound leaves. Each seedling was therefore in soils that were conditioned by the mature conspecific tree, but not in competition with half-siblings. When the precise location for the “away” site was identified, the four seeds were planted with the same distance and orientation as the paired “near” site.
A 0.75-m line quantum sensor (EMS-7, PP Systems, Amesbury, MA, USA) was employed to quantify the incident light for each of the 16 locations. The photosynthetically active radiation (PAR) transmission at the location was measured from 1100 h to 1300 h on several clear days. The sensor was positioned directly above each of the four seeds and the mean PAR was designated as the PAR for each site. For the “near” sites, half of the sensor was beneath the conspecific canopy and half was outside of the canopy.
The original objectives were to observe the 64 seedlings every 6 months until the study plants reached 100% mortality. However, termination of the investigation was required in June 2005 because A. yasumatsui immigrated into the study site and infested the surviving experimental plants. The year of mortality was recorded for each plant that died prior to June 2005. The stem height, stem diameter, leaf number, and maximum leaf length were measured for each surviving seedling in June 2005. The mortality data were presented as a function of time. The plant size metrics for surviving seedlings were averaged to provide one number for each of the 16 sites. The plant size metrics were subjected to t-test to compare the eight “near” and “away” sites.
2.4. Invasive Plant Legacies
Forest microsites that had been conditioned by invasive canopy tree species were identified in northern Guam for high density monostands of Leucaena leucocephala and Vitex parviflora fragments (Figure 3). The invaded sites with nearby C. micronesica trees were selected for this study. Mature seeds from a C. micronesica female tree growing within the native tree cover adjacent to the L. leucocephala site were harvested on 10 March 2002 (n = 107). Mature seeds from a C. micronesica female tree growing within the native tree cover adjacent to the V. parviflora site were harvested on 12 March 2002 (n = 118). Seed preparation and germination treatments were as described in Section 2.3.
On 16 March 2003, 72 synchronized germinated seeds were taken to the L. leucocephala habitat to be planted among three forms of plant cover. Six mature male C. micronesica trees were located nearby the female source tree but close to the ecotone that separated the native plant cover and the L. leucocephala monostand. Four seeds were planted under each C. micronesica tree, in six sites in the native forest cover 10–15 m away from each C. micronesica tree, and in six sites within the adjacent L. leucocephala cover as described in Section 2.3. The male C. micronesica trees were 3.1–3.7 m in stem height. This procedure provided six replications of four seedlings each for each of the three soil conditioning treatments. Sympatric native canopy species in this habitat included Aglaia mariannensis Merr., Morinda citrifolia L., and O. oppositifolia. Each native forest cover location was positioned where mature individuals of all three of these trees contributed to the litter layer.
On 18 March 2003, 72 synchronized germinated seeds were taken to the V. parviflora habitat. Treatments conformed to those described for the L. leucocephala study. The male C. micronesica trees were 2.4–3.3 m in stem height. Sympatric native canopy species in this habitat included A. mariannensis, Meiogyne cylindrocarpa (Burck) Heusden, and O. oppositifolia. Each native forest cover location was positioned where mature individuals of all three of these trees contributed to the litter layer.
The line quantum sensor was employed to quantify the incident light for each of the 18 locations for the two invasive tree studies. The 72 seedlings in each study were observed to determine mortality every January and June until 2005 when A. yasumatsui invaded the study sites in July and termination of the investigation was required. Seedling data were collected as described in Section 2.3. For each study, the plant size metrics were subjected to ANOVA. Means separation among levels of significant factors was conducted with Tukey’s honestly significant difference as pairwise comparisons. The proportion of surviving seedlings at each of the 18 sites did not conform to parametric test requirements. The non-parametric Kruskal–Wallis H test was used to compare the proportion of survivors for the three plant cover types for each of the two studies. For significant response variables, means were separated using a post-hoc Dunn–Bonferroni test.
2.5. Container Soil Conditioning Study
Traditional soil conditioning treatments were used to determine subsequent C. micronesica seedling growth in a container study conducted in Angeles City, Philippines. The experimental seeds were derived from hand-pollinated strobili of garden-grown C. micronesica plants in Angeles City. The pollen parent was from a northern Guam locality, and the ovulate parent was from a southern Guam locality. Therefore, these seedlings were of no conservation value due to the absence of pure pedigree linked to a single area of occupancy. Extreme genetic structuring among Guam’s habitats [20] indicates plants grown for conserving Guam’s genes and eventual reintroduction to Guam must be derived from parents obtained from the same area of occupancy.
The soil was harvested from a healthy forest fragment containing Gmelina arborea Roxb and Pterocarpus indicus Willd. canopy cover on 9 May 2019. The coarse loamy-textured Entisol was nutrient deficient with excessive hydraulic conductivity. The litter layer was harvested with soils to a 15 cm depth then homogenized. The living soil was placed into 1-L containers for Phase 1 of the study during which the soils were conditioned by five different plant types.
Phase 1 of the investigation was initiated on 10 May 2019. Seeds for conspecific seedlings were from hand-pollinated strobili from the C. micronesica plants in Angeles City. Seeds for M. citrifolia, A. mariannensis, V. parviflora, L. leucocephala, and Carica papaya were collected from native and naturalized plants on a commercial farm in northern Guam. Seeds for Tabernaemontana rotensis (Kaneh.) B.C. Stone were collected from mature ex situ trees at the experimental site. These trees originated from northern Guam. Each 1-L container received seedlings of one of five species identities: (1) three seedlings of C. micronesica; (2) heterospecific native plants with one M. citrifolia, one A. mariannensis, and one T. rotensis seedling; (3) three seedlings of V. parviflora; (4) three seedlings of L. leucocephala; (5) three seedlings of C. papaya L. The three invasive species and three heterospecific native species commonly occur in C. micronesica habitats in northern Guam. The heterospecific seedlings were growing in community pots and were 10–15 cm tall when transplanted into the 1-L pots.
The conditioning plants were grown under 50% shade cloth on raised nursery benches for three months. No fertilizer was added but the nursery was irrigated as needed. The soil from each of the five conditioning treatments was removed from the containers and seedling roots were rinsed, then the soil was homogenized for each of the five plant types. The 1-L pots were re-filled with each of the five soils, then seedlings with the same identities were planted into the soils for a second conditioning phase. This was begun on 11 August 2019 and the plants were grown for three months. The procedures were repeated on 14 November 2019 for a third conditioning phase. The third and final conditioning cycle was terminated on 5 February 2020. Therefore, the conditioned soils for Phase 1 of the investigation were influenced by a total of 9 months of seedling growth provided in three cycles.
The C. micronesica seeds that provided the experimental units were harvested from the megastrobilus in March 2019. The integuments were removed from the sclerotesta of the 74 seeds, then they were stored in ambient conditions for embryo maturation. The seeds were germinated by sowing in perlite beds in December 2019. There were six replications and five soil conditioning treatments, so 30 synchronized germinated seeds were selected as the experimental units to begin Phase II of the investigation on 6 February 2020.
The containers for Phase II were square air-root-pruning containers with a 15 cm top diameter, 11 cm bottom diameter, and 13 cm height (Lacebark Inc., Stillwater, OK, USA). Six containers were filled with each of the five conditioned soils from the Phase I treatments. One germinated C. micronesica seed was placed in each of the 30 containers. The plants were grown under 50% shade cloth on raised nursery benches. They were irrigated as needed, usually once per week initially and every other day in the final year.
The study was terminated on 12–18 February 2023. Stem height, stem diameter, and longest leaf length were measured for each of the three-year-old experimental units, then the number of living and dead leaves were counted. Soils were rinsed from the root system, and the plant was separated into fine roots, structural roots, coralloid roots, leaves, and stems. Tissue was dried for 48 h in a forced draft oven at 75 °C, then weighed. In addition to the dry weight of each tissue type, total plant dry weight was calculated by summing all tissue dry weights and the shoot:root ratio was calculated. The response variables were subjected to ANOVA. Means separation among levels of significant factors was conducted with Tukey’s honestly significant difference for pairwise comparisons.
3. Results
3.1. Direct Metrics
General population-level observations of C. micronesica in northern Guam revealed the rapid death of small seedlings located directly beneath the maternal parent trees. About 50% of the original seedlings were dead within 12 months, and 60% were dead within 18 months (Figure 4). These preliminary observations confirmed the speed with which in situ seedlings may die due to distance–density effects in congruity with Janzen–Connell model predictions.
3.2. Janzen–Connell Seedling Mortality
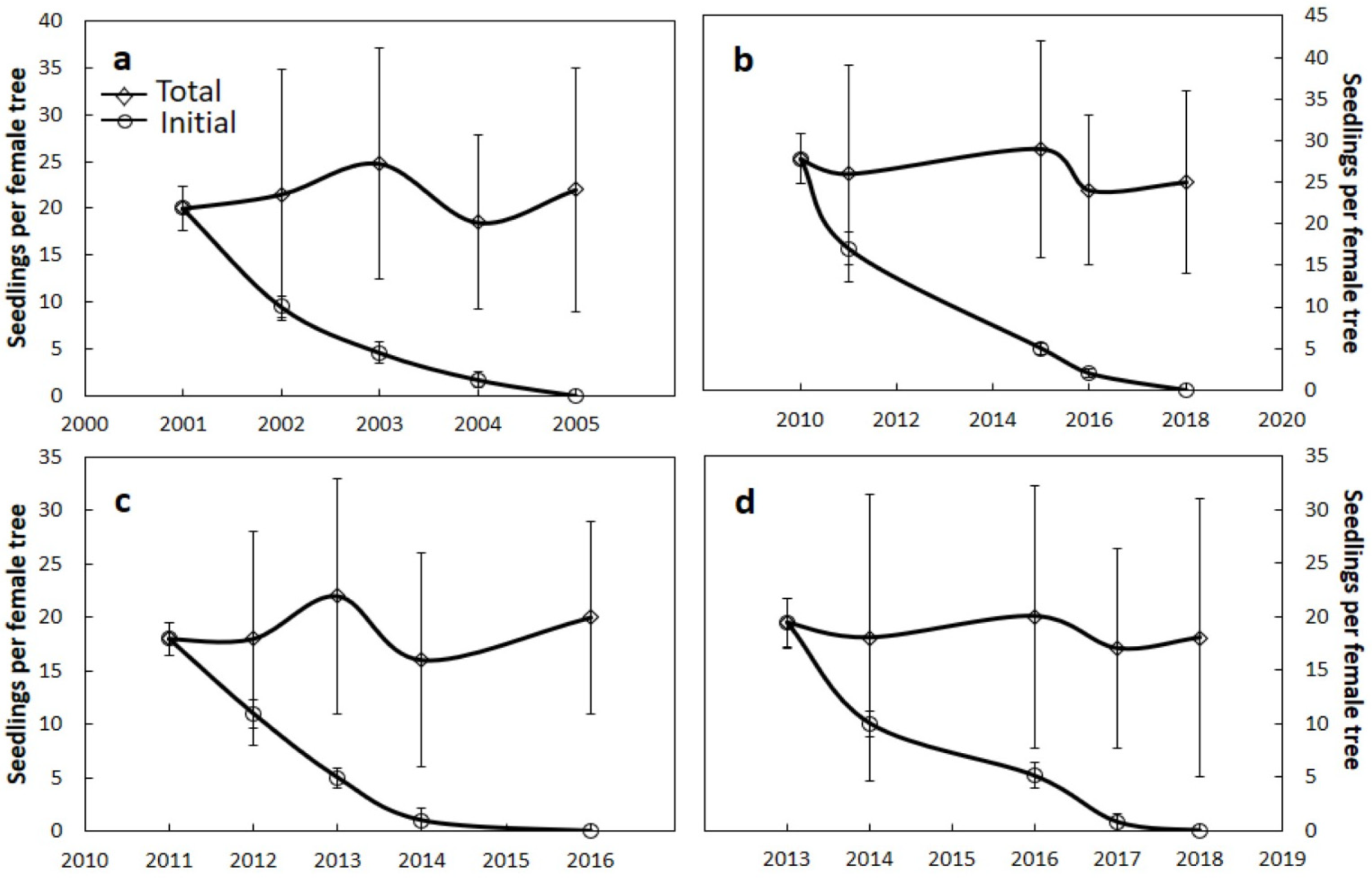
The population of young one-leaf seedlings beneath female trees exhibited the most rapid mortality for C. micronesica in Guam and C. nitida in Sorsogon and Samar, with 50% mortality occurring in 1 to 2 years and 100% mortality occurring in 4 to 5 years (Figure 5a,c,d). The seedling attrition was slower for C. micronesica in Yap, with 100% mortality occurring in 8 years (Figure 5b).
The total seedling population beneath these female Cycas trees exhibited an undulating pattern based on the mortality of pre-existing seedlings and addition of new germinating seedlings resulting from fresh seed additions. As a result, the mean and standard deviation for the total seedling population data fluctuated among the observation dates (Figure 5). The C. nitida trees maintained a seedling population of 18 to 22, the Guam C. micronesica trees maintained a population of 18 to 25, and the Yap C. micronesica trees maintained a population of 24 to 29 seedlings. These results corroborated the preliminary results in Section 3.1 by revealing the speed with which in situ Cycas seedlings die when they germinate beneath the canopy of the maternal parent tree.
3.3. Distance-Dependent Growth and Mortality
Seedlings growing beneath mature conspecific trees experienced a mean PAR of 322 µmol·m−2·s−1 on clear days, with incident light ranging from 11–19% of sunlight. The “away” seedlings experienced a mean PAR of 336 µmol·m−2·s−1 on clear days, with incident light ranging from 12–20% of sunlight.
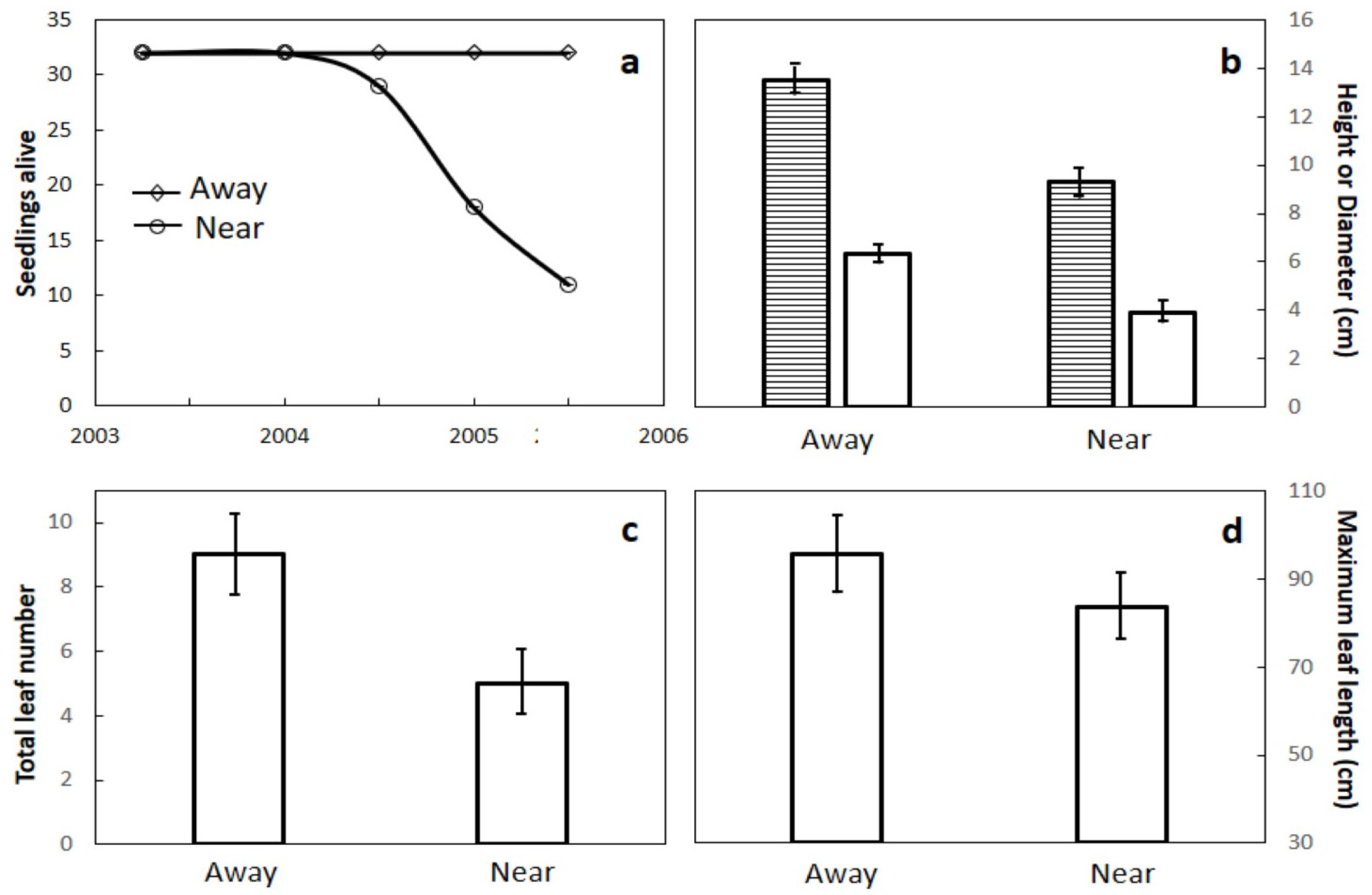
Seedlings planted beneath a mature conspecific male tree in March 2003 began to die by 18 months, and reached 68% mortality by June 2005 when the study had to be terminated due to A. yasumatsui infestations (Figure 6a). None of the seedlings that were growing away from a conspecific individual died during the course of the investigation. The “away” plants were 44% greater than “near” plants in stem height (Figure 5b; t = 8.10, p < 0.001) and 62% greater in stem diameter (Figure 6b; t = 6.67, p < 0.001). The “away” plants also exhibited 88% more leaves per seedling than “near” plants (Figure 6c; t = 5.01, p < 0.001) and 14% greater maximum leaf length than “near” plants (Figure 6d; t = 2.85, p = 0.006). There was no leaf mortality in the “away” plants, but a mean of two leaves on each of the “near” plants were dead at the end of the investigation.
These results revealed that rapid in situ seedling mortality was a function of proximity to a conspecific adult individual, and if the seeds germinated at a distance from a conspecific individual, the mortality was reduced and growth was increased. The ubiquity of A. yasumatsui throughout Guam since 2005 has removed the possibility of repeating this investigation.
3.4. Invasive Plant Legacies
The ranges in incident light at the seedling leaf level were substantial with considerable overlap occurring among the various canopy cover categories. The C. micronesica seedlings growing beneath mature conspecific trees experienced a mean PAR of 316 µmol·m−2·s−1 on clear days, with incident light ranging from 10–21% of sunlight. The seedlings growing beneath conspecific native tree cover experienced a mean PAR of 323 µmol·m−2·s−1 on clear days, with incident light ranging from 12–20% of sunlight. The seedlings growing beneath L. leucocephala tree cover experienced a mean PAR of 313 µmol·m−2·s−1 on clear days, with incident light ranging from 10–21% of sunlight. The seedlings growing beneath conspecific native tree cover experienced a mean PAR of 322 µmol·m−2·s−1 on clear days, with incident light ranging from 11–19% of sunlight.
The two-year-old C. micronesica seedlings growing under conspecific adults or L. leucocephala cover exhibited stems that were 76% of the height and 67% of the diameter of the seedlings under native tree cover (Table 1). Seedlings growing under conspecific adults or L. leucocephala cover were supported by 56% of the leaf number when compared with the seedlings growing under native tree cover. Moreover, the maximum leaf length was reduced by 18% for the seedlings growing under conspecific adults or L. leucocephala when compared with seedlings growing under native tree cover. None of the seedlings growing under native tree cover died during the investigation. In contrast, about 40% of the seedlings under L. leucocephala cover died and about 60% of the seedlings under conspecific adult cover died.
The influence of V. parviflora canopy cover on C. micronesica seedlings was similar to that of L. leucocephala. The stem height of these seedlings exhibited two overlapping groups, with stem height under V. parviflora being intermediate between conspecific cover and native tree cover (Table 1). The seedlings under conspecific tree cover and V. parviflora cover exhibited stems that were 48% of the diameter of the seedlings under native tree cover. Seedlings growing under conspecific adults or V. parviflora cover were supported by 56% of the leaf number when compared with the seedlings growing under native tree cover. Moreover, the maximum leaf length was reduced by 17% for the seedlings growing under conspecific adults or V. parviflora when compared with seedlings growing under native tree cover. All of the seedlings growing under native tree cover survived until the end of the investigation. In contrast, seedling mortality in the other two plant cover types was similar to that in the L. leucocephala study.
These results revealed that a legacy of invasive L. leucaena or V. parviflora tree cover in northern Guam increased mortality and reduced growth of C. micronesica seedlings when compared with native tree cover. The influence of these invasive trees on C. micronesica seedling mortality was less severe than the influence of conspecific adult tree cover. The presence of A. yasumatsui throughout Guam since 2005 has led to regeneration failures and removed the potential for repeating this investigation.
3.5. Container Soil Conditioning Study
Each of the plant size traits of the soil conditioning study exhibited similar patterns for C. micronesica seedlings grown in soils conditioned by heterospecific native plant species and in soils conditioned by heterospecific invasive plant species (Table 2). Seedlings grown in the native plant treatments exhibited more robust size metrics and the seedlings grown in the invasive plant treatments exhibited less robust size metrics. In contrast, the seedlings grown in soils conditioned by conspecific plants exhibited heterogeneous relationship patterns with the seedlings grown in the other four soil treatments, in that some traits aligned with the seedlings grown in the native plant soils and other traits aligned with the seedlings grown in the invasive plant soils.
Stem height, stem diameter, and leaf number were greatest for C. micronesica seedlings grown in soils conditioned by heterospecific native plants than the other four soil conditioning treatments (Table 2). For the soils conditioned by invasive species, these metrics generally ranked in the order V. parviflora < C. papaya < L. leucocephala. Leaf length and leaf dry weight were greatest for C. micronesica seedlings grown in soils conditioned by conspecifics and heterospecific natives and least for the seedlings grown in soils conditioned by the three invasive species. Leaf number was greatest for seedlings grown in soils conditioned by heterospecific natives and least for soils conditioned by V. parviflora. All of the leaves of seedlings grown in soils conditioned by heterospecific natives were alive, but a mean of one leaf per seedling was dead for the seedlings grown in soils conditioned by conspecifics, and a mean of two leaves per seedlings were dead for the soils conditioned by the invasive species. The seedlings in the V. parviflora soils exhibited the smallest leaf traits. Stem dry weight, fine root dry weight, and large root dry weight were separated into three overlapping groups with similar patterns. Seedlings grown in soils conditioned by heterospecific natives exhibited a greater dry weight than the other four soil treatments, and the seedlings grown in soils conditioned by conspecifics and V. parviflora exhibited the least dry weights.
Coralloid root dry weight, total plant dry weight, and shoot:root ratio exhibited unique patterns among the five soil treatments (Table 2). For coralloid root dry weight, the seedlings grown in soils conditioned by conspecifics and heterospecific natives exhibited a dry weight that was 2.4-fold greater than that of seedlings grown in soils conditioned by the three invasive species. For total plant dry weight, the means revealed a pattern of V. parviflora < C. papaya = L. leucocephala < conspecifics < heterospecific natives for the conditioned soil treatments. The seedlings grown in soils conditioned by conspecifics exhibited the greatest shoot:root ratio. The seedlings grown in soils conditioned by the invasive tree species had shoot:root ratios that were only 66% of the values for conspecific soil treatment.
These results confirmed that legacy effects due to plant–soil feedback from three invasive tree species exerted profound influences on C. micronesica seedling growth. Leaf dry weight and coralloid root dry weight were more heavily influenced by the five soil conditioning treatments than the other plant traits.
4. Discussion
Observations of C. micronesica and C. nitida habitats revealed that small seedlings were consistently found beneath the canopy of most female trees. At the same time, juvenile plants were seldom found in the same space. These general observations indicated that ongoing and fairly rapid seedling mortality maintains an undulating pool of young seedlings with the deaths replaced by newly germinating seeds. Long-term monitoring of C. micronesica habitats in Yap revealed considerable amplitude in the standing seedling population, with declines or increases of 1000+ seedlings per ha repeatedly occurring within one-year periods [15]. I utilized a range of methods to determine the influence of antecedent and contemporary species identity on current Cycas survival and growth, and the results conformed to the tenets of the Janzen–Connell hypothesis, plant–soil feedback, and soil legacy effects. These findings may improve our understanding of distance- and density-dependent in situ seedling mortality, but also inform conservation decisions regarding translocation projects during species recovery programs.
First, the survival of natural high-density seedlings beneath maternal parents in one study and experimentally planted low-density seedlings beneath mature male conspecific trees in two studies confirmed that the seedlings in close proximity to a mature conspecific began to die in less than 1 year. The naturally dispersed seedlings reached 100% mortality in less than 8 year. At the same time, paired planting sites in sympatric native forest cover that were at least 10 m away from a conspecific individual exhibited 100% survival of seedlings in two experimentally planted studies. Second, experimentally planted C. micronesica seedlings in monostands of two invasive tree species in Guam revealed more rapid seedling mortality and reduced growth of survivors than for seedlings growing in adjacent native forest cover. Third, traditional soil conditioning treatments in container culture revealed that 9 months of conditioning with invasive tree species reduced C. micronesica seedling growth, conditioning with conspecifics reduced relative root growth but increased coralloid root growth, and conditioning with heterospecific native species stimulated whole plant growth.
The duration of antecedent soil conditioning, density and size asymmetry of competitors, and the degree of current plant stress may interact with the influence of soil legacy effects on Cycas seedling growth. The precise age of the adult conspecifics and invasives described in Section 3.1, Section 3.2, Section 3.3 and Section 3.4 were not known, but the large mature trees unquestionably conditioned the soils with roots and leaf litter for decades before the investigations were initiated. Moreover, the seedlings in these investigations were not provided any horticultural care, so they may have experienced consequential abiotic stress in the highly competitive forest setting. These in situ studies revealed rapid mortality among the high- and low-density seedlings in close proximity to a conspecific adult tree. In contrast, the container study employed 9 months of soil conditioning in Phase 1, and provided benign nursery conditions with no plant competition in Phase II. In this setting, every seedling survived for the duration of the three-year study. Moreover, conspecific predecessors and competitors were most damaging to C. micronesica seedling growth for the in situ investigations but heterospecific invasive predecessors were more damaging to C. micronesica seedling growth for the nursery investigation.
The results indicated that historical soil legacy effects and contemporary competition with other plants may interact to determine the extent and speed of mortality of newly germinating Cycas seedlings. Indeed, the influence of asymmetric competition with the adult parent tree [21] and density-dependent competition with same-age conspecifics [22] may be difficult to separate from soil conditioning effects due to plant–soil feedback. The results also illuminated the importance of augmenting ecologically relevant in situ investigations with controlled nursery investigations to tease apart all of the drivers of seedling mortality of threatened plant species.
4.1. Conservation Implications
Cycas micronesica is listed as Endangered under the IUCN Red List [23] and Threatened under the United States Endangered Species Act [24]. Reintroductions and translocations of endangered plant species are critical components of recovery plans for many imperiled plant species. Knowledge of the extent to which the identities of previous plant occupants of a recipient site can condition soils is needed to inform many conservation issues. Embracing the need to utilize knowledge about habitat requirements of conserved species may mean the differences between translocation success and failure [25]. The collective results herein point to the need to select biodiverse native forest cover for positioning Cycas transplants while refraining from the use of planting sites where mature conspecific or invasive tree species have conditioned the soils.
One tangible conservation action that exploits this new knowledge may proactively aid in situ regeneration. Indeed, passively allowing Cycas seeds to germinate beneath mature conspecific trees is detrimental to species recovery goals for endangered species, as the fate of these recruits is certain death. An inexpensive conservation action may include the harvest of mature seeds then manual dispersal to locations in the same area of occupancy that are at least 10 m away from a conspecific mature tree. Passive germination in these biodiverse sites may enable regeneration and recruitment in a manner that greatly benefits population growth without the need for an expensive conservation nursery. The selection of planting sites that vary in slope, aspect, exposure, and resident native tree species may improve outcomes.
In situ conservation of C. micronesica includes protecting local genes from non-local genes. This conservation need requires conservationists to ensure that C. micronesica germplasm is not translocated out of the area of occupancy from which it is derived [26] unless there are no sympatric C. micronesica plants in the recipient site [27]. The Guam C. micronesica population is highly structured [20] and the conservation community cannot justify translocation projects that risk moving genes into novel habitats where those genes do not belong [28]. I repeat the previously published recommendation that restoration sites designed for the receipt of salvaged C. micronesica plants should be constructed in forests adjacent to the source sites to ensure the genes in the restoration sites are local genes [27]. These methods would also maximize the likelihood that the transplants would be adapted to the soils within the recipient sites.
Cycad biologists have demonstrated increased interest in the function of coralloid roots in cycads [29,30,31]. These unusual root structures host endosymbionts that fix nitrogen and provide other services, but little is known about the role of the rhizosphere in pre-coralloid inception and further development of the root structure. One unexpected result herein was that Phase I conditioning of the bulk soil with conspecific plants or sympatric heterospecific native plants more than doubled the coralloid root dry weight in Phase II C. micronesica plants when compared to Phase I conditioning with invasive tree species. Continued research to more fully understand the legacy mechanisms by which the history of plant–soil feedback outcomes influence coralloid root development would provide relevant new knowledge for improving in situ and ex situ cycad conservation. I began with living soils rather than sterilized soils, so the native plant roots during Phase I may have selectively amplified the microbes that subsequently benefitted coralloid root development. Continued research may employ sterilized soils in similar studies to develop a greater understanding of how the abiotic and biotic attributes of soils influence cycad coralloid root organogenesis and ontogeny.
The use of an off-site garden setting for some of this research highlighted the utility of ex situ germplasm for expanding knowledge to inform in situ conservation needs. The in situ research reported herein was conducted prior to Guam’s invasion of A. yasumatsui, and a repeat of similar studies is no longer possible due to the ubiquitous invasive A. yasumatsui population that is lethal to in situ seedlings. Using the off-site production of seeds to provide experimental units for controlled studies is crucial for filling in the knowledge gaps for threatened species such as C. micronesica. This form of research may also be open to more relaxed methods than in situ research on threatened species, as in situ research of threatened species is under the control of federal biologists empowered with approving research plans and issuing permits. For example, the most accurate measure of plant growth is the increase in dry weight of the tissue [32]. There is a bias against use of this informative approach in the research of threatened taxa because of the destructive nature, a bias that may not be justified in case studies where in situ regeneration and recruitment are nil, such as C. micronesica on Guam. However, when seeds derived from off-site cultivated plants are of no conservation value due to adulterated or unknown provenance information, the approach should be considered more often to provide the most accurate new knowledge. In this ex situ study, the pollen parent originated from a Guam habitat that was distant from the ovulate parent’s original habitat, therefore the seedlings were of no conservation value because natural gene flow between distant habitats does not occur [20]. The non-destructive stem and leaf dimension traits of the seedlings growing under the five soil conditioning treatments did not conform to the unambiguous dry weight traits, and if the dry weights had not been reported herein, the new knowledge about the extreme stimulation of C. micronesica root growth resulting from the antecedent conditioning by native plants would have remained unreported.
Kin recognition during co-mingled Cycas seedling growth has been studied for two species, where a reduction in root growth in response to competition with close kin can be interpreted as cooperation [33]. A reduction in root growth in order to cooperate with half-siblings may be one of the reasons that low-distance, high-density seedlings exhibit reduced longevity in accordance with Janzen–Connell model predictions. The underlying mechanisms that drive this cooperative growth response are not fully understood, and may include rhizosphere plant–soil feedback loops or may include direct root contact communication. The results herein indicated that the Phase I soil legacy effects of conspecifics were enough to reduce root growth during Phase II of the conditioning studies, and revealed that soil conditioning may be partly responsible for the kin recognition growth responses of Cycas. As discussed above, the reduction in root growth in soils that were conditioned with conspecifics would not have been demonstrated in this study if organ dry weights had not been measured. Indeed, contemporary competition with the parent tree and half-siblings is not the only driver of Janzen–Connell seedling attrition, and the years of ephemeral conspecifics that conditioned the low-distance soils prior to germination of a new seedling also appear to contribute.
4.2. Cycas micronesica and Biogeochemical Cycling
Long-term growth of a sessile Cycas micronesica tree influences the soil chemistry to create small niche spaces [34]. When compared to nearby soils that were not conditioned by a C. micronesica plant, the soils beneath a mature tree exhibited increased nitrogen and mineralization rates, which changed consequential ecological stoichiometry characteristics. Among many sympatric forest species, C. micronesica exhibited leaf litter with recalcitrant chemical traits, predicting relatively slow decomposition and a decline in the rates of ecosystem nutrient cycling [35]. Indeed, the release of nitrogen and carbon from C. micronesica leaf litter was slower than from litter of other sympatric species [36].
These general descriptions of chemistry of the soils that had been conditioned by C. micronesica trees for many years illuminate two issues that deserve further study. First, we know that a mature tree directly changes the soils with which it interacts, and seedlings that germinate beneath the canopy of the maternal parent are destined to die before recruiting to the juvenile stage. However, how do these two processes interact or influence each other and are there any feedback loops in this system? For example, if a seedling lives for several years, develops coralloid roots, then actively recruits nitrogen-fixing endosymbionts, what role does the ultimate death of this seedling play in creating the increased pool of nitrogen beneath a female tree? This could be preliminarily studied by comparing the soil chemistry beneath old female trees under which hundreds or thousands of seedlings have lived and died versus beneath male trees where no or few seedlings have influenced the soils.
Second, the 2003 invasion of Guam by A. yasumatsui has imposed profound changes in how C. micronesica influences terrestrial system biogeochemistry. This gymnosperm tree was the most abundant tree in Guam prior to the invasion [37], and therefore influenced ecosystem properties as a foundation species. The substantial amounts of nitrogen that the population of this species contributed to the terrestrial ecosystems and the recalcitrant nature of sequestering minerals in the litter layer have been decimated since the invasion due to 96% C. micronesica plant mortality to date [15]. In addition to this direct influence on the number of individual trees contributing to ecosystem services, the armored scale form of herbivory also changed leaf litter chemistry such that scale-infested litter exhibited a greater mass loss and faster release of nitrogen, phosphorus, calcium, and magnesium than healthy litter that experienced no scale damage [38]. Moreover, several years of scale-infested leaf litter contributions and scale-induced tree mortality and decomposition created soil legacies beneath damaged C. micronesica trees that decreased sympatric plant growth [39]. The buildup of phytotoxic compounds by the decomposition of scale-damaged C. micronesica organs was partly mitigated by activated charcoal incubations. This unusual cascading plant–soil feedback response to the A. yasumatsui invasion deserves further study.
4.3. Invasive Plant Species
This study adds to the literature linking invasive plant species with conservation principles for imperiled native plants. Leucaena leucocephala is known to employ allelopathic properties during invasions that displace native plants [40,41]. In Guam, L. leucocephala and V. parviflora have been shown to change soil chemistry in a manner that differs from native tree cover [42,43]. The results herein verify that mature monostands of V. parviflora and L. leucocephala reduce C. micronesica seedling growth and survival. This is also the first time that experimental methods have shown that conditioning soils with plants of these two species and C. papaya reduced subsequent C. micronesica seedling growth. These results support past recommendations [43,44] that urge conservationists to develop protocols that ensure in situ protected areas and restoration sites used for translocations of C. micronesica remain free of these invasive tree species.
Soil conditioning by invasive tree species in this study reduced C. micronesica coralloid root growth by 50% when compared with soil conditioning by conspecific and heterospecific native tree species. Investments into the root module that hosts endosymbionts are costly, and this C. micronesica root growth response may have been a form of sanctioning non-cooperative symbionts within the invaded soils [45]. Complex signaling and communication occurs among neighboring plants, and a similar stimulation of Vicia faba L. root nodule growth by some but not all heterospecific neighbors has been attributed to root exudates [46]. The findings indicate that invasive tree species may reduce the capacity of co-occurring C. micronesica plants to contribute nitrogen to the terrestrial ecosystems.
Programs to restore unhealthy degraded forests into healthy native tree habitats have become a global agenda. Planting and active management of native tree species to displace invasive trees may be an effective approach for restoring degraded habitats [47]. However, widespread, abundant native tree taxa should be included in the palette of species to be used for this purpose, and the use of threatened taxa in degraded habitats has been discouraged [43]. The results herein support the suggestion to refrain from using threatened taxa such as C. micronesica to restore degraded habitats, as the action is contrary to species recovery goals.
4.4. Long-Term Approaches Needed
The fostering of biodiversity recovery in the face of global change requires planners and practitioners to understand that a long-term commitment to planning and adaptive management research is required [48]. When C. micronesica is planted in habitats that do not provide adequate species requirements, the consequences of maladaptation may require years to begin exposing the increases in mortality [49]. All evidence to date indicates that germination of a C. micronesica seedling directly beneath a conspecific adult individual is a guarantee of impending death, yet the results herein indicate the journey may require 8 years to reach the end. Conservation projects and species recovery plans may provide misleading and inaccurate information if the decision-makers employ the traditional one- to three-year timeline to assess outcomes of federally funded projects. This has been the unfortunate approach used by conservationists in Guam. Clearly, long-term monitoring on the scale of decades is required to assess the efficacy of conservation and restoration actions for C. micronesica.
5. Conclusions
Conservationists often plan projects designed to improve in situ recruitment or harvest plants for translocation projects. Knowledge concerning the habitat requirements for plant growth and survival is required to ensure the success of these projects. The recent literature concerning C. micronesica conservation on the island of Guam and the collective results herein underscore the need to select biodiverse native forest cover for positioning Cycas plants while refraining from the use of planting sites where conspecific or invasive tree species have conditioned the soils. These and similar recommendations have not been adopted by the conservation decision-makers in recent federally funded projects on the island of Guam, where degraded habitats have been selected as translocation sites for C. micronesica salvage projects. This case study has emerged as an example where meaningful engagement with the scientific community and the concerned public has not been integrated into the conservation planning processes, and the result is that this unique gymnosperm tree remains on the journey toward an extinction vortex.
Funding
This research was funded by the USDA CSREES grant number 2003-05495; and the United States Forest Service grant numbers 10-DG-11059702-095 and 13-DG-11052021-210.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available upon request.
Acknowledgments
Guam field support was provided by Nirmala Dongol, Gil Cruz, and Frankie Matanane. Matnog and Samar field support was provided by Jane Ferreras. Angeles City logistical and facilities support was provided by Gerard Sigua.
Conflicts of Interest
The author declares no conflict of interest.
References
- Bever, J.D. Soil community feedback and the coexistence of competitors: Conceptual frameworks and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Kulmatiski, A.; Beard, K.H.; Stevens, J.R.; Cobbold, S.M. Plant–soil feedbacks: A meta-analytical review. Ecol. Lett. 2008, 11, 980–992. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant–soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Bradford, M.A.; Pernilla Brinkman, E.; van de Voorde, T.F.; Veen, G.F. Where, when and how plant–soil feedback matters in a changing world. Funct. Ecol. 2016, 30, 1109–1121. [Google Scholar] [CrossRef]
- Fischer, A.P. Forest landscapes as social-ecological systems and implications for management. Landsc. Urban Plan. 2018, 177, 138–147. [Google Scholar] [CrossRef]
- Raum, S. Land-use legacies of twentieth-century forestry in the UK: A perspective. Landsc. Ecol. 2020, 35, 2713–2722. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Aci, M.M.; Sidari, R.; Araniti, F.; Lupini, A. Emerging trends in allelopathy: A genetic perspective for sustainable agriculture. Agronomy 2022, 12, 2043. [Google Scholar] [CrossRef]
- Jordan, N.R.; Larson, D.L.; Huerd, S.C. Soil modification by invasive plants: Effects on native and invasive species of mixed-grass prairies. Biol. Invasions 2008, 10, 177–190. [Google Scholar] [CrossRef]
- Pernilla Brinkman, E.; Van der Putten, W.H.; Bakker, E.-J.; Verhoeven, K.J.F. Plant–soil feedback: Experimental approaches, statistical analyses and ecological interpretations. J. Ecol. 2010, 98, 1063–1073. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In Dynamics of Populations; Den Boer, P.J., Gradwell, G.R., Eds.; Centre for Agricultural Publications and Documentation: Wageningen, The Netherlands, 1971; pp. 298–310. [Google Scholar]
- Janzen, D.H. The uncertain future of the tropics. Nat. Hist. 1972, 81, 80–89. [Google Scholar]
- Janzen, D.H. Tropical dry forests. In Biodiversity; Wilson, E.O., Peter, F.M., Eds.; National Academy Press: Washington, DC, USA, 1988; pp. 130–137. [Google Scholar]
- Marler, T.E.; Krishnapillai, M.V. Longitude, forest fragmentation, and plant size influence Cycas micronesica mortality following island insect invasions. Diversity 2020, 12, 194. [Google Scholar] [CrossRef]
- Deloso, B.E.; Terry, L.I.; Yudin, L.S.; Marler, T.E. Biotic threats to Cycas micronesica continue to expand to complicate conservation decisions. Insects 2020, 11, 888. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Lindström, A.J.; Watson, G.W. Aulacaspis yasumatsui delivers a blow to international cycad horticulture. Horticulturae 2021, 7, 147. [Google Scholar] [CrossRef]
- Marler, T.E.; Griffith, M.P.; Krishnapillai, M.V. Height increment of Cycas micronesica informs conservation decisions. Plant Signal. Behav. 2020, 15, e1830237. [Google Scholar] [CrossRef] [PubMed]
- Hirsh, H.; Marler, T. Damage and recovery of Cycas micronesica after Typhoon Paka. Biotropica 2002, 34, 598–602. [Google Scholar] [CrossRef]
- Cibrian-Jaramillo, A.; Daly, A.C.; Brenner, E.; DeSalle, R.; Marler, T.E. When North and South don’t mix: Genetic connectivity of a recently endangered oceanic cycad, Cycas micronesica, in Guam using EST-microsatellites. Mol. Ecol. 2010, 19, 2364–2379. [Google Scholar] [CrossRef]
- Ehlers, B.K.; Bilde, T. Inclusive fitness, asymmetric competition and kin selection in plants. Oikos 2019, 128, 765–774. [Google Scholar] [CrossRef]
- Postma, J.A.; Hecht, V.L.; Hikosaka, K.; Nord, E.A.; Pons, T.L.; Poorter, H. Dividing the pie: A quantitative review on plant density responses. Plant Cell Environ. 2021, 44, 1072–1094. [Google Scholar] [CrossRef]
- Bösenberg, J.D. Cycas micronesica. The IUCN Red List of Threatened Species 2022: E.T61316A68906033. Available online: https://apiv3.iucnredlist.org/api/v3/taxonredirect/61316 (accessed on 20 November 2023).
- United States Fish & Wildlife Service. Endangered and threatened wildlife and plants; endangered status for 16 species and threatened status for 7 species in Micronesia. Fed. Regist. 2015, 80, 59424–59497. [Google Scholar]
- Maschinski, J.; Albrecht, M.A. Center for Plant Conservation’s Best Practice Guidelines for the reintroduction of rare plants. Plant Divers. 2017, 39, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E. Conservation of local genotypes when planting native plants on Guam. Micrones. Suppl. 2002, 6, 55–60. [Google Scholar]
- Marler, T.E.; Lindström, A.J. First, do no harm. Communic. Integr. Biol. 2017, 10, e1393593. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Cruz, G.N. Adventitious rooting of mature Cycas micronesica K.D. Hill tree stems reveals moderate success for salvage of an endangered cycad. J. Threat. Taxa 2017, 9, 10565–10570. [Google Scholar] [CrossRef]
- Gutiérrez-García, K.; Bustos-Díaz, E.D.; Corona-Gómez, J.A.; Ramos-Aboites, H.E.; Sélem-Mojica, N.; Cruz-Morales, P.; Pérez-Farrera, M.A.; Barona-Gómez, F.; Cibrián-Jaramillo, A. Cycad coralloid roots contain bacterial communities including cyanobacteria and Caulobacter spp. that encode niche-specific biosynthetic gene clusters. Genome Biol. Evol. 2019, 11, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, J.; Xu, H.; Liu, J.; Zhao, Z.; Gong, X. Core microbiome and microbial community structure in coralloid roots of Cycas in ex situ collection of Kunming Botanical Garden in China. Microorganisms 2023, 11, 2144. [Google Scholar] [CrossRef] [PubMed]
- Ndlovu, S.; Suinyuy, T.N.; Pérez-Fernández, M.A.; Magadlela, A. Encephalartos natalensis, their nutrient-cycling microbes and enzymes: A story of successful trade-offs. Plants 2023, 12, 1034. [Google Scholar] [CrossRef] [PubMed]
- Leopold, A.C.; Kriedemann, P.E. Plant Growth and Development, 2nd ed.; McGraw-Hill: New York, NY, USA, 1975; p. 545. [Google Scholar]
- Marler, T.E.; Dongol, N.; Cruz, G.N. Plastic responses mediated by identity recognition in below-ground competition in Cycas micronesica K.D. Hill. Trop. Conserv. Sci. 2016, 9, 648–657. [Google Scholar] [CrossRef]
- Marler, T.E.; Calonje, M. Two cycad species affect the carbon, nitrogen, and phosphorus content of soils. Horticulturae 2020, 6, 24. [Google Scholar] [CrossRef]
- Marler, T.E. Leaf elemental concentrations, stoichiometry, and resorption in Guam’s coastal karst forests. Diversity 2021, 13, 545. [Google Scholar] [CrossRef]
- Paulino, C.A.; Marler, T.E. Nitrogen and carbon mineralization from green and senesced leaf litter differ between cycad and angiosperm trees. Biology 2022, 11, 1758. [Google Scholar] [CrossRef] [PubMed]
- Donnegan, J.A.; Butler, S.L.; Grabowiecki, W.; Hiserote, B.A.; Limtiaco, D. Guam’s Forest Resources, 2002; Resource Bulletin PNW-RB-243; Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2004.
- Marler, T.E. Aulacaspis yasumatsui infestations accelerate Cycas leaf litter decomposition and nutrient release. Pedosphere 2023, in press. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N. Do phytotoxic compounds in soils after scale-infested Cycas micronesica litter deposits explain reduced plant growth? HortScience 2013, 48, 1571–1573. [Google Scholar] [CrossRef]
- Sahoo, U.; Upadhyaya, K.; Meitei, C. Allelopathic effects of Leucaena leucocephala and Tectona grandis on germination and growth of maize. Allelopathy J. 2007, 20, 135–143. [Google Scholar]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelochemicals of Leucaena leucocephala as an invasive plant species. Plants 2022, 11, 1672. [Google Scholar] [CrossRef]
- Marler, T.E.; Dongol, N.; Cruz, G.N. Leucaena leucocephala and adjacent native limestone forest habitats contrast in soil properties on Tinian Island. Commun. Integr. Biol. 2016, 9, e1212792. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E. Three invasive tree species change soil chemistry in Guam forests. Forests 2020, 11, 279. [Google Scholar] [CrossRef]
- Lindström, A.; Terry, I.; Deloso, B.; Tang, W.; Donaldson, J.; Marler, T. Typhoon Mawar enables an assessment of Cycas micronesica conservation plans. J. Geogr. Nat. Disasters 2023, 13, 280. [Google Scholar]
- Mohd-Radzman, N.A.; Drapek, C. Compartmentalisation: A strategy for optimizing symbiosis and tradeoff management. Plant Cell Environ. 2023, 46, 2998–3011. [Google Scholar] [CrossRef]
- Callaway, R.M.; Li, L. Decisions, decisions, decisions: Plant roots detect and respond to complex environmental cues. New Phytol. 2020, 226, 11–12. [Google Scholar] [CrossRef]
- Ceccon, E.; Sánchezm, I.; Powers, J.S. Biological potential of four indigenous tree species from seasonally dry tropical forest for soil restoration. Agrofor. Syst. 2015, 89, 455–467. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Marler, T.E. Reciprocal garden study reveals acute spatial-edaphic adaptation for Cycas micronesica. Diversity 2021, 13, 237. [Google Scholar] [CrossRef]
Figure 1.
Location of five studies designed to understand competitor plant identity influences on Cycas micronesica and Cycas nitida seedling survival.
Figure 1.
Location of five studies designed to understand competitor plant identity influences on Cycas micronesica and Cycas nitida seedling survival.

Figure 2.
Numerous young Cycas micronesica seedlings characterized the space directly beneath female trees prior to the 2003 invasion of Guam by the scale Aulacaspis yasumatsui.
Figure 2.
Numerous young Cycas micronesica seedlings characterized the space directly beneath female trees prior to the 2003 invasion of Guam by the scale Aulacaspis yasumatsui.

Figure 3.
Invasive tree species that displace natives to develop monostands in Guam include (a) Vitex parviflora and (b) Leucaena leucocephala.
Figure 3.
Invasive tree species that displace natives to develop monostands in Guam include (a) Vitex parviflora and (b) Leucaena leucocephala.

Figure 4.
Number of seedlings beneath 50 female Cycas micronesica trees in northern Guam for an 18-month period. Mean ± SD.
Figure 4.
Number of seedlings beneath 50 female Cycas micronesica trees in northern Guam for an 18-month period. Mean ± SD.

Figure 5.
Number of original seedlings and total number of seedlings for a 4- to 8-year period. (a) Guam Cycas micronesica, n = 15; (b) Yap Cycas micronesica, n = 10; (c) Sorsogon Cycas nitida, n = 10; (d) Samar Cycas nitida, n = 12. Mean ± SD.
Figure 5.
Number of original seedlings and total number of seedlings for a 4- to 8-year period. (a) Guam Cycas micronesica, n = 15; (b) Yap Cycas micronesica, n = 10; (c) Sorsogon Cycas nitida, n = 10; (d) Samar Cycas nitida, n = 12. Mean ± SD.

Figure 6.
The influence of growing Cycas micronesica seedlings beneath a mature conspecific tree (Near) versus 10 to 15 m away from any conspecific individual (Away) on plant growth and mortality. (a) Number of living seedlings; (b) ending stem height (shaded bars) and diameter (open bars); (c) ending total leaf number; (d) maximum leaf length. Mean ± SD, n = 8.
Figure 6.
The influence of growing Cycas micronesica seedlings beneath a mature conspecific tree (Near) versus 10 to 15 m away from any conspecific individual (Away) on plant growth and mortality. (a) Number of living seedlings; (b) ending stem height (shaded bars) and diameter (open bars); (c) ending total leaf number; (d) maximum leaf length. Mean ± SD, n = 8.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Survival and plant size metrics for Cycas micronesica seedlings growing from March 2003 until June 2005 beneath conspecific adults, heterospecific native trees, Vitex parviflora, or Leucaena leucocephala trees. Mean ± SE, n = 6.
Table 1.
Survival and plant size metrics for Cycas micronesica seedlings growing from March 2003 until June 2005 beneath conspecific adults, heterospecific native trees, Vitex parviflora, or Leucaena leucocephala trees. Mean ± SE, n = 6.
| Cover Type | Stem Ht (cm) | Stem Dia (cm) | Leaf Number | Leaf Length (cm) | Proportion Alive Plants |
|---|---|---|---|---|---|
| Leucaena Study | |||||
| Conspecific | 10.1 ± 1.5 b y | 3.7 ± 0.7 b | 5 ± 1 b | 82.3 ± 7.2 b | 0.38 ± 0.06 c |
| Native z | 13.8 ± 1.0 a | 6.7 ± 0.9 a | 9 ± 1 a | 103.8 ± 7.7 a | 1.00 ± 0.00 a |
| Leucaena | 10.8 ± 1.2 b | 4.7 ± 0.8 b | 5 ± 1 b | 88.7 ± 8.6 b | 0.58 ± 0.05 b |
| Test statistic x | 15.192 | 32.167 | 31.623 | 10.630 | 14.488 |
| Significance | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Vitex Study | |||||
| Conspecific | 9.2 ± 1.0 b y | 3.4 ± 0.5 b | 5 ± 1 b | 86.3 ± 4.5 b | 0.42 ± 0.05 c |
| Native w | 11.8 ± 1.3 a | 6.9 ± 0.9 a | 9 ± 1 a | 107.8 ± 7.3 a | 1.00 ± 0.00 a |
| Vitex | 10.8 ± 0.8 ab | 3.5 ± 0.6 b | 5 ± 1 b | 92.0 ± 6.2 b | 0.63 ± 0.06 b |
| Test statistic x | 9.899 | 44.071 | 16.775 | 19.809 | 14.488 |
| Significance | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
z Heterospecific native species included Aglaia mariannensis, Morinda citrifolia, and Ochrosia oppositifolia. y Values with the same letter within columns are not significantly different. x F test statistic for plant size metrics; H test statistic for survival. w Heterospecific native species included Aglaia mariannensis, Meiogyne cylindrocarpa, and Ochrosia oppositifolia.
Table 2.
Container-grown plant size traits for Cycas micronesica seedlings grown from February 2020 until February 2023 in soils that were conditioned for nine months by conspecific seedlings, heterospecific native plants, Carica papaya, Leucaena leucocephala, or Vitex parviflora. Mean ± SE, n = 6.
Table 2.
Container-grown plant size traits for Cycas micronesica seedlings grown from February 2020 until February 2023 in soils that were conditioned for nine months by conspecific seedlings, heterospecific native plants, Carica papaya, Leucaena leucocephala, or Vitex parviflora. Mean ± SE, n = 6.
| Conditioning Taxa | Stem Ht (cm) | Stem Dia (cm) | Leaf Number | Leaf Length (cm) | Leaf dw (g) | Stem dw (g) | Fine Root dw (g) | Large Root dw (g) | Coralloid Root dw (g) | Total Plant dw (g) | Shoot: Root Ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Conspecific | 10.0 ± 0.3 b z | 4.4 ± 0.2 c | 6 ± 1 b | 91.7 ± 1.6 a | 53.7 ± 2.9 a | 46.3 ± 1.9 bc | 16.5 ± 0.8 c | 26.8 ± 1.1 b | 3.0 ± 0.2 a | 166.8 ± 4.3 b | 2.61 ± 0.11 a |
| Native y | 13.1 ± 0.4 a | 6.0 ± 0.2 a | 8 ± 1 a | 90.3 ± 1.8 a | 51.7 ± 3.1 a | 71.3 ± 2.5 a | 27.6 ± 1.9 a | 41.9 ± 3.9 a | 3.1 ± 0.3 a | 214.2 ± 6.8 a | 1.95 ± 0.08 b |
| Carica | 9.8 ± 0.3 b | 4.7 ± 0.1 b | 5 ± 1 bc | 74.7 ± 2.3 b | 32.7 ± 1.8 b | 49.9 ± 1.4 b | 20.0 ± 1.6 bc | 34.9 ± 2.9 ab | 1.3 ± 0.1 b | 151.5 ± 4.8 c | 1.72 ± 0.09 c |
| Leucaeana | 9.9 ± 0.3 b | 5.0 ± 0.1 b | 6 ± 1 b | 75.2 ± 1.9 b | 32.0 ± 1.2 b | 51.9 ± 1.4 b | 22.1 ± 1.8 ab | 33.7 ± 3.2 ab | 1.2 ± 0.1 b | 153.3 ± 6.1 c | 1.74 ± 0.10 c |
| Vitex | 9.3 ± 0.2 b | 4.6 ± 0.1 bc | 4 ± 1 c | 61.7 ± 2.3 c | 25.1 ± 1.1 b | 43.3 ± 1.8 c | 18.6 ± 0.5 c | 26.6 ± 1.3 b | 1.3 ± 0.1 b | 124.4 ± 2.8 d | 1.68 ± 0.08 c |
| F 4,25 | 26.041 | 44.485 | 14.695 | 24.824 | 37.899 | 33.142 | 9.906 | 8.414 | 131.285 | 45.563 | 12.521 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.005 | <0.001 | <0.001 | <0.001 |
z Values with the same letter within columns are not significantly different. y Heterospecific native species included Aglaia mariannensis, Morinda citrifolia, and Tabernaemontana rotensis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marler, T.E. Soil Conditioning and Neighbor Identity Influence on Cycas Seedling Performance. Ecologies 2023, 4, 731-748. https://doi.org/10.3390/ecologies4040048
AMA Style
Marler TE. Soil Conditioning and Neighbor Identity Influence on Cycas Seedling Performance. Ecologies. 2023; 4(4):731-748. https://doi.org/10.3390/ecologies4040048
Chicago/Turabian StyleMarler, Thomas E. 2023. "Soil Conditioning and Neighbor Identity Influence on Cycas Seedling Performance" Ecologies 4, no. 4: 731-748. https://doi.org/10.3390/ecologies4040048